天然香料植物油樟叶可培养内生细菌群落结构与多样性
2019-11-14周万海杨立艳李苗黄金凤梁玉娟王鑫罗思灿魏琴刘洋
周万海,杨立艳,李苗,黄金凤,梁玉娟,王鑫,罗思灿,魏琴*,刘洋*
1(宜宾学院,香料植物资源开发与利用四川省高校重点实验室,四川 宜宾,644000)2(北京科技大学 化学与生物工程学院,北京,100083)
植物内生细菌是指一定时间或全部时间定殖于植物体内对宿主无明显不利影响的细菌[1],是植物微生态系统中的重要组成部分。近年来,人们已经从水稻[2]、葡萄[3]、巨菌草[4]等不同生境的多种植物分离获得大量内生细菌,发现几乎所有的植物体内都存在内生细菌,这些内生菌在与宿主共生过程中,可通过为宿主植物提供所需的部分营养物质和激素来调控植物的生长发育[5-6],亦可合成和分泌部分抗菌素和酶等活性物质进而增强植物对环境的适应能力[7-8]。同时还广泛参与宿主次生代谢产物的合成与转化,对宿主植物的种群结构、生存、进化和健康状态产生重大影响[9];植物内生细菌在农业生产、天然产物合成、生物医药、发酵工程、环境保护和生物防治等方面有巨大应用潜力[10],已成为研究的热点。
作为食品添加剂家族重要成员的香料香精在食品工业生产中发挥着难以估量的作用,然而近些年来食品安全问题受到空前关注,符合“绿色”要求的天然香料香精的需求不断上升,与之相关研究也非常活跃[11]。油樟(Cinnamomumlongepaniculatum(Gamble) N. Chao)是樟科樟属常绿乔木植物,为我国特有的产天然芳香油的主要树种之一,其体内富含的1,8-桉叶油素、松油烯-4-醇、α-松油醇等重要化学成分,是生产香料香精、食品、调味品的重要原料,在医药制品、食品添加剂、日用化工的生产中具有广阔的开发应用前景[12]。位于四川省南部的宜宾市,油樟种植面积已超过50万亩,占全国油樟总资源的75%以上,且与同属植物种相比,宜宾油樟芳香油含量更高,约为广东、江西、台湾等省油樟含芳香油量的2.5倍,这可能与其共生微生物等特殊环境条件有关[13],大量研究证实由于宿主植物(基因型)的差异及受分布地域、温度、湿度、季节变化、气候条件等环境因素影响,植物内生细菌的数量和种类存在丰富多样性[14],且在不同宿主及不同器官的种类、数量、群落结构和多样性等方面存在较大的差异[15]。特别是一些高树龄的植物、在特殊环境下生存的植物和未受人为活动干扰的地区可能蕴含更多的特殊内生菌类群;宜宾作为全国油樟最集中的分布区域,该地区生长的油樟可能存在与其他地区、其他物种完全不同的潜在内生菌资源。同时受植物内生菌在次生代谢物合成和转化的启示,香料植物内生菌在天然香料的合成及制备中的作用及应用也成为研究热点[16]。因此,分离和研究油樟内生菌,对了解和开发本区域油樟生产性能极为重要;然而,目前对油樟的研究主要集中在苗木繁育、樟油提取、单体分离和提取物功能特性等方面[17-19],关于油樟内生菌,多集中在内生真菌,如王涛等从油樟根茎叶中分离到104株内生真菌,并对其抑菌性能进行分析[20],游玲等对78株油樟内生真菌发酵产物进行了分析[21],冯瑞章等则对油樟内生溶磷细菌进行了筛选和分析[22],而对油樟内生细菌的研究几乎未见相关报道,因此,本研究以宜宾地区不同生长季节油樟叶为材料,采用本实验室筛选的叶片研磨加中和剂的方法,研究叶片内生细菌的分布情况与多样性,以期为后续油樟内生细菌的深度挖掘与解析宜宾油樟独特产油特性奠定基础。
