微小花蝽对梨瘿蚊的室内捕食作用
2020-03-27胡长效强承魁王胜永
胡长效 强承魁 王胜永

摘要:为明确各虫态微小花蝽(Orius minutes L.)对梨瘿蚊(Dasumeira pyri Bouch)幼虫的控害潜能,室内研究了微小花蝽1~5龄若虫及雌成虫捕食梨瘿蚊3~4龄幼虫的功能反应、寻找效应、种内干扰及竞争作用。结果表明,微小花蝽1~5龄若虫及雌成虫对梨瘿蚊3~4龄幼虫的捕食功能反应均属于HollingⅡ型圆盘方程和HollingⅢ型功能反应新模型。寻找效应(S)随猎物密度(N)的增加而降低,呈负线性相关关系,微小花蝽1~5龄若虫及雌成虫的最佳寻找密度(b)分别为8.916 4、11.944 5、20.935 7、23.316 0、29.846 1、25.898 4头。随自身密度(P)的增加,微小花蝽1~5龄若虫及雌成虫对梨瘿蚊幼虫捕食作用率逐渐减小,干扰和竞争均增大,干扰系数(m)分别为0.584 1、0.642 5、0.590 8、0.549 7、0.577 2、0.612 7,分摊竞争强度(I)与自身密度的常用对数(lgP)呈正线性相关关系。说明微小花蝽对梨瘿蚊幼虫具有良好的控害潜能。
关键词:微小花蝽;梨瘿蚊;功能反应;寻找效应;干扰系数;分摊竞争强度
中图分类号:S436.612.2+9文献标识码:A文章编号:1000-4440(2020)01-0057-06
Abstract:To clarify the potential pest control capacity of Orius minutes on the larva of Dasumeira pyri, the functional response, searching efficiency, intraspecific interference and competitive intensity of 1st-5th instar nymphs and female adults of O. minutes on 3rd-4th instar nymphs of D. pyri were observed under laboratory conditions. The results showed that the predation efficacies of O. minutes on D. pyri all followed Holling-II disc equations and new model of Holling-Ⅲfunctional response. There was a negative relationship between searching efficiency(S)and prey density(N), and the optimum searching density(b)of 1st-5th instar nymphs and female adults of O. minutes was 8.916 4, 11.944 5, 20.935 7, 23.316 0, 29.846 1 and 25.898 4, respectively. Predation rate was negatively