小麦小穗不同粒位粒重形成的生理特性差异
2019-10-10李艳霞杨卫兵尹燕枰郑孟静杨东清骆永丽庞党伟王振林
李艳霞 杨卫兵,2 尹燕枰 郑孟静 陈 金 杨东清 骆永丽 庞党伟 李 勇,* 王振林,*
小麦小穗不同粒位粒重形成的生理特性差异
李艳霞1杨卫兵1,2尹燕枰1郑孟静1陈 金1杨东清1骆永丽1庞党伟1李 勇1,*王振林1,*
1山东农业大学作物生物学国家重点实验室, 山东泰安 271018;2北京市农林科学院/ 北京杂交小麦工程技术研究中心, 北京 100097
为探明小麦小穗上不同粒位籽粒粒重形成的生理机制, 明确限制小穗上位弱势粒充实的主要原因, 本试验选用大穗型小麦品种泰农18 (TN18)和多穗型小麦品种山农20 (SN20)为材料, 调查检测了灌浆过程中小穗上不同粒位籽粒内源激素、可溶性糖、全氮含量的动态变化以及籽粒与籽粒柄连接处横面的组织结构与不同粒位籽粒粒重的关系。花后籽粒灌浆过程中灌浆速率与籽粒内GA和IAA含量呈极显著或显著相关, 小穗基部籽粒中较高的GA和IAA含量可使蔗糖向淀粉转化开始早, 籽粒分化快, 灌浆速率高, 是小穗基部籽粒粒重高的生理机制; 扫描电镜图显示小麦籽粒灌浆初期小穗基部籽粒柄维管束横面面积明显大于上位籽粒, 微观空隙小且排列较整齐, 有利于同化物和生理活性物质的运输, 是小穗基部籽粒粒重增长快、灌浆速率高的解剖学基础;
粒位; 粒重; 内源激素; 可溶性糖; 全氮含量
小麦小穗上不同粒位籽粒发育的不均衡性限制着小麦产量的提高[1-2]。生长在小穗基部维管束结上开花较早的小花形成的籽粒灌浆速率大、充实好, 为强势粒, 生长在小穗维管组织分支上的上位小花开花迟, 灌浆速率小、充实差, 形成的籽粒发育为弱势粒[3]。梁太波等[4]以排水法研究表明山农12小麦品种强势粒的最大体积比弱势粒高出21.07%。
稻麦籽粒发育及灌浆进程和最终粒重与籽粒内源激素含量有密切关系[5-7]。内源激素可以通过影响同化物在强、弱势粒间的分配来控制灌浆启动及灌浆速率。王瑞英等[8]研究表明, 内源激素含量的不同是导致小麦强势粒与弱势粒之间粒重差异的原因之一。杨建昌等[9]也认为水稻籽粒灌浆速率与其ZR、ABA和IAA含量存在显著相关性。杨卫兵等[10]研究表明喷施ABA可显著提高灌浆前期强势粒的灌浆速率和粒重, 但对弱势粒粒重的影响不明显。植物内源激素的含量以及激素间的协同或拮抗作用调节着蔗糖、氨基酸、蛋白质及多种酶类的合成和运输速度, 进而调节籽粒灌浆进程[11-12]。
张晓融和王世之[13]研究表明稻麦不同部位籽粒的粒重和内含物与其花序上颖花不同的分化、发育及穗部维管束系统有关。也有研究表明弱势粒灌浆弱是由于蔗糖到淀粉转化过程中相关酶活性低或者调控相关酶活性基因的表达低[14]; 还有研究表明碳水化合物供应不足或库容的限制[15], 植物激素间的不平衡[5]、以及粒位间物质流的多少[16]制约着小穗上位弱势粒的发育。
