外源钙对荷花适应盐胁迫的影响
2020-03-07刘艺平苏少文黄志远孔德政
刘艺平,苏少文,张 琳,刘 莹,黄志远,贺 丹,孔德政
(河南农业大学 林学院,河南 郑州 450000)
盐碱土由于盐分含量高、结构差和缺乏养分而对植物生长产生不利影响,甚至导致植物死亡,这严重阻碍了农林业生产和土地绿化进程[1-2]。我国盐碱地总面积约占全国总面积的1/10[3],人口快速增加、城市化进程加速和耕地质量下降加剧了对土地资源的需求[4]。我国盐碱地面积大且分布广[5],选择适应盐碱地生长发育的植物对充分利用盐碱地和改善生态环境具有积极作用。
荷花(Nelumbonucifera)隶属睡莲科莲属,为多年生挺水草本植物,因其外形高雅圣洁、花色艳丽、花型丰富,观赏价值极高,被广泛应用于园林造景之中[6]。近年来,通过使用外源物质来缓解盐胁迫对植物的伤害,成为一种克服土壤盐渍化的途径。盐胁迫对植物造成的危害主要是离子毒害、渗透胁迫和膜透性改变,以及生理代谢紊乱[7]。CIPK家族基因在植物的生长发育和抗逆性中起着关键的调节作用。余爱丽等[8]从谷子[Setariaitalica(L.) P. Beauv.]基因组中鉴定出SiCIPK6基因,该基因在ABA、低温、高温、干旱和高盐诱导下表达量上调幅度较大,推测该基因在逆境胁迫中起到一定作用。He等[9]从棉花体细胞中克隆了GhCIPK6基因,将该基因转入拟南芥,转基因拟南芥表现出耐盐性和耐旱性,表明CIPK6在植物应激反应中起到双重作用。钙是植物必需的大量营养元素,在植物抗逆机制中起着重要作用[10]。研究表明,钙能提高植物耐盐[11]、耐低氧[12]、耐干旱[13]等多种抗性。有关外源钙对荷花盐胁迫下生理响应研究已有部分报道。印荔[14]对莲藕耐盐性进行了研究,认为Ca2+对盐胁迫下莲藕的生长具有较好的改善作用,证实NnCDPK13基因与其耐盐性相关。李淑艳等[15]研究表明,低浓度的Ca2+对缓解莲藕盐胁迫具有较好的效果,但Ca2+浓度增加时,缓解作用不明显,甚至出现抑制效果。本文通过研究盐敏感品种粉美人和盐抗性品种水晶粉七号在盐胁迫下对外源钙离子的生理响应,初步探究外源钙离子对盐胁迫下荷花生长的缓解效果,并分析与Ca2+相关的关键基因NnCIPK6基因的表达模式,为深入研究荷花耐盐分子机理和利用分子辅助育种途径改良荷花资源的耐盐性提供理论指导。
1 材料与方法
1.1 试验材料
试验中所用药剂氯化钙、氯化钠由天津市瑞金特化学品有限公司提供。荷花材料由河南省驻马店市荷花培植研究会(郑州基地)提供,品种为凤舞、粉美人、红楼、鲜娇、春红和水晶粉七号。分别选取各品种健康状况良好、生长势基本一致的植株用于盆栽试验。于2019年4月分别将6种幼苗栽植于橡胶盆内并施肥,以保证植株正常生长,每盆定植1株。在河南省优质花卉蔬菜种苗工程研究中心露天苗圃进行培育供试,雨天进行遮盖。
