新型间日疟原虫红细胞结合蛋白在中国中部及周边国家边境地区的遗传多样性研究*
2020-03-25张杨明慧张学星魏海潮胡钰冰王庆辉曹雅明
张杨明慧 张学星 赵 艳 魏海潮 胡钰冰 庞 维 王庆辉 曹雅明
(中国医科大学基础医学院,免疫学教研室,辽宁沈阳 110122)
疟疾的高发病率和致死率造成全世界严重的健康负担,其中,间日疟原虫已成为全球分布最广的疟原虫种类(Battleetal., 2019)。开发血液期疫苗是抗疟最具实际效能的策略之一,由于裂殖子表面呈现高度多态性的抗原常介导疟原虫的免疫逃避(Patarroyoetal., 2020),进而针对多态性抗原的疫苗无法诱导广泛的交叉保护作用,仅显示出对同源虫株的功效(Ouattaraetal., 2015)。因此,研究间日疟原虫候选抗原的遗传多样性对规避抗原变异及后续疫苗的设计与评估至关重要。
研究发现,疟原虫的初始入侵由裂殖子表面蛋白与宿主细胞相应配体的结合介导(Cowmanetal., 2017)。目前,仅报道了两种与间日疟原虫裂殖子入侵网织红细胞相关的关键受体—配体相互作用,即间日疟原虫达菲结合蛋白1—达菲抗原趋化因子受体(PvDBP1-DARC)和间日疟原虫网织红细胞结合蛋白2b—转铁蛋白受体(PvRBP2b-CD71)(Patarroyoetal., 2020)。近年一项研究从柬埔寨间日疟患者的基因组(C127)中鉴定出一种新型红细胞结合蛋白(EBP),属红细胞结合样(EBL)蛋白超家族的成员,与间日疟原虫达菲结合蛋白(DBP)同源,预测其具有存在II区富含半胱氨酸保守的达菲结合样结构域(DBL)和C端富含半胱氨酸的结构域(Hesteretal., 2013)。进一步功能研究还表明,重组PvEBP不直接依赖DARC,优先与未成熟并高表达CD71的网织红细胞结合,并表现出与达菲阴性网织红细胞的最小结合能力,由此推测PvEBP可能是驱动间日疟原虫侵袭网织红细胞的替代途径,使得间日疟原虫非常规侵袭达菲阴性网织红细胞(Ntumngiaetal., 2016; Golassaetal., 2020)。在疟疾流行区,间日疟患者体内还可检测到高分泌水平的PvEBP-RII抗体,该抗体对临床间日疟显示出较强的独立保护作用(Heetal., 2019)。此外,通过对马达加斯加流行区间日疟分离株分析证实PvEBP-RII蛋白在介导疟原虫侵袭红细胞过程中的重要作用,并提出其可作为诱导机体产生获得性免疫的重要靶标(Roeschetal., 2018)。
本研究通过对PvEBP的遗传多样性、基因中性选择和分化程度及其单倍型网络图聚类研究,揭示出PvEBP在中国中部地区及周边国家边境地区流行特点,为寻找有效的、特异性的疫苗靶向位点提供新的分子基础。
1 材料与方法
1.1 试剂
TIANamp Blood Spots DNA Kit(DP334-03)购于中国天根公司,10×KOD-Plus-Neo 缓冲液、dNTPs、MgSO4、DL2000 DNA Marker(3427 A)和10×Loading Buffer(KA1801 A)购于日本TaKaRa公司,KOD-Plus-Neo DNA聚合酶(KOD-401)购于日本TOYOBO公司。
1.2 间日疟原虫样本采集
经中国医科大学伦理委员会和缅甸卫生体育部医学研究系伦理委员会审批,并获得所有参与者及其监护人的知情同意下,本研究共采集到53份间日疟原虫患者血液样本(滤纸血及血涂片),其中包括2009年中国(安徽,n=31)和2016年缅甸(Laiza,n=22)样本。同时从GenBank中获取越南间日疟原虫样本序列24例(MN853276—MN853300)和缅甸间日疟原虫样本序列10例(MN853178—MN853187)(Hanetal., 2020),对所有样本进行基因多态性和种群遗传学的分析。
1.3 PvEBP基因的PCR扩增及测序