虽然前人已经做了大量关于稻麦强弱势粒灌浆速率、粒重差异及激素调控强弱势粒粒重的研究, 但是前人多将小穗基部第1、第2位籽粒定义为强势粒, 第3位以上籽粒定义为弱势粒, 对不同穗型小麦小穗上不同粒位籽粒粒重差异形成的生理特性的研究较少, 且更少系统研究花后灌浆初期不同粒位籽粒柄维管束组织结构与籽粒灌浆过程中激素、同化物含量变化。本试验选用不同穗型小麦系统研究花后小穗上不同粒位籽粒灌浆过程中激素、碳氮化合物含量变化与籽粒灌浆及粒重间的关系, 旨在探讨限制小麦小穗上位籽粒粒重的主要生理机制。
1 材料与方法
1.1 试验设计
试验于2013年10月至2015年6月在山东省泰安市山东农业大学农学试验站进行。该地区位于山东中部, 地理坐标为36°09’N, 117°09’E, 平均海拔130 m, 属于温带大陆性半湿润季风气候区。年平均气温为12.9℃, 1月份最低气温–2.6℃, 7月份平均气温26.4℃, 无霜期平均为195 d。气候适合一年两熟种植制度。试验田土壤为棕壤土, 2013—2014年试验地0~20 cm 耕层土壤含有机质17.30 g kg–1、全氮1.20 g kg–1、速效磷17.0 mg kg–1和速效钾113.0 mg kg–1。2014—2015年冬小麦播种前试验地0~20 cm耕层含有机质18.11 g kg–1、全氮1.23 g kg–1、速效磷17.05 mg kg–1和速效钾112.8 mg kg–1。两年间的土壤肥力基本一致。
试验品种为多穗型小麦品种山农20 (SN20)和大穗型小麦品种泰农18 (TN18)。播种密度210万株hm–2。试验地施磷肥折合纯磷(以P2O5计算) 75 kg hm–2, 钾肥折合纯钾(以K2O计) 120 kg hm–1。磷钾肥全部底施。氮肥(全氮含量) 240 kg hm–2, 50%底施, 50%拨节期开沟追施。试验小区面积为3 m × 3 m = 9 m2, 小区内设10行, 行距0.25 m。两年试验品种、施肥量、播种密度及小区面积都相同, 播期分别为2013年10月5日和2014年10月8日。小麦生育期人工去除杂草, 其他管理同一般高产田。
1.2 测定项目及方法
开花期选择同日开花生长一致的穗挂牌标记, 花后每5~7 d取40个穗, 其中10个穗用液氮速冻后保存在–40℃冰箱, 用于激素含量的测定, 30个穗放入纸袋做好标记放入烘箱烘至恒重。取小麦穗中部小穗(基部第5~12个小穗)按照不同粒位手工脱粒, 称重计算千粒重和灌浆速率及其他生理指标。