1 材料与方法
1.1 样品采集及处理
样品采集时间分别为2018年6月、2018年9月、2018年12月和2019年3月,采样地点为四川省宜宾市高县月江森林经营所油樟母本园;采样时选取成年健康植株5株,根据随机取样原则,取样的株距在30~50 m,在树冠的上、中、下3个部位随机采取枝条上完全展开的叶片各5片,采集后混合均匀装入无菌封口塑料袋,立即带回实验室,并于采样后24 h内启动样品内生细菌的分离。
1.2 培养基及试剂
根据前期预实验结果,分离培养基选用牛肉膏蛋白胨培养基(LB)、大豆酪蛋白琼脂培养基(TSA)、阿须贝氏培养基和卵磷脂吐温80营养琼脂培养基;肉汁胨培养基(NA)用于内生菌的活化和保存。
由于油樟叶片含有大量黏性物质,严重抑制内生菌的分离,本研究根据前期实验结果,采用中和剂处理研磨叶片,其配方为:0.5%(质量分数)甘氨酸,3%(质量分数)卵磷脂吐温80,磷酸缓冲溶液pH 7.2。
Ezup柱式细菌基因组DNA抽提试剂盒、细菌的16S rDNA通用引物27F(5′-AGAGTTTGATAGAGTTTGATCCTGGCTCAG-3′)和1492R(5-GGTTACCTTGTTACGACTT-3′)、PCR反应试剂,均由生工生物工程(上海)股份有限公司合成和提供。
1.3 油樟叶片表面灭菌
挑选采集样本中无病变、品相良好的油樟叶片5~10 g,先用流水冲洗1~2 h,无菌水冲洗干净后,用70%乙醇浸泡2 min,再用3%次氯酸钠溶液浸泡5 min,最后用无菌水冲洗4次。取最后1次冲洗液0.1 mL涂布到LB培养基平板上,28 ℃恒温培养箱中培养72 h,检测表面消毒效果。
1.4 内生细菌的分离与纯化
油樟叶内生细菌群落多样性研究采用传统分离培养技术,取1.0 g消毒后的材料置于无菌研钵中磨成粉末(加入5 mL无菌水和4 mL中和剂混合研磨成匀浆),采用梯度稀释法制备1×10-1~1×10-3g/mL的系列稀释液,每一梯度稀释液分别取0.2 mL涂布于LB、TSA、阿须贝氏培养基和卵磷脂吐温80营养琼脂培养基平板上,重复3次,置于28 ℃恒温培养3~5 d,待培养基中长出的菌落数平稳后,根据各平板上菌落的形态(大小、颜色、形状、表面光泽度、透明度、边缘整齐度等)随机挑取具有代表性单菌落[23],纯化编号后4 ℃保存于NA斜面上备用。根据菌落数计算每克叶片中的内生细菌总数,同时利用新复极差法进行显著性分析。
1.5 内生细菌的16S rDNA序列分析
用细菌基因组DNA提取试剂盒(生工生物)提取细菌基因组DNA。采用通用引物27F和1492R扩增16S rDNA。PCR反应体系(25 μL): Taq PCR Master Mix(2x)12.5 μL、DNA模板1 μL、正向引物(10 μmol/L)1 μL、反向引物(10 μmol/L)1 μL、ddH2O补足至25 μL;PCR反应条件:94 ℃、4 min、30个循环,94 ℃、30 s,55 ℃、30 s,72 ℃、90 s,72 ℃、10 min。