1.2 试验方法
将所有供试材料培养1个月后,选取生长健壮均匀的植株,进行NaCl处理。试验共设5个处理:A1,0 mmol·L-1NaCl;A2,50 mmol·L-1NaCl;A3,100 mmol·L-1NaCl;A4,150 mmol·L-1NaCl;A5,200 mmol·L-1NaCl。每个处理3个重复,共计15盆,处理7 d后进行盐害指数测定。
进行NaCl处理后选出抗性品种与敏感品种,再选取生长健壮均匀的植株,待长出3~5叶时,随后进行外源钙处理。采用盐浓度为100 mmol·L-1溶液培养,模拟盐胁迫环境。试验共设计6个处理:对照(CK)0 mmol·L-1NaCl+0 mmol·L-1CaCl2;T1,100 mmol·L-1NaCl+0 mmol·L-1CaCl2;T2,100 mmol·L-1NaCl+5 mmol·L-1CaCl2;T3,100 mmol·L-1NaCl+10 mmol·L-1CaCl2;T4,100 mmol·L-1NaCl+15 mmol·L-1CaCl2;T5,100 mmol·L-1NaCl+20 mmol·L-1CaCl2。试验采取随机区组设计,每个处理3个重复,共计18盆,处理14 d后结束试验。分别于处理后的第0、7和14天08:00测量生长量和采样,用电子天平称取0.2 g叶,用锡箔纸包住,放于液氮中,贮存在-80 ℃备用,用于测量各项生理生化指标。
1.3 测定方法
1.3.1 盐害指数测定
根据以下盐害级别判定标准[16-17]进行耐盐性评价:1级,生长正常,无明显盐害症状;2级,轻度盐害,少数叶片尖缘枯焦或黄化;3级,中度盐害,1/2叶片尖缘枯焦黄化或少量叶片脱落失水萎蔫;4级,重度盐害,大部分叶片枯焦,1/2左右叶片脱落;5级,极重度盐害,多数叶片脱落,枝条枯死,植株濒临死亡。
1.3.2 株高和叶面积测定
用尺子测量植株的株高、叶片长宽。净高生长量计算公式如下:净高生长量=处理后植株高度-原始植株高度。参考王永皎等[18]图像处理法计算植物叶面积。
1.3.3 生理指标的测定
叶绿素含量测定参考波钦诺克[19]和朱广廉等[20]的方法并加以改进。超氧化物歧化酶(SOD)活性的测定采用的氮蓝四唑(NBT)法。丙二醛含量的测定采用硫代巴比妥酸法。脯氨酸含量测定采用酸性茚三酮比色法[21]。
1.3.4 基因表达量的测定
采用改良的CTAB法提取RNA,以18S-rRNA为内参基因。上下游引物用Primer 5.0设计,引物由北京六合华大基因科技有限公司合成,引物序列如表1所示。

表1 引物序列Table 1 Primers sequence