correlated with predator self-density(P), and mutual interference and competition were strengthened with the interference coefficients(m)of 0.584 1, 0.642 5, 0.590 8, 0.549 7, 0.577 2, 0.612 7. In addition, there was a positive relationship between intensity of scrambling competition(I) and common logarithm of prey density(lgP). These results indicate that the O. minutes is a potential natural enemy for D. pyri.
Key words:Orius minutus;Dasumeira pyri;functional response;searching efficiency;interference coefficient;intensity of scrambling competition
梨癭蚊(Dasumeira pyri Bouch), 属双翅目(Diptera)瘿蚊科(Cecidomyiidae),寄主仅为梨,主要以幼虫危害梨新梢芽叶,导致叶片纵卷,变硬变脆成肿瘤状,失绿呈紫红色,危害严重时新梢中下部叶片大量脱落[1-4]。中国1984年在安徽省首次发现梨瘿蚊,现已遍及江苏、浙江等15个省,成为中国梨树生产上的重要害虫[2-5]。梨瘿蚊幼虫喜躲藏在卷叶中取食,田间呈聚集分布[6-7],化学农药防治效果不理想。因此,保护和利用天敌控制梨瘿蚊已引起人们的关注。
微小花蝽(Orius minutes L.),属半翅目(Hemiptera)花蝽科(Anthocoridae),是古北区重要的捕食性天敌昆虫,国内分布广泛,以长江以北各地密度较高[8]。国内学者对微小花蝽捕食朱砂叶螨、桃蚜、西花蓟马、牛角花齿蓟马、烟粉虱、温室白粉虱、小菜蛾等的功能反应研究结果表明,微小花蝽对多种猎物均有较强的控制能力[9-14] 。笔者在田间多次观察到微小花蝽捕食梨瘿蚊幼虫情况,但迄今为止,在小花蝽、瓢虫、草蛉、蜘蛛及瘿蚊广腹细蜂等众多梨瘿蚊天敌中[1],仅见有异色瓢虫对梨瘿蚊捕食功能反应的研究报道[15]。鉴于此,本研究利用 HollingⅡ模型、Holling-Ⅲ型功能反应新模型、Hassell模型、邹运鼎等分摊竞争强度算式等对微小花蝽捕食梨瘿蚊幼虫的功能反应、种内干扰反应、最佳寻找密度、分摊竞争强度进行拟合或测定,以期为合理利用微小花蝽控制梨瘿蚊提供理论依据和数据支撑。
1材料与方法
1.1供试虫源
供试微小花蝽成虫采自徐州生物工程职业技术学院琴湖园艺实验场,用小号毛笔将成虫配对,置于培养皿(14.90 cm×2.85 cm)内用梨瘿蚊幼虫为猎物进行饲养,以四季豆豆荚为产卵基质,培养皿上覆盖150目纱布并用橡皮筋固定,置于HPJ-280型人工气候箱,温度为(25±1) ℃,相对湿度为70%±5%,光周期为16∶8(昼∶夜)。梨瘿蚊3~4龄幼虫采自徐州生物工程职业技术学院梨园试验田,梨树品种为圆黄,树龄24年,当年未施用任何农药,梨瘿蚊幼虫现采现用。