1.2.1 小麦籽粒增重及灌浆动态 在80℃的烘箱内将小麦穗烘至恒重, 按粒位人工分开后测不同粒位籽粒的粒重并计算灌浆速率, 采用Richards方程对小麦籽粒增重动态进行模拟[17]。

式中,为籽粒重量,为生长终止量,为开花后天数,、、为参数。
1.2.2 可溶性总糖和蔗糖含量测定 采用蒽酮法测定可溶性总糖和蔗糖含量[18], 称烘干磨碎样品0.1000 g置10 mL离心管中, 加入6~7 mL水, 置沸水浴中加热提取20 min, 取出冷却, 离心5 min (2900 ×), 转移上清液至50 mL容量瓶中, 以同样方法重复提取2次, 合并上清液于50 mL容量瓶中, 定容至刻度。分别加入0.1 mL 30% KOH, 在沸水浴中放10 min, 冷却至室温(冷水), 加入3 mL蒽酮试剂, 沸水浴中10 min, 冷却震荡混匀后于620 nm比色。以分析纯蔗糖制作标准曲线。
1.2.3 全氮含量的测定 以H2SO4-H2O2联合消煮, 用半微量凯氏定氮法测不同粒位籽粒的全氮含量。
1.2.4 籽粒柄与籽粒连接处维管束横面显微结构的观察 在花后5 d籽粒形成的初期, 用刀片取小麦穗中部小穗上不同粒位籽粒与籽粒柄连接处的横切面, 放入3.5%戊二醛溶液固定24 h后, PBS缓冲液冲洗5次(每20 min一次)。用OSO4固定3 h, 然后用梯度乙醇(45%、55%、65%、75%、85%、95%、100%、100%、100%)脱水, 每30 min一次。再用醋酸异戊脂冲洗2次(每小时1次)。CO2临界点干燥, 取出粘贴样品, 喷金镀膜后。用JEDL JSM-6610LV扫描电镜观察照相。
1.2.5 内源激素含量测定 采用高效液相色谱法(HPLC)测定籽粒中4种内源激素的含量[10]。采用流动相∶甲醇∶乙腈∶0.6%乙酸(50∶45∶5, v/v), 所用分析柱为Symmetry C18 (150.0 mm × 4.6 mm, 5 μm), 柱温为25℃, 流动相流速0.6 mL min–1, 检测波长为254 nm。玉米素、赤霉素、生长素、脱落酸标样均购自美国Sigma公司。
1.2.6 数据统计分析 采用Microsoft Excel 2003和DPS7.05 (Data Processing System, V7.05)软件分析系统统计分析有关数据, 用SigmaPlot 10.0处理数据及作图。
2 结果与分析
2.1 籽粒增重动态及灌浆速率变化