采用SYBR Green I染料法,在200 μL PCR管中,于冰上依次加入试剂:10.0 μL SYBR Premix ExTaqII、1.0 μL 上游引物、1.0 μL 下游引物、0.4 μL ROX II、1.0 μL反转录产物和6.6 μL ddH2O,总反应体系共20 μL。PCR扩增程序:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 34 s,40个循环。
1.4 数据处理
数据采用Excel软件进行初步分析,并用SPSS 19.0软件进行方差分析并分析其差异显著性,使用Excel和Origin Pro8绘制各项指标变化图。
2 结果与分析
2.1 NaCl胁迫下荷花的盐害指数
由表2可以看出,50 mmol·L-1NaCl胁迫下,粉美人盐害指数最高,说明其受害表现最为明显;其次是凤舞和春红,水晶粉七号未表现出受害现象。100 mmol·L-1NaCl胁迫下,红楼的受害指数增长最快,与粉美人相近。150 mmol·L-1NaCl胁迫下,所有品种的盐害指数均大幅度上涨,这说明150 mmol·L-1NaCl对荷花生长状况有明显影响,植株明显受到伤害。当NaCl浓度升高到200 mmol·L-1时,所有品种的盐害指数为17.33%~28.00%,粉美人盐害指数最大,达到28.00%。总体来看,水晶粉七号耐盐性最强,鲜娇和春红次之,粉美人最低。各品种耐盐能力为水晶粉七号>鲜娇>春红>红楼>凤舞>粉美人。
2.2 外源钙对NaCl胁迫下荷花生长指标的影响
2.2.1 净高生长量
由图1可看出,NaCl胁迫不同程度上抑制了盐敏感品种粉美人和盐抗性品种水晶粉七号的生长,胁迫7 d和14 d后,盐敏感品种和盐抗性品种的净高生长量与对照组相比均减小,敏感品种在T1处理下的净高生长量分别是4.23、6.87 cm,而抗性品种的净高生长量分别是4.87、7.43 cm,抗性品种水晶粉七号的净高生长量显著(P<0.05)大于敏感品种粉美人。经过不同浓度的CaCl2处理后,2种荷花的高生长量均大于T1处理,这说明不同浓度的CaCl2处理对缓解盐胁迫有一定的作用,其中以10 mmol·L-1CaCl2处理的缓解效果最好。抗性品种在盐胁迫7 d和14 d后,T3处理下净高生长量达到9.50 cm和15.57 cm,而盐敏感品种在盐胁迫7 d和14 d后,T3处理下净高生长量达到8.32 cm和14.40 cm,且差异均显著(P<0.05)。由此可以看出,盐胁迫对荷花的危害随胁迫时间的延长而加重。
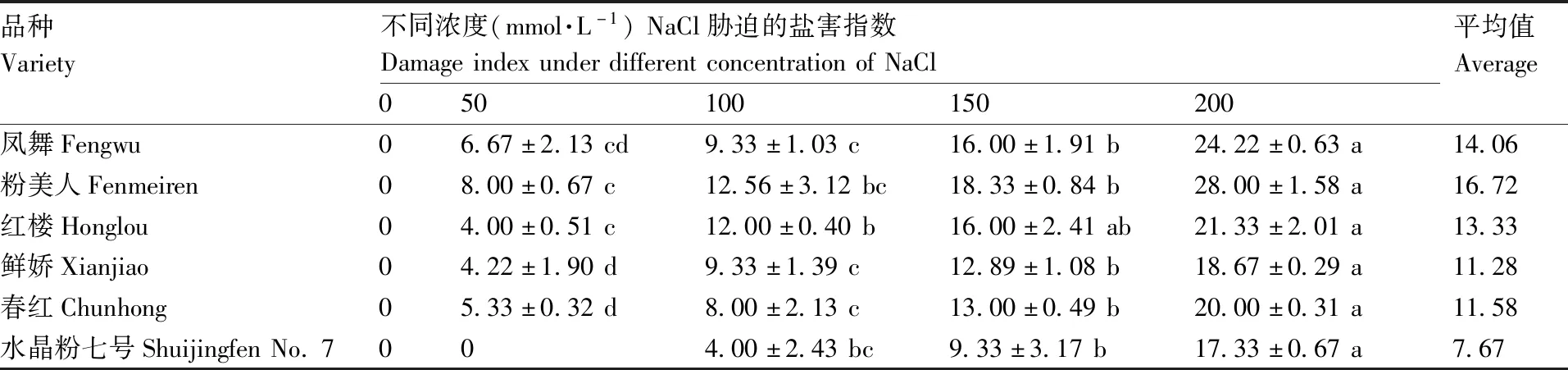
表2 NaCl胁迫下荷花的盐害指数Table 2 Salt damage index of lotus under stress
同行数据后无相同小写字母表示在P<0.05水平上差异显著。
Data marked without the same lowercase letter in each row indicated significant differences atP<0.05.