根据TIANamp Blood Spots DNA Kit试剂盒说明书,从采集到的53份间日疟原虫感染患者的干滤纸血片中提取间日疟原虫基因组DNA。以提取的DNA为模板,PCR扩增PvEBP基因(核苷酸位点:1~2 453 bp),特异性引物F1: 5′-C T G T A C T T G T T T T G G T C A A T T T G T G T A GC-3′,R1: 5′-G T A T G A T C C A T G A T T C A T A G G C T G T G C A TG-3′,F2: 5′-T G G A C T T C T T A A C G A A A A T A G A T G C A A T A A CG-3′,R2: 5′-A C A T A T T T C C T G A C T T G T T T G C C G CC-3′。30 μL的PCR反应体系包括:10×KOD-Plus-Neo buffer,2 mmol/L dNTPs,25 mmol/L MgSO4,0.6 U KOD-Plus-Neo DNA聚合酶,0.2 μL上下游引物及1 μL DNA模板。扩增反应条件均为:94℃ 2 min;98℃ 10 s,58℃ 30 s,68℃ 45 s,共40个循环;68℃ 7 min。取用1.2%琼脂糖凝胶电泳检测PCR产物,送至北京诺赛基因公司测序,测序引物为:SGM-A3825: 5′-A C C T T A A C C T C G T C A G T G AA-3′,SGM-A3826: 5′-A T T G C G A T T C A G T G T T T A G TG-3′;SGM-I7182: 5′-T C G T C A G T C C C T T C T C A A A T C AA-3′,SGM-A3828: 5′-T G T C C T G G A A A A G T T A T GC-3′。
1.4 PvEBP基因序列比对与多态性分析
测序得到的序列及GenBank中下载的PvEBP基因序列通过MEGA7.0软件进行拼接,并以参考序列C127(GenBank: KC987954.1)作为标准进行DNA序列比对。通过DnaSP 6.12.03软件计算分离位点数目(number of segregating sites, S)、核苷酸多样性(nucleotide diversity,π)、单倍体型数目(number of haplotypes, H)和单倍型多样性(haplotype diversity, Hd)。
1.5 PvEBP基因的中性检验及基因重组分析
为确定PvEBP基因是否偏离中性选择及存在的自然选择压力,使用DnaSP 6.12.03软件计算Tajima′s D,Fu-Li′s F*&D*检验值。根据MEGA7.0中改良的Nei-Gojobori方法,基于密码子选择的Z检验计算非同义替换(dN)和同义替换(dS)的比率。使用DnaSP 6.12.03软件评估基因的最小重组事件(Rm)。
1.6 PvEBP基因的群体分化(Fst)检验
通过ARLEQUIN v3.5.2.2软件中Wright固定指数(Fst)计算等位基因频率的差异,衡量PvEBP基因在不同群体间的分化程度和遗传结构(Excoffieretal., 2010)。Fst的取值范围为0-1,当取值为0时,意味着不同地方群体遗传结构完全一致,群体间没有分化;取值为0.15时,认为群体间存在较大程度的遗传分化;取值为1时,表明等位基因在各地方群体中固定,完全分化。
1.7 PvEBP基因系统发育及单倍型网络分析
采用MEGA 7.0中邻接法对所有序列构建系统发育进化树,并通过1000次重复步长检验并得出节点可信值,从而推测对比PvEBP基因在不同地区之间的关联程度及亲缘关系。采用PopART 1.7中中位数连接法绘制单倍型网络图,以探讨出PvEBP基因在种群间的谱系关系。

表1 流行区分离株PvEBP基因序列多样性分析
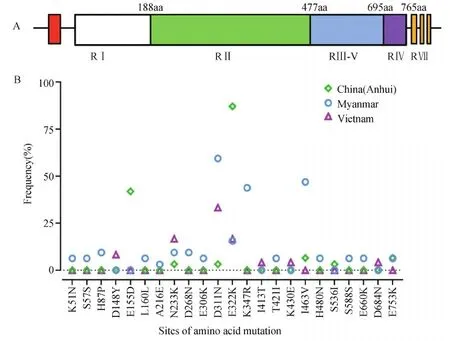
图1 PvEBP氨基酸多态位点频率及分布
2 结果
2.1 PvEBP基因多样性特征
对比3个地区间日疟原虫分离株PvEBP基因(1-2 325 bp)的多态性,中国安徽PvEBP基因核苷酸多样性(π=0.00051)和单倍型多样性(Hd=0.634)较低,缅甸PvEBP基因核苷酸多样性(π=0.00149)和越南PvEBP基因单倍型多样性(Hd=0.790)较高(表1)。从氨基酸位点上共得到20处非同义突变和3处同义突变,且主要发生在PvEBP-RII中(图1-A)。3个地区分离株中均存在N233K,D311N和E322K位点突变,其中后者E322K在中国安徽的突变率较高(87.1%)(图1-B)。
2.2 PvEBP-RII基因自然选择和重组分析