1.2试验方法
1.2.1微小花蝽对梨瘿蚊幼虫的捕食功能反应微小花蝽饥饿24 h后,置于培养皿(14.90 cm×2.85 cm)中,底部铺湿润滤纸保湿,放梨树嫩叶4片,每皿投放1头微小花蝽。微小花蝽1龄若虫捕食功能反应试验中,梨瘿蚊3~4龄幼虫密度梯度设为每皿5、10、20、30、40、50头;微小花蝽2龄若虫捕食功能反应试验中,梨瘿蚊3~4龄幼虫密度梯度设为每皿10、15、20、35、50、75头;微小花蝽3龄若虫、4龄若虫、5龄若虫、成虫捕食功能反应试验中,梨瘿蚊3~4龄幼虫密度梯度设为每皿20、40、60、90、120、150头,每个密度梯度重复5次,培养皿上盖150目纱布并用橡皮筋固定,试验在HPJ-280型人工气候箱中进行,饲养条件同方法1.1,24 h后统计剩余的猎物数量。
1.2.2微小花蝽自身密度对梨瘿蚊幼虫的寻找反应培养皿(14.90 cm×2.85 cm)底部铺湿润滤纸保湿,放梨树嫩叶4片,每皿放入200头梨瘿蚊3~4龄幼虫,微小花蝽1~5龄若虫、雌成虫的密度梯度均为每皿1、2、3、4、5头,每个处理重复5次,培养皿上盖150目纱布并用橡皮筋固定,试验在HPJ-280型人工气候箱中进行,饲养条件同方法1.1,24 h后统计剩余的猎物数量。
1.3数据分析
功能反应[16-17]:用 HollingⅡ圆盘方程Na=a′TN/(1+a′ThN)和HollingⅢ新模型方程Na=a·exp(-bN-1)进行拟合,式中,Na为天敌捕食量,N为猎物初始密度,a′为瞬时攻击率,T为搜寻总时间(本试验为1 d),Th为处理1头猎物消耗的时间,b为最佳寻找密度,a为捕食上限。两方程分别直线化为1Na=1a′1N+Th和lnNa=lna-b1N ,用excel2013和DPS9.5软件對数据进行处理。
寻找效应[16,18-19]:根据Holling的方法[16]进行估算,寻找效应 ( S ) 与害虫种群密度(N)有关的模型计算公式为:S=a′/(1+a′ThN)。
干扰效应[20]:采用Hassell-&Varley干扰模型E=NaNP或E=QP-m,式中,E为捕食效率,Na为捕食的猎物数,N为猎物密度,P为捕食者密度,Q为搜索常数,m为干扰系数。方程直线化为lnE=lnQ-mlnP,用excel2013和DPS9.5软件对数据进行处理。
分摊竞争强度[21]:估算公式为:I=(E1-Ep)/E1,式中, I为分摊竞争强度, E1为1头天敌的捕食作用率,Ep为密度为P的天敌捕食作用率。
2结果与分析
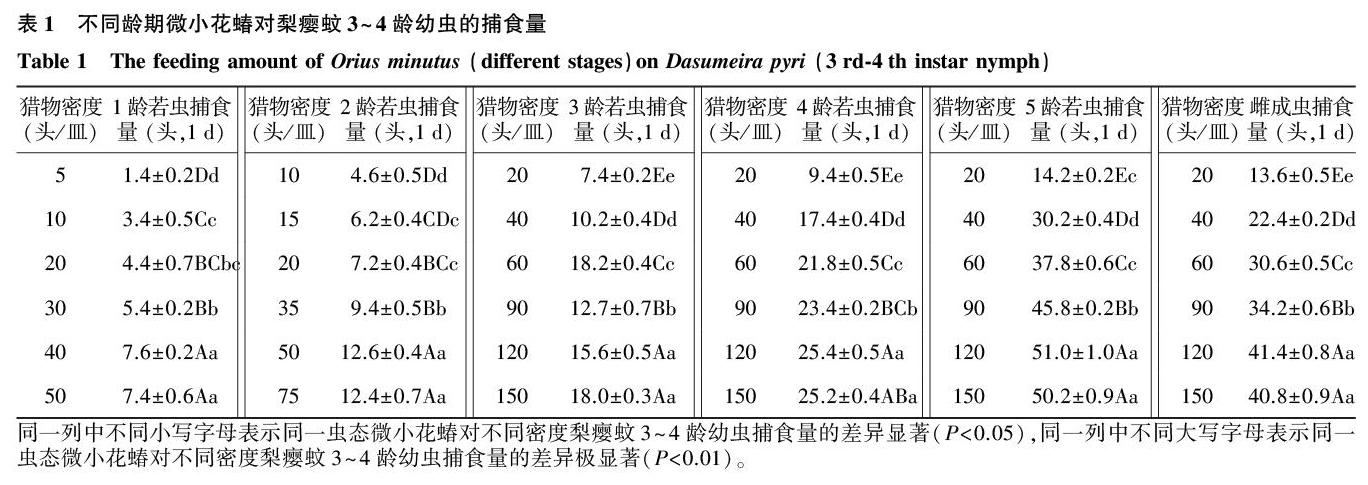
2.1微小花蝽各虫态对梨瘿蚊3~4龄幼虫的捕食量
微小花蝽各虫态在试验设置的梨瘿蚊幼虫密度下,捕食量随供试猎物密度的增加而增加,当增加到一定程度时增势减缓或略微下降,微小花蝽各虫态对梨瘿蚊幼虫的捕食功能反应符合HollingⅡ圆盘方程(表1)。不同虫态微小花蝽日平均捕食量大小顺序为:5龄若虫>雌成虫>4龄若虫>3龄若虫>2龄若虫>1龄若虫,但通过对供试猎物密度相同的微小花蝽3、4、5龄若虫、雌成虫差异显著性分析发现,在梨瘿蚊幼虫供试密度为每皿20头条件下,微小花蝽5龄若虫与雌成虫捕食量的差异不显著。