PCR产物用1.5%琼脂糖凝胶电泳进行检测。扩增产物切胶纯化后送上海生工生物进行序列测定,将所有序列信息提交到NCBI,测序得到的结果在EzTaxon server 2.1进行比对,确定与已知序列同源关系,比对后序列相似性达到98.65%以上的归为同一个运算的分类单位(operational taxonomic units,OTU),并将代表菌株序列信息提交到GenBank并获得登录号。同时选择和下载近缘种序列,用MEGA 6软件Neighbour-Joining法构建系统进化树[24]。
1.6 内生细菌群落多样性分析
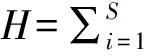
2 结果与分析
2.1 不同季节油樟叶片可培养内生细菌量变化
平板分离计数结果显示,不同季节油樟叶可培养内生细菌总数存在显著差异,夏季油樟叶内生细菌总数为1.4×103CFU/g,而冬季油樟叶中内生细菌总数为0.7×103CFU/g,显著低于夏季油樟叶(P<0.05),是内生菌丰度最低的生长阶段;油樟叶内生菌的丰度最大值出现在秋季,达到4.5×104CFU/g,其次为春季的1.65×104CFU/g,二者之间存在显著差异,但都显著高于夏季和冬季。
2.2 不同季节油樟叶片中可培养内生细菌分子鉴定
对传统分离培养方法获得的全部菌株通过形态观察,最终挑取166株细菌进行16S rDNA序列的测定,通过在相关公共数据库中进行相似性搜索,并调出相关典型菌株的有效序列,用相关软件进行序列比对、相似性计算、进化距离矩阵计算、聚类分析和系统进化树构建(图1~图4)。将代表菌株序列信息提交到GenBank并获得登录号。
挑取的166株内生细菌中,春夏秋冬4个季节分别有94、56、10和6株,对应有23、4、11和2个OTU(表1~表4),表明不同季节油樟叶可培养内生菌的数量和OTU数目发生显著变化,其中春季油樟叶OTU数目最多,秋季油樟叶的OTU数目次之,而冬季油樟叶的OTU数目最少,仅为2个。
春季油樟叶分离的94株内生细菌隶属于3门9属(图1),其中变形菌门(Proteobacteria)有不动杆菌(Acinetobacter),假单胞菌(Pseudomonas)和鞘酯单孢菌(Sphingomonas)3个属,厚壁菌门(Firmicutes)有葡萄球菌(StaphyLococcus)、芽孢杆菌(Bacillus),气球菌(Aerococcus)和赖氨酸芽孢杆菌(LysinibaciLLus)4个属;放线菌门(Actinobacteria)仅有考克氏菌(Kocuria)和罗氏菌(Rothia)2个属。春季油樟叶中的优势种为Bacillusamyloliquefaciensstrain YC16 (MK713569.1),共计51株,丰度达到56.1%(表1)。

图1 基于16S rDNA序列构建的春季油樟叶内生细菌系统发育树Fig.1 Neighbor-joining phylogenetic tree of endophyticbacteria in Cinnamomum longepaniculatum (Gamble)N. Chao leaves at spring based on 16S rDNA gene sequences