图1 外源CaCl2对盐胁迫下荷花净高生长量的影响Fig.1 Effects of exogenous CaCl2 on the clear height growth of lotus under salt stress

柱上无相同小写字母表示差异显著(P<0.05)。下同。Data on the bars marked without the same lowercase letter indicated significant differences at P<0.05. The same as below.图2 外源CaCl2对盐胁迫下敏感品种和抗性品种叶面积的影响Fig.2 Effects of exogenous CaCl2 on leaf area of sensitive variety and tolerant variety lotus under salt stress
2.2.2 叶面积
由图2可看出,NaCl胁迫对荷花的叶面积增长也产生了一定影响。敏感品种对照组在7 d和14 d后叶面积分别增长7.29%和14.22%,抗性品种对照组在7 d和14 d后叶面积分别增长7.46%和14.45%。T1处理下,敏感品种在7 d和14 d后叶面积分别增长2.75%和5.49%,抗性品种叶面积分别增长3.49%和6.43%,无论是正常条件下还是盐胁迫下抗性品种水晶粉七号的叶面积增长量均显著(P<0.05)大于敏感品种粉美人。在T3处理中,敏感品种在盐胁迫7 d和14 d后叶面积分别增长5.57%和10.28%,抗性品种在盐胁迫7 d和14 d后叶面积分别增长4.95%和9.66%,这说明在外源钙处理下,敏感品种粉美人的叶面积增长量大于抗性品种水晶粉七号,表明在盐胁迫条件下外源钙可能对敏感品种粉美人的缓解效果更好。
2.3 外源钙对NaCl胁迫下荷花生理生化指标的影响
2.3.1 叶绿素含量
从表3可以看出:T1处理中,7 d后敏感品种粉美人和抗性品种水晶粉七号的叶绿素a(Chl a)、叶绿素b(Chl b)和叶绿素总量(Chla+Chlb)含量分别为21.22、7.32、28.51 mg·g-1和25.37、10.21、31.64 mg·g-1,比对照组分别下降18.69%、40.95%、25.87%和7.46%、31.12%、22.73%,盐胁迫下荷花叶绿素含量明显降低,敏感品种粉美人的降幅更大。T3处理中,与处理7 d相比,处理14 d荷花的叶绿素含量有一定的下降,敏感品种处理Chl a、Chl b和Chla+Chlb分别下降4.67%、12.37%和4.51%,抗性品种Chl a、Chl b和Chla+Chlb分别下降8.66%、15.44%和2.69%。
2.3.2 SOD活性
图3表明:T1处理中,7 d和14 d 抗性和敏感品种荷花的SOD活性与对照组相比显著升高,敏感品种的SOD活性分别达到241.49、293.72 U·g-1,比对照组分别增加了5.07%和13.68%,表明敏感品种粉美人的SOD活性随着盐胁迫时间的延长增大;T1处理中抗性品种的SOD活性在7 d和14 d分别达到了278.37 U·g-1和294.35 U·g-1,比对照组分别增加了1.44%和2.79%。胁迫14 d后,T1处理组敏感品种的SOD活性显著(P<0.05)高于对照组和抗性品种。4个不同浓度的CaCl2处理后,2种荷花的SOD活性均有所升高,10 mmol·L-1和15 mmol·L-1CaCl2处理的SOD活性升高的幅度最大。其中T3处理7 d和14 d后,敏感品种和抗性品种的SOD活性分别比T1处理升高了26.42%、18.24%和13.89%、16.83%。

表3 外源CaCl2对盐胁迫下荷花叶绿素含量的影响Table 3 Effects of exogenous CaCl2 on chlorophyll content of lotus under salt stress
同列数据后无相同小写字母表示差异显著(P<0.05)。
Data marked without the same lowercase letter in each column indicated significant differences atP<0.05.