由图1可以看出小穗不同粒位籽粒粒重及灌浆速率粒位间差异较大, SN20小穗基部第1、第2位籽粒灌浆速率明显高于第3位籽粒, 以最大灌浆速率出现时(花后15~20 d)差异较大。TN18小穗第1、第2位籽粒在灌浆的前期和中期籽粒灌浆速率高于第3位籽粒, 但是在灌浆后期(20 d以后), 随着第1、第2位籽粒的充实度越来越饱满, 其灌浆速率在灌浆后期下降较第3和第4、第5位快, 出现第3位籽粒的灌浆速率开始接近并高于第1、第2位籽粒灌浆速率; 小穗上第4、第5位籽粒的最大灌浆速率和粒重都明显低于小穗基部第1、第2和第3位籽粒, 仅在花后30~35 d随着小穗基部籽粒灌浆速率的快速下降, 此时第4、第5位籽粒的灌浆速率降低较缓慢, 造成其灌浆速率超过第3位和第1、第2籽粒的灌浆速率, 但是此时的灌浆速率很低, 后期植株叶片衰老, 光合产物较少, 源供应不足导致微弱的灌浆速率不足难以弥补其灌浆前中期与第1、第2位籽粒粒重较大的差异。

图1 2014–2015年花后小穗上不同粒位籽粒粒重和灌浆速率
1 2: 小穗基部第1位和第2位籽粒; 3: 小穗基部第3位籽粒; 4 5: 小穗基部第4位和第5位籽粒。
1 2: Grains from the first and second kernel positions in the base of the spikelet; 3: Grains from the third kernel position in the base of the spikelet; 4 5: Grains from the fourth and fifth kernel positions in the base of the spikelet.
2.2 成熟期不同粒位间粒重的差异
由图2可以看出不同穗型小麦品种小穗上不同粒位籽粒粒重差异较大。大穗型小麦品种TN18小穗上第3位籽粒与第1、第2位籽粒粒重差异较小, 第4、第5位籽粒与第3位籽粒粒重差异较大; 多穗型小麦品种SN20小穗上第3位籽粒粒重明显小于第1、第2位籽粒。品种间SN20小穗第1、第2位籽粒粒重大于TN18小穗基部第1、第2位籽粒粒重。由此可以得出多穗型品种SN20小穗基部第1、第2位籽粒较上位籽粒有较大的优势; 成熟期大穗型小麦品种TN18小穗基部第3位与第1、第2位籽粒间的粒重差异明显小于SN20小穗第3位籽粒与第1、第2位籽粒粒重间的差异。
2.3 花后不同粒位籽粒可溶性总糖和蔗糖含量的变化
花后籽粒灌浆初期籽粒可溶性总糖和蔗糖含量最高, 之后其含量不断下降且灌浆前期籽粒可溶性总糖和蔗糖含量下降速率快。灌浆前期TN18小穗上位(第3位和第4、第5位)籽粒可溶性总糖和蔗糖含量都高于基部籽粒(第1、第2位)籽粒(图3)。随着灌浆进程, 小穗上位籽粒(第3位和第4、第5位)在灌浆中后期可溶性总糖和蔗糖含量下降速率较基部第1、第2位籽粒快。
2.4 籽粒灌浆期不同粒位籽粒全氮含量的变化