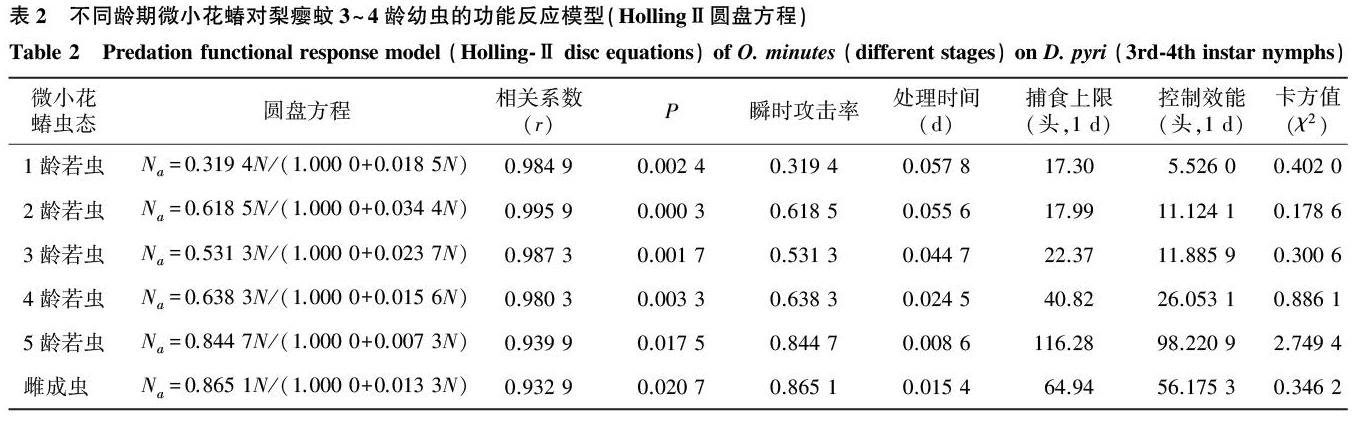
2.2微小花蝽各虫态对梨瘿蚊3~4龄幼虫的捕食功能反应
HollingⅡ圆盘方程较好地描述了微小花蝽各龄若虫及成虫对梨瘿蚊幼虫的捕食效应(r>r0.05,χ2<χ2(5,0.05)=11.071)。微小花蝽若虫对梨瘿蚊3~4龄幼虫的处理时间(Th)随虫龄的增加逐渐缩短,雌成虫处理时间(Th)介于4、5龄若虫之间。当 N→∞时,微小花蝽1~5龄若虫及雌成虫最大捕食量分别为17.30头、17.99头、22.37头、40.82头、116.28头、64.94头,瞬时攻击率(a′)表现为:雌成虫>5龄若虫>4龄若虫>2龄若虫>3龄若虫>1龄若虫(表2)。
2.3微小花蝽各虫态对梨瘿蚊3~4龄幼虫的寻找效应
2.3.1微小花蝽各虫态寻找效应与梨瘿蚊3~4龄幼虫密度的关系将捕食功能反应模型中参数值a′、Th及梨瘿蚊3~4龄幼虫供试密度值N,代入S=a′/(1+a′ThN)计算出不同梨瘿蚊3~4龄幼虫密度(N)下的寻找效应(S),S与N数据拟合模型分别为:S1龄若虫=0.297 3-0.002 8N(r=-0.988 2,p=0.001 5),S2龄若虫=-0.466 6-0.004 3N(r=-0.964 9,p=0.007 9),S3龄若虫=0.353 8-0.001 8N(r=-0.949 9,p=0.013 4),S4龄若虫=0.488 6-0.002 2N(r=-0.965 5,p=0.007 6),S5龄若虫=0.759 4-0.002 5N(r=-0.985 8,p=0.002 0),S雌成虫=-0.691 5-0.002 9N(r=-0.970 8,p=0.006 0)。由此可见,微小花蝽1~5龄若虫及成虫对梨瘿蚊3~4龄幼虫的寻找效应均随着猎物密度的增加而降低,呈负线性相关关系。
2.3.2微小花蝽各虫态对梨瘿蚊3~4龄幼虫的最佳寻找密度对本试验数据进行HollingⅢ型功能反应新模型拟合,算出最佳寻找密度(b)、捕食上限(a)及新模型方程Na=a·exp(-bN-1),经相关分析,所得r值均大于r0.05(P<0.05)。利用新模型方程计算所得的理论捕食量与实际捕食量进行χ2检验,所得χ2值均小于χ2(5,0.05) =11.071,表明所得新模型能够较好反映不同虫态微小花蝽对梨瘿蚊幼虫的捕食功能反应。微小花蝽1~5龄若虫及成虫最佳寻找密度分别为8.916 4头、11.944 5头、20.935 7头、23.316 0头、29.846 1头、25.898 4头,益害比1∶9、1∶12、1∶21、1∶23、1∶30、1∶26可作为微小花蝽对梨瘿蚊幼虫的生物防治指标的参考值(表3)。
2.4微小花蝽对梨瘿蚊幼虫捕食作用的种内干扰
在猎物密度为200头/皿时,微小花蝽平均捕食量(Na))和实际捕食作用率(E=Na/N·P)均随自身密度的增大逐渐减少(表4)。采用Hassell-Varley干扰反应模型E=QP-m拟合,得到干扰常数(m)、搜索常数(Q)及微小花蝽各虫态捕食梨瘿蚊幼虫的自我干扰方程。相关分析结果显示,所得r值均大于r0.05(P<0.05),利用自我干扰方程计算所得的理论捕食作用率与实际捕食作用率进行χ2检验,所得χ2值均小于χ2(4,0.05) =9.488,表明所得模型能够较好反映不同虫态微小花蝽对梨瘿蚊幼虫捕食作用的种内干扰情况。不同虫态间的干扰常数大小顺序为:2龄若虫>雌成虫> 3龄若虫>1龄若虫>5龄若虫>4龄若虫(表4)。