缅甸分离株PvEBP-RII的Tajima′s D>0,Fu-Li′s F*&D*>0,中国安徽和越南分离株PvEBP-RII的中性检验值均为负值,所有研究地区分离株PvEBP-RII均有dN/dS>1。仅在缅甸的分离株中PvEBP-RII存在最小基因重组事件(Rm=1)(表2)。
2.3 PvEBP基因在不同群体间的分化
种群分化结果显示PvEBP基因在两两地区间均存在较高的遗传分化,且遗传差异均具有统计学意义(P<0.05),其中中国安徽与缅甸及越南间的遗传分化程度较大(Fst>0.15)(表3)。
2.4 不同群体间PvEBP基因进化关系
依据构建的系统发育树显示出不同地区PvEBP基因存在较为明显的地理聚类,各地区间序列相似性较小(图2-A)。单倍型网络图显示地理隔离导致的单倍型分布具有地区差异性,其中Hap_4是频率最高且所有群体共享的单倍型,除缅甸特有的单倍型Hap_7和Hap_11外,大部分单倍型之间仅有一步或两步突变。中国安徽仅有3个特有的单倍型(Hap_1,Hap_3,Hap_5),缅甸的单一单倍型数最多(图2-B)。

表2 分离株PvEBP-RII基因自然选择和基因重组分析

表3 各流行区间PvEBP基因的遗传分化程度
3 讨论
研究发现,裂殖子入侵网织红细胞是所有疟原虫血液阶段发育的关键,入侵配体家族一直是疫苗设计的靶标(Thametal., 2017)。基因多态性介导的疟原虫免疫逃逸是制约疫苗有效性的关键因素(Garzón-Ospinaetal., 2018)。尽管已有研究表明PvEBP可充当间日疟原虫入侵网织红细胞的替代配体,但遗传多样性和种群进化特征分析是评估PvEBP作为候选疫苗抗原的前提。本研究通过对我国中部安徽及周边国家缅甸和越南边境地区的PvEBP基因特征分析发现,各流行区PvEBP基因存在遗传差异,氨基酸突变位点主要集中在PvEBP-RII,提示此功能区为间日疟血液阶段疫苗研制的靶点。PvEBP基因在不同群体中呈现较大的遗传分化。同时出现在3个地区中的频率最高的单倍型(Hap_4)可能成为研制PvEBP疫苗的基础。
遗传多样性分析显示,与周边国家相比,我国安徽分离株PvEBP基因核苷酸多样性和单倍体数目(π=0.00051,Hd=0.634)相对较低。这可能是由于间日疟输入病例的阻断,使得我国中部省份间的基因流动受限造成的(Liuetal., 2014)。此外,也可能由于温带地区间日疟原虫休眠期较长,多克隆感染的发生率普遍较低,从而减少了基因重组的概率,降低了PvEBP基因多样性(Petersenetal., 2013)。缅甸和越南间日疟流行水平较高,且流动性较大(Brashearetal., 2020),使得它们的PvEBP基因多态性较高。
以往研究发现,PvDBP-RII区中的DBL结构域具有可介导交叉反应的表位,介导寄生虫粘附(Mitranetal., 2019),提示DBL结构域作为开发间日疟原虫入侵红细胞疫苗靶点的潜能(Singhetal., 2006)。本研究结果也显示,PvEBP-RII遗传多样性相对较高,氨基酸突变主要集中于PvEBP-RII,结合各流行区PvEBP-RII的选择压力分析结果(dN/dS>1),提示PvEBP-RII基因可能受到正向选择压力的影响。已有报道显示抗PvEBP-RII的抗体在间日疟感染患者中显著升高(Hanetal., 2020),进一步证实PvEBP-RII是免疫系统识别的部位,可成为间日疟血液阶段疫苗研制的合适靶标。
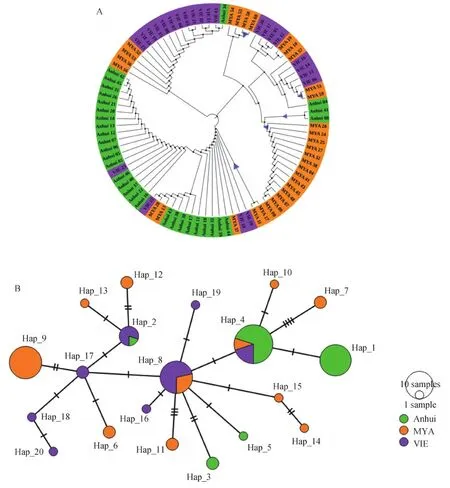
图2 群体间PvEBP基因的进化关系
系统发育树分析种群分化结果显示(图2-A),短时期内群体间的PvEBP基因交流不频繁,同一群体的分离株聚集明显且群体间序列相似性不高。同样,与PvEBP同源的诺氏疟原虫红细胞结合蛋白(PkEBP)在马来西亚两地区也表现出中等程度的遗传分化,提示地理隔离导致寄生虫不同的进化方向(Wangetal., 2019)。由于地理局限性导致极少的基因迁移和遗传漂变,PvEBP基因在不同群体中呈现较大的遗传分化(表3),应根据地区遗传特点开发更加有效的疟疾疫苗。本研究中,频率最高的单倍型(Hap_4)同时出现在3个不同流行区,提示该单倍体型具有地区适应性,以此为基础的疫苗在这3个流行区可能有效,然而,此单倍型的抗原性尚需进一步的实验验证。