由图4可以看出灌浆过程中籽粒全氮含量先降低后升高的变化趋势。灌浆早期SN20全氮含量明显高于TN18, 成熟期籽粒全氮含量反而低于TN18; 整个灌浆过程中SN20全氮含量的变化幅度较TN18大; 粒位间, 除TN18灌浆早期第4、第5位籽粒高于第1、第2位籽粒外, 灌浆过程中小穗基部第1、第2位籽粒全氮含量高于小穗基部第3和第4、第5位籽粒, 且大穗型小麦品种TN18小穗第4、第5位籽粒的全氮含量的最低点较小穗基部第1、第2和第3位籽粒出现的晚, 这种不同粒位籽粒全氮含量的变化进一步证明了小穗上位弱势籽粒灌浆启动晚的发育特性。
2.5 灌浆期不同粒位籽粒内源激素含量的变化
籽粒灌浆过程中GA呈先升高后降低的单峰变化趋势, 其峰值出现在花后20 d左右。籽粒灌浆过程中SN20籽粒GA含量上升快, 花后15 d时含量已经明显高于TN18, 且灌浆中期保持较高含量, 花后25 d以后开始下降; 与SN20相比, TN18籽粒中GA上升慢, 达到峰值(花后20 d)后就马上下降。不同粒位间SN20小穗第1、第2位籽粒GA含量高于第3位籽粒; TN18小穗基部第1、第2位籽粒GA含量明显高于第4、第5位籽粒, 第3位籽粒灌浆前期和中期(花后20 d以前)低于第1、第2位籽粒但高于第4、第5位籽粒, 灌浆后期第3位籽粒GA含量下降慢, 其含量高于第1、第2和第4、第5位籽粒。ZR含量在10 d左右有一小峰值后下降, 直到花后20~25 d达到低谷接着快速上升到最高点后又开始下降(图5)。花后IAA含量呈单峰变化曲线, 峰值出现在花后20 d; 第1、第2位籽粒IAA含量高于第3位, 仅TN18小穗上第3位籽粒IAA含量在花后15 d前后高于第1、第2位籽粒。籽粒ABA含量在灌浆过程中含量波动较大, 呈双峰变化趋势, 峰值分别出现在花后10 d和花后20~25 d, 花后15 d籽粒ABA含量处于低谷; 灌浆前期SN20小穗第3位籽粒ABA含量高于小穗第1、第2位籽粒, TN18小穗第4、第5位籽粒ABA含量高于第1、第2和第3位籽粒, 小穗上位籽粒的ABA含量高于基部籽粒。
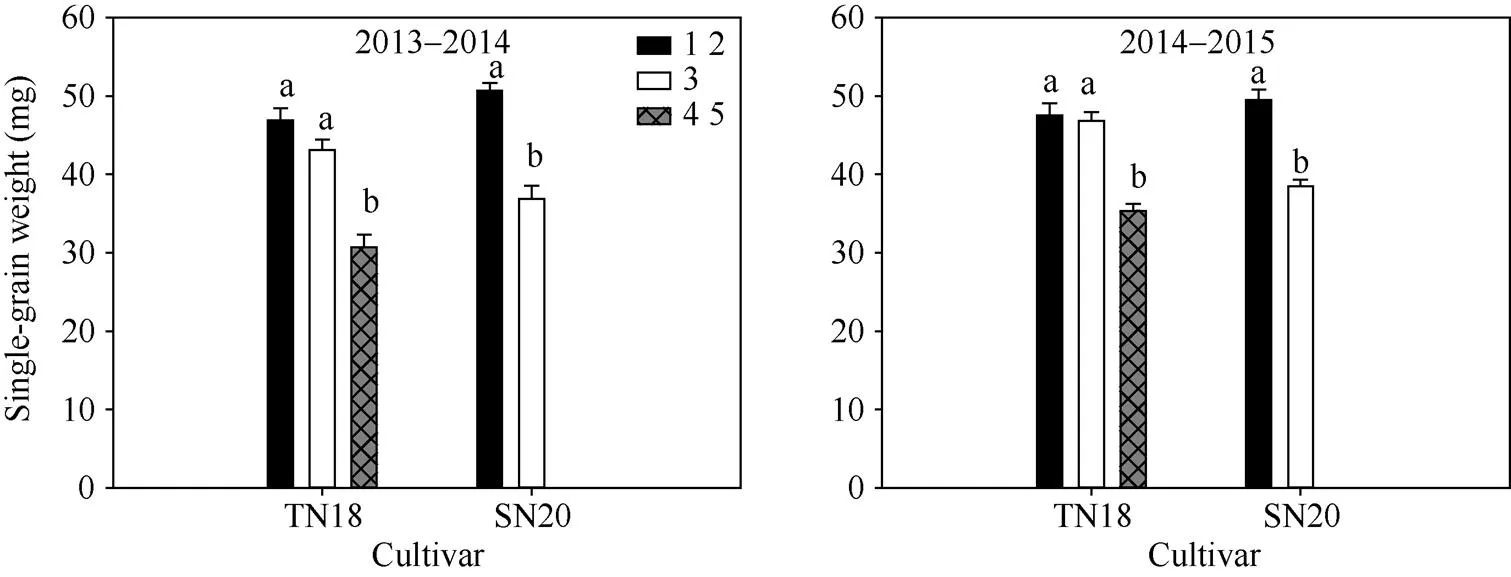
图2 2013–2014和2014–2015年成熟期小麦小穗上不同粒位籽粒单粒重
1 2: 小穗基部第1位和第2位籽粒; 3: 小穗基部第3位籽粒; 4 5: 小穗基部第4位和第5位籽粒。
1 2: grains from the first and second kernel positions in the base of the spikelet; 3: grains from the third kernel position in the base of the spikelet; 4 5: grains from the fourth and fifth kernel positions in the base of the spikelet.