利用邹运鼎等[21]方法计算各虫态微小花蝽捕食梨瘿蚊幼虫所产生的分摊竞争强度(I)。结果(表5)显示,微小花蝽1~5龄若虫、雌成虫捕食梨瘿蚊幼虫所产生分摊竞争强度(I)与自身密度(P)数据拟合所得模型分别为:I1龄若虫=0.031 1+0.889 8lgP(r=0.990 3,p=0.009 7),I2龄若虫=-0.037 5+0.941 6lgP(r=0.988 4,p=0.011 6),I3龄若虫=0.038 0+0.875 9lgP(r=0.986 0,p=0.014 0),I4龄若虫=0.025 7+0.839 5lgP(r=0.992 0,p=0.008 0),I5龄若虫=0.007 2+0.872 7lgP(r=0.999 4,p=0.000 6),I雌成蟲=-0.002 8+0.907 3lgP(r=0.999 7,p=0.000 3),由此可以看出,随着微小花蝽自身密度的增加种内分摊竞争强度也随之增强。
3讨论
本试验中不同龄期微小花蝽对梨瘿蚊幼虫的捕食量与梨瘿蚊幼虫的密度呈负加速曲线,同时符合HollingⅡ型圆盘方程和HollingⅢ新模型方程。HollingⅡ模型中参数1/Th与HollingⅢ型功能反应新模型中参数a均代表日最大捕食量,但1/Th值明显大于a,这可能与两模型建立的基础是基于天敌食欲一直是无限的还是基于最佳猎物密度时才会发挥最积极的搜索运动有关[17]。显然,天敌的食欲不可能是无限的,因此利用HollingⅢ新模型方程拟合获得的参数值更具科学性和适用性,方程中参数b值(最佳寻找密度)对田间天敌的释放密度设计具有一定指导意义。
微小花蝽对梨瘿蚊幼虫的捕食量、瞬时攻击率、控制效果均随自身龄期的增高而增加,5龄若虫达到最高,雌成虫略低于5龄若虫。这可能与5龄若虫羽化前需要储存更多的物质和能量用于发育繁殖有关[22]。这与孙丽娟等[12]研究微小花蝽捕食小菜蛾(Plutella xylostella)的结果以及宋苞雪等[10]研究微小花蝽捕食烟粉虱(Bemisia tabaci)、朱砂叶螨 (Tetranychus cinnabarinus)、西花蓟马(Frankliniella occidentalis)、桃蚜 (Myzus persicae)的结果一致,也与曾广等[23-24]研究方小花蝽捕食烟蚜的结果以及付步礼等[25]研究东亚小花蝽捕食黄胸蓟马的结果一致。因此,田间释放微小花蝽应以5龄若虫为宜。
根据微小花蝽在无竞争状态下的最佳寻找密度,可拟定微小花蝽5龄若虫在田间释放比例为益害比1∶30。但由于本试验是在室内特定条件下进行的,空间、温湿度、光照、捕食者与猎物自身密度等与自然条件均存在一定的差异,特别是田间梨蚜、梨木虱、梨网蝽等多种猎物并存及梨瘿蚊幼虫又喜躲藏在卷叶中,因此,还应通过田间观察及笼罩试验,对此益害比值进行修正。
参考文献:
[1]胡长效,朱静,张芋,等. 梨瘿蚊的生物学、生态学及其防治[J].中国植保导刊,2005,25(7):11-13.