图3 外源CaCl2对盐胁迫下荷花SOD活性的影响Fig.3 Effects of exogenous CaCl2 on SOD activity of lotus under salt stress

图4 外源CaCl2对盐胁迫下荷花MDA含量的影响Fig.4 Effects of exogenous CaCl2 on MDA content of lotus under salt stress
2.3.3 丙二醛含量
由图4可知:在T1处理下,粉美人和水晶粉七号MDA含量最高,与对照组相比,粉美人在7 d和14 d后MDA含量分别增长69.68%和36.92%,水晶粉七号在7 d和14 d后MDA含量分别增长59.19%和47.84%。T3处理7 d和14 d粉美人MDA含量分别达到5.74、8.42 μmol·g-1,与对照组相比分别增长23.44%和9.07%,分别比T1处理下降27.25%和20.34%;T3处理7 d和14 d水晶粉七号MDA含量分别达到4.39、7.25 μmol·g-1,与对照组相比分别增长18.65%和16.00%,比T1处理分别下降25.47%和21.54%。
2.3.4 脯氨酸含量
由图5可知:T1处理中,7 d和14 d粉美人的脯氨酸含量分别比对照组增加20.02%和19.55%,水晶粉七号的脯氨酸含量分别比对照组增加38.71%和14.77%,处理7 d后水晶粉七号脯氨酸含量的增幅明显大于粉美人。随着外源CaCl2浓度的升高,不同处理下的脯氨酸含量均有不同程度的升高,T3和T4处理中脯氨酸含量的积累速率加快。

图5 外源CaCl2对盐胁迫下荷花脯氨酸含量的影响Fig.5 Effects of exogenous CaCl2 on Proline content of lotus under salt stress

图6 外源CaCl2对盐胁迫下荷花NnCIPK6表达量的影响Fig.6 Effects of exogenous CaCl2 on NnCIPK6 gene expression of lotus under salt stress
2.4 外源钙对NaCl胁迫下荷花NnCIPK6基因表达量的影响
从图6可以看出,与对照组相比,盐胁迫下荷花NnCIPK6基因表达量明显上升,加入CaCl2后NnCIPK6表达量进一步升高。处理7 d,随着CaCl2浓度的升高,粉美人和水晶粉七号的NnCIPK6表达量均表现出先升高后降低的趋势;T3处理的NnCIPK6表达量最高,水晶粉七号的NnCIPK6表达量是对照组的2.19倍,粉美人的NnCIPK6表达量是对照组的2.02倍;CaCl2浓度超过10 mmol·L-1后,NnCIPK6表达量呈现下降趋势,其中T5处理的抗性品种水晶粉七号NnCIPK6表达量显著(P<0.05)低于T3、T4处理。处理14 d后,2个荷花品种NnCIPK6表达量的变化趋势和处理7 d较为相似,但整体低于7 d的表达量;T3处理中,敏感品种粉美人和抗性品种水晶粉七号NnCIPK6基因表达量比处理7 d分别下降19.44%和28.92%。从图6可知,T3处理中NnCIPK6表达量最高,表明10 mmol·L-1CaCl2能缓解盐胁迫对荷花生长发育的伤害,粉美人NnCIPK6表达量均低于水晶粉七号,说明CaCl2对粉美人的缓解效果更好,此结果与荷花生长发育指标呈现的结果相符。
3 结论与讨论
在不同浓度的NaCl处理下,荷花的净生长量和叶面积均显著降低,并且随着NaCl处理时间延长,盐胁迫对荷花的危害加重,抗性品种水晶粉七号和敏感品种粉美人对盐胁迫的反应有显著差异。NaCl处理下,添加外源钙使荷花叶绿素含量升高,一定浓度下SOD活性和脯氨酸积累量达到最高,一定浓度的外源CaCl2处理能降低MDA含量,而高浓度的Ca2+也会对植物造成胁迫,即离子毒害,使膜脂过氧化程度和MDA含量增加。NaCl处理下,添加外源钙使抗性品种水晶粉七号和敏感品种粉美人的NnCIPK6基因表达量均显著升高,且在10 mmol·L-1CaCl2处理时最高,表明10 mmol·L-1CaCl2的缓解效果最好。