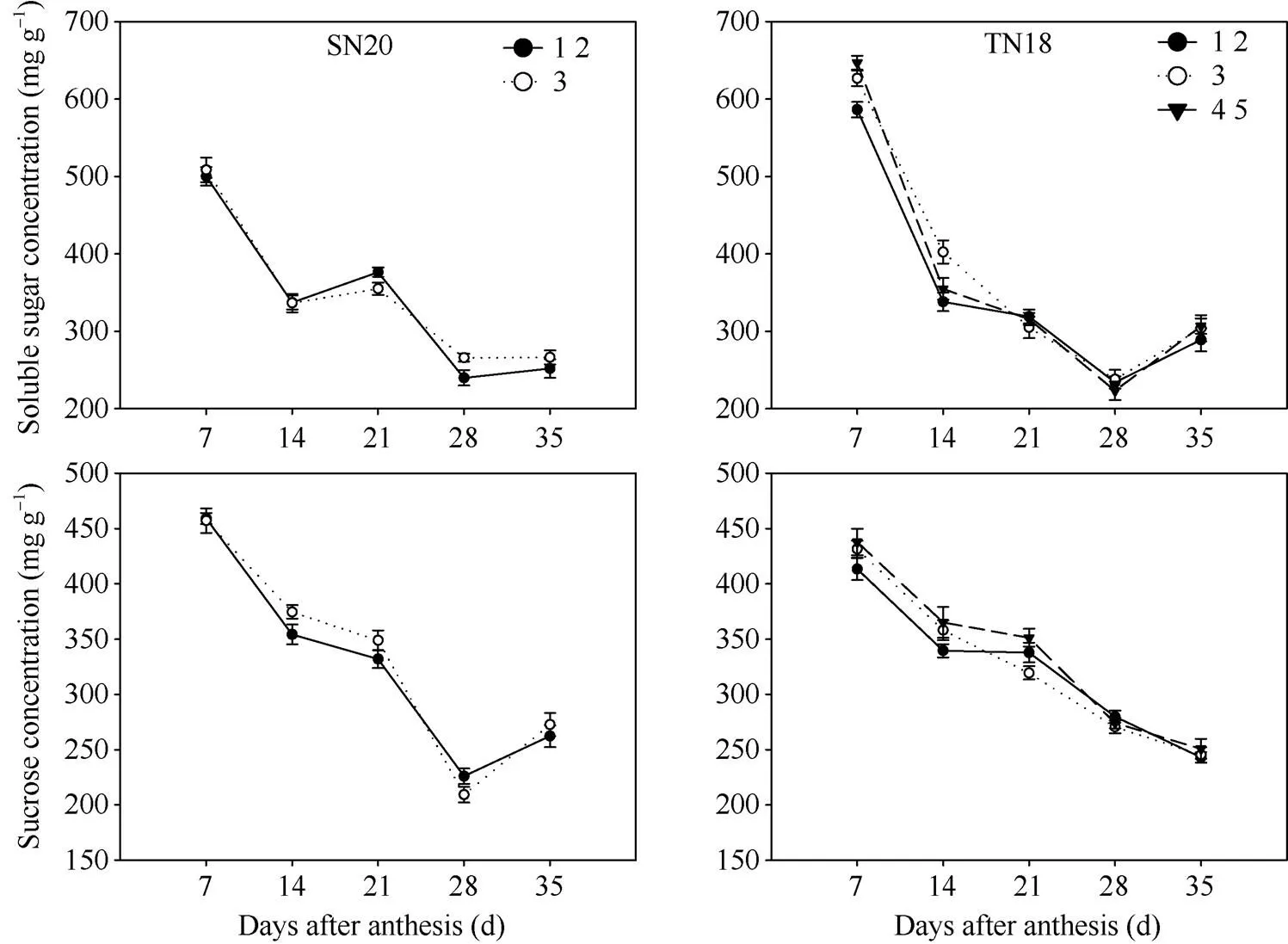
图3 2013–2014不同穗型小麦小穗上不同粒位籽粒灌浆期可溶性总糖和可溶性蔗糖含量变化
1 2: 小穗基部第1位和第2位籽粒; 3: 小穗基部第3位籽粒; 4 5: 小穗基部第4位和第5位籽粒。
1 2: grains from the first and second kernel positions in the base of the spikelet; 3: grains from the third kernel position in the base of the spikelet; 4 5: grains from the fourth and fifth kernel positions in the base of the spikelet.

图4 2013−2014年花后小穗上不同粒位籽粒全氮含量的变化
1 2: 小穗基部第1位和第2位籽粒; 3: 小穗基部第3位籽粒; 4 5: 小穗基部第4位和第5位籽粒。
1 2: grains from the first and second kernel positions in the base of the spikelet; 3: grains from the third kernel position in the base of the spikelet; 4 5: grains from the fourth and fifth kernel positions in the base of the spikelet.
2.6 籽粒灌浆速率与籽粒可溶性总糖、蔗糖及内源激素含量的相关性
两品种小麦籽粒灌浆速率与籽粒内可溶性总糖的相关性不显著, 与蔗糖的相关性品种间有差异, SN20籽粒灌浆速率与蔗糖含量相关性不显著, TN18籽粒灌浆速率与籽粒内蔗糖含量呈显著相关。籽粒灌浆速率与IAA呈显著或极显著相关, TN18籽粒灌浆速率与GA含量呈极显著相关, SN20籽粒灌浆速率与GA含量相关性不显著。籽粒灌浆速率与籽粒中ZR和ABA含量两品种都表现相关性不显著。

表1 小麦籽粒灌浆过程中灌浆速率与籽粒中碳水化合物及内源激素的相关性
*,**分别表示在0.05和0.01水平相关性显著。
*,**Significant at the 0.05 and 0.01 probability levels, respectively.