[2]万津瑜,周玲,张青文,等. 梨瘿蚊生物学及综合防治研究进展治[J].北方园艺,2012(14):194-196.
[3]江奇卿,张晓阳,金健,等. 江西梨瘿蚊发生与综合防治技术[J].生物灾害科学,2015, 38(1): 27-30.
[4]杨晓平,胡红菊,陈启亮,等. 武汉地区梨瘿蚊生物学特性及其种群动态变化研究[J].北方园艺,2016(13):111-115.
[5]蔡平. 梨瘿蚊研究初报[J].安徽农学院学报,1984(1):60-65.
[6]田瑞,胡红菊,王友平,等. 梨瘿蚊幼虫的空间分布型及序贯抽样技术[J].华中农业大学学报,2008,27(6):728-731.
[7]胡长效,朱静,张芋,等. 梨瘿蚊幼虫空间分布型及二阶抽样技术研究[J].安徽农业科学,2005,33(4):587-588.
[8]章士美,赵泳祥. 中国农林昆虫地理分布 [M].北京:中国农业出版社,1996:97.
[9]李向永,陈福寿,赵雪晴,等. 微小花蝽的发生及其对西花蓟马的捕食作用[J].环境昆虫学报, 2011,33(3): 346-350.
[10]宋苞雪,杨群芳,李庆,等. 微小花蝽对四种猎物的喜好性[J].应用昆虫学报 , 2018, 55(1): 112-116.
[11]汤方,李生臣,刘玉升,等. 微小花蝽对温室白粉虱的捕食作用[J].昆虫知识,2007,44(5):703-706.
[12]孙丽娟,衣维贤,郑长英. 微小花蝽对小菜蛾捕食控制的能力[J]. 应用生态学报,2017,28(10): 3403-3408.
[13]张世泽,吴林,许向利,等. 小花蝽对牛角花齿蓟马的捕食作用[J].应用生态学报,2006,17(7):1259-1263.
[14]李向永,陈福寿,赵雪晴,等. 微小花蝽的发生及其对西花蓟马的捕食作用[J]. 环境昆虫学报,2011,33 (3): 346-350.
[15]杜浩,高旭辉,刘坤,等. 异色瓢虫对梨瘿蚊幼虫的捕食功能反应及捕食偏好[J]. 中国生物防治学报,2017,33(6):811-816.
[16] HOLLING C S. Some characteristics of simple types of predation and parasitism[J]. The Canadian Entomologist, 1959, 91(7): 385-398.
[17]汪世泽,夏楚贵. Holling-Ⅲ型功能反应新模型[J]. 生态学杂志,1988,7(1):1-3.
[18]丁岩钦,兰仲雄,陈玉平. 天敌-害虫系统中寻找效应数学模型的研究[J]. 生态学报,1983,3(2):141-147.
[19]丁岩钦. 昆虫数学生态学[M].北京:科学出版社,1994:304.
[20]HASSELL M P, VARLEY G C. New inductive population model for insect parasites and its bearing on biological control [J]. Nature,1969,223:1133-1137.
[21]邹运鼎,耿继光,陈高潮,等. 异色瓢虫若虫对麦二叉蚜的捕食作用[J]. 应用生态学报,1996,7(2):197-200.
[22]唐艺婷,郭义,何國玮,等. 不同龄期的益蝽对粘虫的捕食功能反应[J]. 中国生物防治学报,2018,34(6):825-830.
[23]曾广,郅军锐,张昌容,等. 南方小花蝽对烟蚜的捕食作用[J]. 西南师范大学学报(自然科学版),2018,43(1):81-88.
[24]曾广,郅军锐,张昌容,等. 南方小花蝽控制害虫的研究现状[J]. 山地农业生物学报, 2017,36(1):71-75.
[25]付步礼,邱海燕,李强,等. 东亚小花蝽对黄胸蓟马的室内捕食作用研究[J]. 应用昆虫学报,2019, 56(1): 91-98.
(责任编辑:张震林)