夏季油樟叶分离的10株内生细菌分别隶属于厚壁菌门(Firmicutes)的葡萄球菌属(Staphylococcus)、芽孢杆菌属(Bacillus)、微杆菌属(Microcoli)和微球菌属(Micrococcusgenera)(图2),Microbacteriumresistensstrain PL-24(MK517595.1)为第一优势种,第二和第三优势种分别为Bacillussp. (in:Bacteria)strain H2(MK704289.1)和Micrococcussp. strain MRDDd(MK578275.1),Staphylococcusnepalensisstrain MFB1(MK602316.1)为第4优势种(表2)。
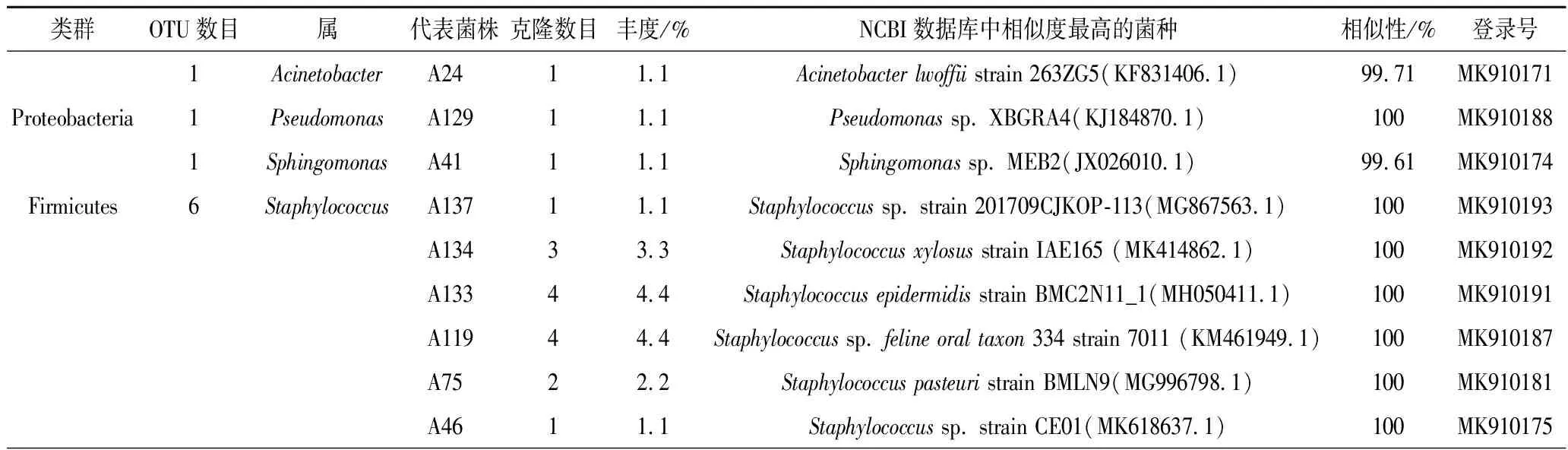
表1 春季油樟叶内生细菌分布Table 1 Distribution of endophytic bacteria in Cinnamomum longepaniculatum(Gamble) N. Chao leaves at spring
续表1
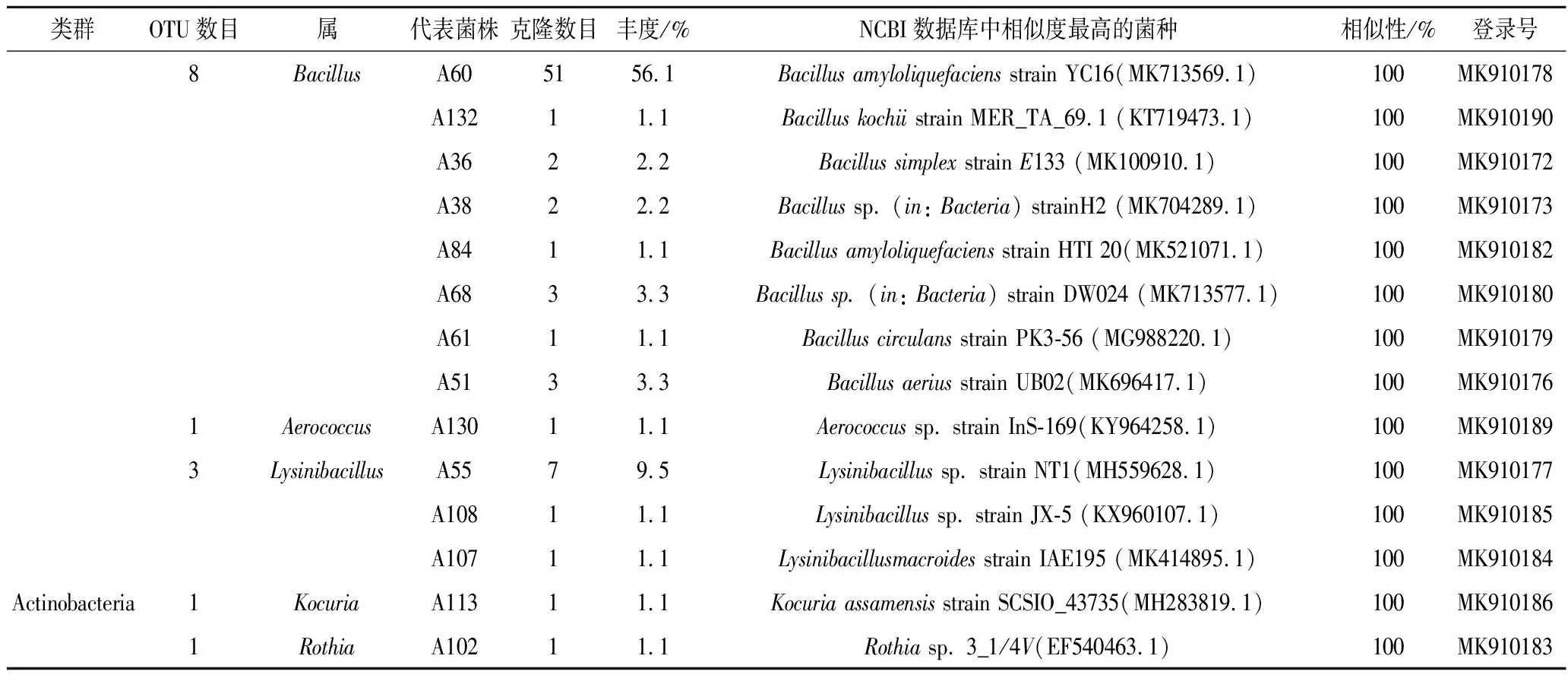
类群OTU数目属代表菌株克隆数目丰度/%NCBI数据库中相似度最高的菌种相似性/%登录号8BacillusA605156.1Bacillus amyloliquefaciens strain YC16(MK713569.1)100MK910178A13211.1Bacillus kochii strain MER_TA_69.1 (KT719473.1)100MK910190A3622.2Bacillus simplex strain E133 (MK100910.1)100MK910172A3822.2Bacillus sp. (in: Bacteria) strainH2 (MK704289.1)100MK910173A8411.1Bacillus amyloliquefaciens strain HTI 20(MK521071.1)100MK910182A6833.3Bacillus sp. (in: Bacteria) strain DW024 (MK713577.1)100MK910180A6111.1Bacillus circulans strain PK3-56 (MG988220.1)100MK910179A5133.3Bacillus aerius strain UB02(MK696417.1)100MK9101761AerococcusA13011.1Aerococcus sp. strain InS-169(KY964258.1)100MK9101893LysinibacillusA5579.5Lysinibacillus sp. strain NT1(MH559628.1)100MK910177A10811.1Lysinibacillus sp. strain JX-5 (KX960107.1)100MK910185A10711.1Lysinibacillusmacroides strain IAE195 (MK414895.1)100MK910184Actinobacteria1KocuriaA11311.1Kocuria assamensis strain SCSIO_43735(MH283819.1)100MK9101861RothiaA10211.1Rothia sp. 3_1/4V(EF540463.1)100MK910183