盐胁迫会使植物叶绿素含量降低,导致光合作用、CO2吸收能力降低,增加引起光系统氧化损伤的活性氧的含量[22-23]。本研究通过外源钙处理显著提高了盐胁迫下荷花的叶绿素含量,推测外源钙可抑制盐胁迫下荷花叶片叶绿素含量的下降,这与王鸿蕉等[22]在白菜 (Brassicarapavar.pekinensis)幼苗上得出的结论相一致。同时外源钙对敏感品种粉美人叶绿素含量下降的抑制效果大于抗性品种水晶粉七号,与颜志明[25]在甜瓜(CucumismeloL.)幼苗上得出的结果一致。这可能是因为敏感品种粉美人更容易对周围环境的变化做出反应,外源钙可能降低了盐胁迫下植物体中叶绿素酶的活性,抑制了因盐胁迫引起的叶绿素的降解,从而提高了盐胁迫下幼苗叶片叶绿素含量[26-27]。
在各种非生物胁迫下,活性氧(ROS)的产生和清除之间的平衡被打破,导致细胞内ROS的水平迅速增加,对细胞结构造成严重伤害[28]。SOD是细胞内有效的抗氧化酶,它能够将O2-转化为H2O2。逆境条件下,植物超氧化物歧化酶(SOD)被激活,以清除ROS的过度生成,保护植物细胞免受ROS的伤害。抗氧化酶活性测定表明,抗性品种水晶粉七号的SOD活性高于敏感品种粉美人。这一结果符合在其他植物物种中获得的结果,即盐胁迫提高抗氧化酶活性和促进活性氧的产生[29]。
MDA是膜脂过氧化的主要产物,其含量与细胞膜损伤程度呈正相关[30]。本试验中NaCl处理下荷花叶片MDA含量升高,表明盐胁迫使植物细胞膜脂过氧化作用增强,加剧对其的氧化损伤。适宜浓度的外源钙处理能有效地降低荷花MDA含量,但仍高于对照;且抗性品种水晶粉七号的MDA含量低于敏感品种粉美人。这与Sarmast等[31]在高羊茅(Festucaarundinacea)上得出的结论一致,即在干旱胁迫下MDA含量相对较低的品种比MDA含量较高的品种具有更好的抗逆性。
脯氨酸可维持细胞的扩张和渗透调节功能,以提高植物对渗透胁迫的耐受性[32],脯氨酸积累与植物胁迫耐受性的改善相关[31]。本试验结果表明,盐胁迫下荷花脯氨酸含量升高,抗性品种水晶粉七号的脯氨酸含量高于敏感品种粉美人,这与Bojórquez-Quintal等[33]在辣椒(CapsicumannuumL.)上得出的结论一致。
CIPK作为CBL的唯一特异性靶蛋白激酶,是目前发现的植物中所特有的一类具有丝氨酸/苏氨酸激酶活性的蛋白激酶家族[34]。CIPK家族基因在植物的抗逆性中起着关键的调节作用。Wang等[35]研究发现,大多数CIPK家族基因在低温、干旱和盐胁迫处理后的棉花叶、茎和根中表达量显著增加;Ma等[36]研究表明,过表达CBL和CIPK基因能显著提高植物抗盐胁迫能力,验证了CIPK家族基因能够在非生物刺激中发挥重要作用。目前CBL和CIPK基因功能在模式植物拟南芥中的研究较为深入,主要集中在揭示CBL或CIPK基因抵抗生物或非生物胁迫的分子机制,尤其以CBL-CIPK介导的Ca2+信号在盐胁迫、低温、干旱和ABA等刺激中的作用机制研究较多[37]。在拟南芥中,AtCIPK6基因是幼苗发育和盐胁迫反应所必需的[38]。近些年,从苹果(PumilaMill.)[39]、棉花(GossypiumhirsutumL.)[40]中克隆了CIPK6基因,结果显示,这些基因受盐胁迫诱导并参与转基因植物对多种非生物胁迫的响应。本研究结果也表明,NaCl处理后荷花NnCIPK6的表达水平明显升高,说明NnCIPK6在荷花应对盐胁迫的反应中起作用,但其具体功能有待进一步研究。