表2 夏季油樟叶内生细菌分布Table 2 Distribution of endophytic bacteria in Cinnamomum longepaniculatum(Gamble) N. Chao leaves at summer

图2 基于16S rDNA序列构建的夏季油樟叶内生细菌系统发育树Fig.2 Neighbor-joining phylogenetic tree of endophyticbacteria in Cinnamomum longepaniculatum (Gamble)N. Chao leaves at summer based on 16S rDNA gene sequences
秋季油樟叶分离的56株内生细菌隶属于3门5属(图3),包括变形菌门(Proteobacteria)的不动杆菌属(Acinetobacter);厚壁菌门(Firmicutes)的芽孢杆菌(Bacillus),葡萄杆菌(Staphylococcus)和病毒性杆菌(Virgibacillus)3个属,拟杆菌门(Bacteroidetes)仅1个属为金杆菌属(Chryseobacterium)(表3)。其中Bacillussiamensisstrain TSS18(MF620076.1)和Bacilluscereusstrain X-7(MF988730.1)为并列第1优势种,Bacillussubtilisstrain HZ-72(MF136610.1)为第2优势种。

图3 基于16S rDNA序列构建的秋季油樟叶内生细菌系统发育树Fig.3 Neighbor-joining phylogenetic tree of endophyticbacteria in Cinnamomum longepaniculatum (Gamble)N. Chao leaves at autumn based on 16S rDNA gene sequences
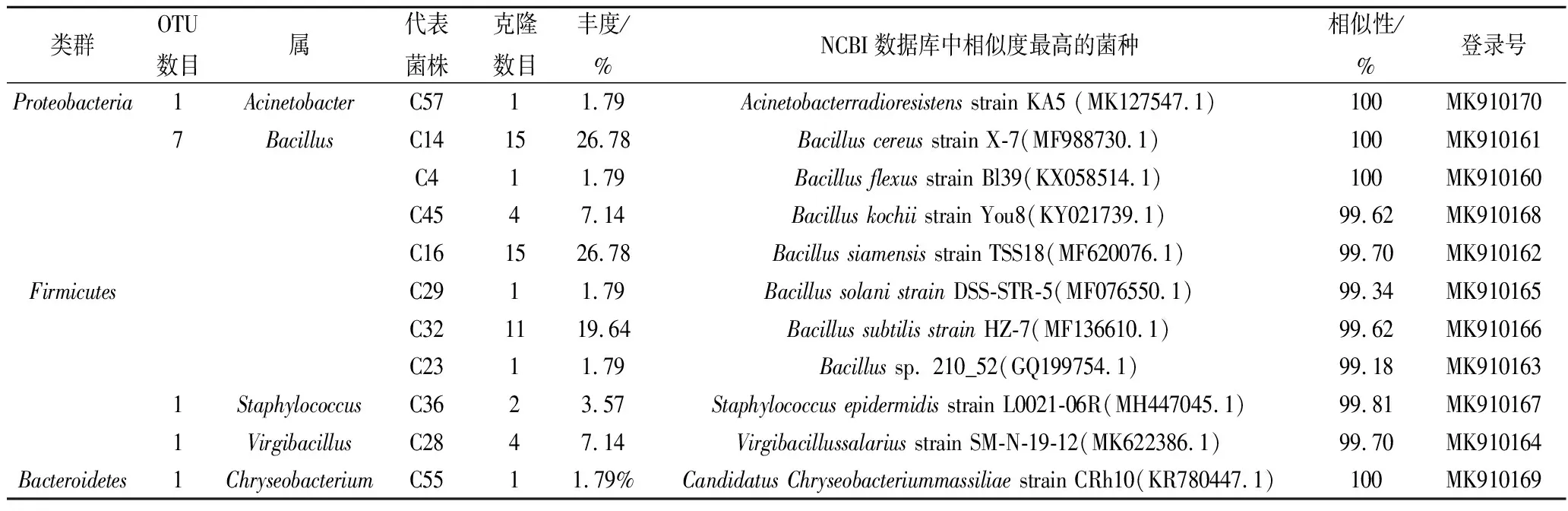
表3 秋季油樟叶内生细菌分布Table 3 Distribution of endophytic bacteria in Cinnamomum longepaniculatum(Gamble) N. Chao leaves at autumn
冬季油樟叶分离的6株内生菌均隶属于厚壁菌门(Firmicutes)芽孢杆菌属(Bacillus)(图4),其第1优势种为Bacillusaltitudinisstrain LZLJ004(KR018737.1),丰度达83.33%,第2优势种为芽孢杆菌属(Bacillus)为Bacillussubtilisstrain JPC-2(EU1 53188.1),丰度为16.67%(表4)。

图4 基于16S rDNA序列构建的冬季油樟叶内生细菌系统发育树Fig.4 Neighbor-joining phylogenetic tree of endophyticbacteria in Cinnamomum longepaniculatum (Gamble)N. Chao leaves at winter based on 16S rDNA gene sequences

表4 冬季油樟叶内生细菌分布Table 4 Distribution of endophytic bacteria in Cinnamomum longepaniculatum (Gamble) N. Chao leaves at winter
此外由以上分析可知,油樟叶中可培养内生菌的数量和种类均表现为春、秋两季较丰富,冬、夏两季较单一,且4个季节油樟叶中内生细菌的优势种发生显著变化,但第1优势种的属不变,都为芽孢杆菌属(Bacillus)(图5)。
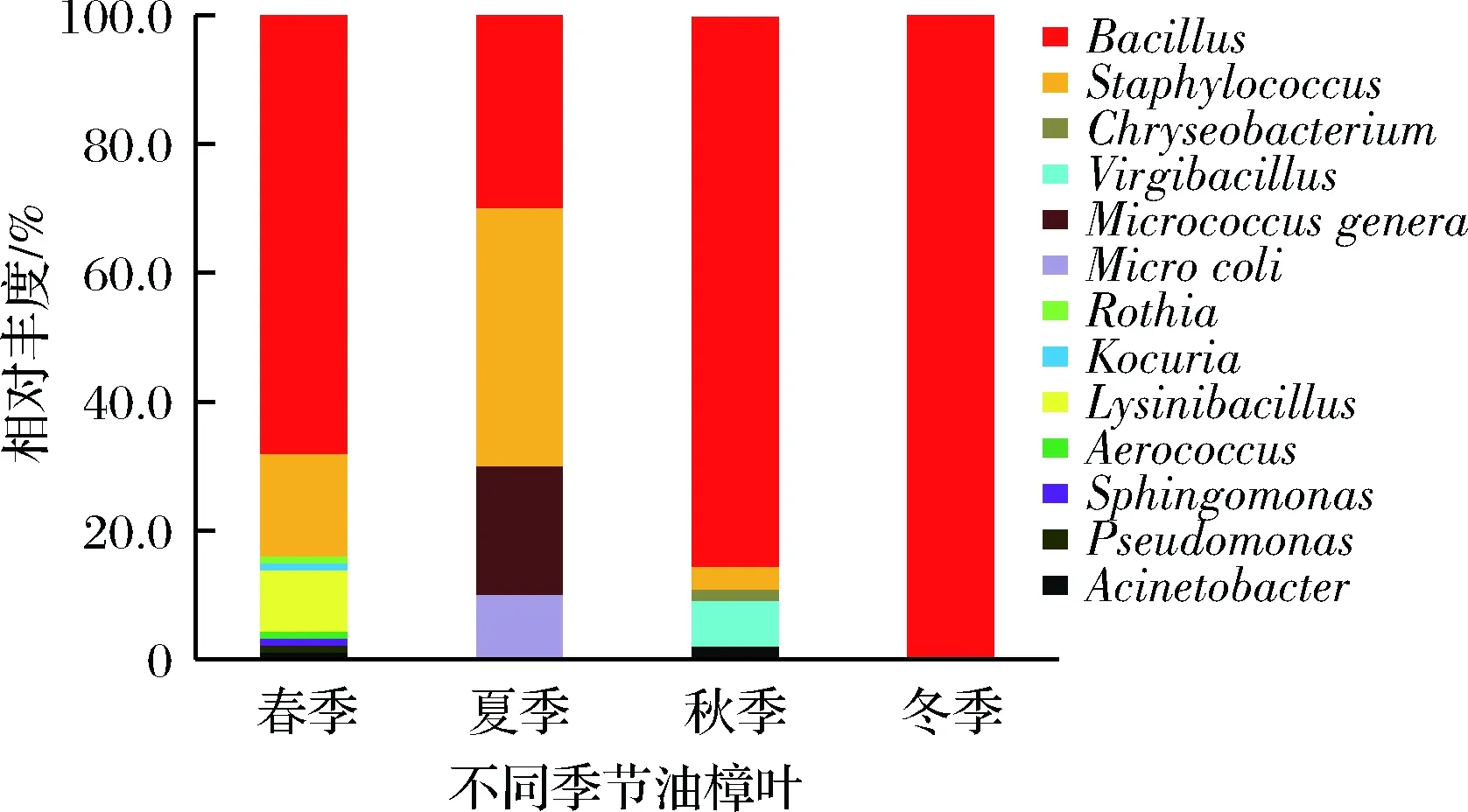
图5 不同季节油樟叶可培养内生细菌种类丰度变化Fig.5 Relative abundance of endophytic bacteriumgenera in Cinnamomum longepaniculatum(Gamble) N. Chao leaves at different seasons
2.3 不同季节油樟叶片中可培养内生细菌的多样性
量化的多样性指数可反映植物生长过程中内生菌群落结构的演替变化规律,如物种丰富度(S)是衡量物种多样性的最直接也是最重要的尺度;Shannon-Wienner指数(H)可以解释物种的丰度和均匀度,也可以解释物种的多样性,物种数量越多,分布越均匀,其数值越大;Simpso指数(D)表示该群落优势度的统计量,其值越大表明群落的优势种越明显,它随一个或几个物种优势度的增加而增加;群落组成的数量及空间分布的不同,形成群落的结构格局[27]。油樟生长过程中不同季节叶内生细菌的丰富度和多样性指数见表5,由表5可以看出,在4个不同季节油樟叶中,物种丰富度指数和Shannon-Wienner指数值反映出基本一致的多样性趋势,其变化分别为2~23和0.11~2.00,变化较为剧烈,但都表现为春季最大,冬季最小;不同季节油樟叶可培养内生细菌Simpson指数值变化也极为剧烈,其中秋季值最大,冬季值最小;而不同季节油樟叶内生细菌均匀度指数的变化在0.15~0.92,表现为夏季>秋季>春季>冬季,由此可见,春季油樟叶中内生细菌群落的物种数最多,多样性最为丰富,秋季次之,而冬季油樟叶内生细菌群落的物种数最少,多样性也最低。

表5 不同季节油樟叶中可培养内生细菌多样性指数Table 5 Diversity of endophytic bacteria in Cinnamomum longepaniculatum(Gamble) N. Chao leaves at different seasons
3 讨论
内生菌是影响植物生长发育和产品品质的重要因素,是潜力巨大且尚待开发的微生物新资源[14]。为了得到能够用于纯培养并能进一步研究、应用的植物内生细菌,本研究采用传统的分离培养方法首次对不同季节油樟叶中内生细菌进行分离、纯化及鉴定。在获得大量微生物资源的同时,也发现油樟叶内生细菌数量随季节变化,其值在0.7×103~4.5×104CFU/g,该结果与多数植物叶片组织内生细菌含量相当[3]。油樟叶内生菌的数量秋季最高,春季次之,冬季则最低,究其原因,可能与油樟叶在当季独特的生理特征有关[28];一些研究指出,随植物生长发育阶段变化的物质代谢会影响植物内生细菌群落数量和结构特征[4],特别是在旺盛生长阶段,大量而又丰富的糖、蛋白等物质为不同内生菌的生长发育提供了足够底物[29]。本研究中,春季是油樟生长量最大的季节,而秋季则是油樟叶含油量最高阶段,此两阶段是油樟叶器官中物质代谢最为旺盛和活跃阶段,丰富的底物是其内生菌数量和种类最多的原因;而在夏季和冬季内生菌数量和种类极少,可能与本区域夏季高温干旱和冬季低温寡日照等因素导致油樟生长减缓有关。进一步的,本研究中油樟中可分离培养内生菌数量较多的季节与其生长最旺和功能物质产量最高阶段显著相关,又因为内生菌与宿主植物生长和有效成分合成有关,进而可以通过对内生菌的筛选而得出有效成分含量较高的植物生长阶段,从而提高植物的利用率。
分离物数量多少会直接影响对多样性的研究,而植物内生菌群落组成极其复杂,采用传统平板法分离的微生物只能占到总数的0.1%到10.0%,不能充分反映微生物群落结构[30]。消毒方法、材料处理方法、培养基等是影响内生菌分离的重要因素[31]。本研究在分离时尽管采用了LB、TSA、阿须贝氏培养基和卵磷脂吐温80营养琼脂等前期筛选的培养基,力求最大限度地分离油樟叶中的内生细菌,但这些培养基与内生菌自然生长条件仍有较大差别,进而造成某些微生物的富集生长,而另一些微生物缺失,导致部分内生细菌资源遗漏。此外,我们前期研究中发现,由于油樟叶中大量的黏液导致接种后几乎不能分离出内生细菌,本研究中一个重大的改进是采用中和剂处理研磨叶片,该方法能较好地中和油樟叶中的黏液,促进内生菌的释放;因此为分离获得油樟叶中更多的内生细菌资源,更全面地获取油樟内生细菌的信息,后期还应进一步优化分离、培养技术。
原核生物的16S rDNA序列含有保守序列和可变序列,保守序列反应物种的亲缘关系为系统发育提供线索;可变序列是每种微生物的特征序列,是鉴定种属的分子基础,在细菌的种属分类鉴定中广泛应用[32],本研究中,对从不同季节油樟叶中根据形态特征挑取的166株内生细菌进行16S rDNA鉴定后共获得40个OTU,归属于4个门13个属,主要门类为变形菌门、厚壁菌门、放线菌门和拟杆菌门,各样品的菌群组成和丰度虽有差异,但厚壁菌门为优势菌群,约占 26.1%~100%。在不同季节,存在不同种类的内生菌主要包括:不动杆菌属、假单胞菌属、鞘氨醇单胞菌属、葡萄球菌属、芽孢杆菌属、气球菌属、赖氨酸芽孢杆菌属、考克氏菌属、罗氏菌属、微杆菌属、微球菌属、枝芽孢菌属和金黄杆菌属,其中4个季节中都有芽孢杆菌属,且表现出绝对优势,这与前人在其他植物上的研究结果相类似[33],表明该类菌可能在油樟与环境的互作过程中发挥着重要的作用。有文献报道,芽孢杆菌属中的解淀粉芽孢杆菌(Bacillusamyloliquefaciensstrain)、枯草芽孢杆菌(Bacillussubtilis)等能够产生和分泌不同类型的抗生素、生长素等,可广泛用于病虫害的生物防治,亦可改善土壤环境,提高农作物产量[34],其他类型的芽孢杆菌还具有产酶、分解纤维素等功能而被应用在饲料生产[35]、代谢工程和合成生物学等领域[36]。本研究中分离到最多的芽孢杆菌也为解淀粉芽孢杆菌和枯草芽孢杆菌,然而这些芽胞杆菌种类并不相同,由于芽胞杆菌具有高度相似的16S rDNA序列区域,后期可通过gyrA、polC和rpoB等基因来进一步鉴定芽胞杆菌属的细菌[37]。此外本研究还分离到的不动杆菌属、假单胞菌属、鞘氨醇单孢菌属、微杆菌属,该类菌已被其他研究人员用于医用抗生素筛选[38]、功能活性物质合成[39]、农业病害防治[40]、环境保护等领域[41],这些研究结果意味着,本研究分离的菌株中也极有可能存在大量未被开发利用的功能菌株,后期还需进一步对分离内生菌进行功能筛选。
植物内生菌生物多样性是衡量其内环境是否稳定和健康的一个重要指标,适宜的植物内环境,其内生细菌多样性丰富,优势度低,细菌种类分布均匀[42]。大部分研究认为内生菌是由土壤微生物入侵后和宿主协同进化而来[43],也可能是因为地区气候特征及周围植被的不同,造成大气中的微生物组成产生差异进而影响宿主[44],因此植物生长的生态环境不同,内生菌种群结构和分布必然会存在一定的差异,本研究中,不同季节油樟叶内生细菌种丰富度、多样性、均匀度、优势度存在显著差异,春季油樟叶中的内生菌多样性最高,优势度和均匀度相比夏季和秋季显著降低,而秋季油樟叶中的内生菌优势度较高,但种群种类丰富度和多样性均低于春季,这可能与同期树体所处不同的生理状态有关;相比较于春秋两季,冬季油樟叶中内生细菌丰富度、多样性指数、优势度和均匀度均急剧降低,这可能是由于同期气温、光照及降水等环境因素造成土壤及大气微生物种群结构的不同,进而是油樟叶内生菌种类向优势种群芽孢杆菌属集中,有群落单一化趋势。而夏季的高温、强光照和干旱等环境因素,使得油樟叶内生细菌种类也向优势种群芽孢杆菌属集中,虽然丰富度急剧降低,但多样性、优势度仍保持较高,均匀度则显著升高,这可能是内生菌对植物适应外界环境导致的内部微环境变化的一种响应,但相关机制还应做更进一步研究。
4 结论
本研究采用传统分离培养方法首次对宜宾地区天然香料植物油樟叶生细菌群落多样性进行研究,结果发现不同季节油樟叶片中的内生细菌数量发生显著变化,其中秋季最高,冬季最低,根据形态特征最终从4个季节的油樟叶中挑取到166株内生细菌,16S rDNA基因序列鉴定所得菌株可分为3个门13个属40个种,其中春季和秋季的内生细菌种类和数量最多,而冬季和夏季群落较为单一,数量较少,但4个季节油樟叶中优势内生细菌均为芽胞杆菌属(Bacillus)细菌,多样性指数分析表明油樟叶中内生细菌具有较丰富的遗传多样性,不同季节的种群数量、均匀度、优势度存在差异,秋季种类较丰富,优势属突出,且分布均匀,而冬季多样性、优势度和均匀度较低。本研究为进一步挖掘植物与共生内生细菌之间的互作关系提供参考依据,可为深入研究油樟叶内生微生物区系结构与其物质代谢和樟油合成调控提供供试菌种资源基础,进而对推进本区域油樟科学研究及樟油产业发展具有积极的理论与实践价值。