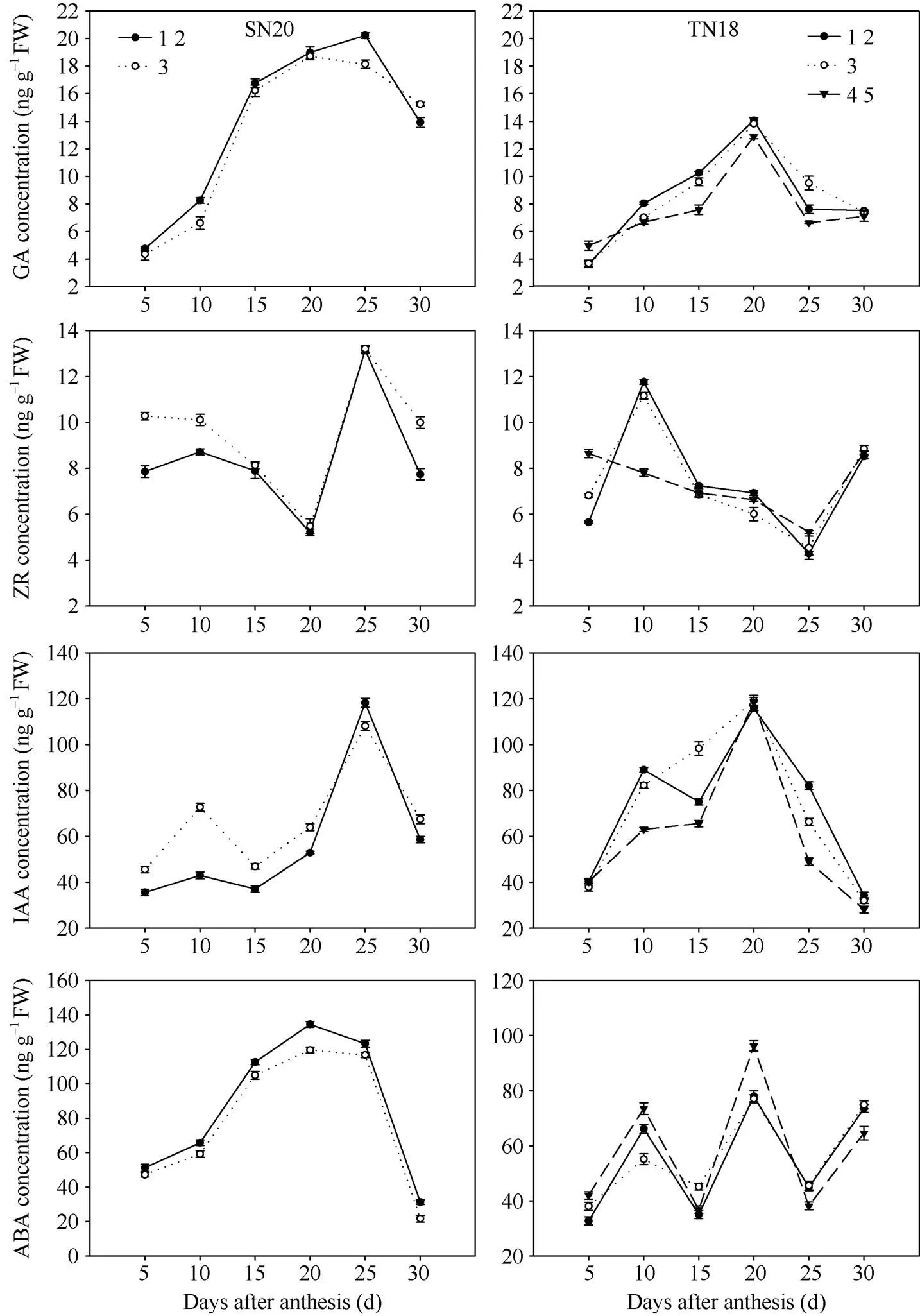
图5 2014–2015年籽粒灌浆过程中不同粒位籽粒内源激素含量的变化
1 2: 小穗基部第1位和第2位籽粒; 3: 小穗基部第3位籽粒; 4 5: 小穗基部第4位和第5位籽粒。
1 2: grains from the first and second kernel positions in the base of the spikelet; 3: grains from the third kernel position in the base of the spikelet; 4 5: grains from the fourth and fifth kernel positions in the base of the spikelet.
2.7 不同粒位籽粒柄维管束横切面的组织结构
由图6可以看出TN18和SN20从茎秆向籽粒运输同化物及生理活性物质的唯一通道即籽粒与籽粒柄连接处维管束, TN18小穗1位籽粒的籽粒柄的横面最大, 粒位间TN18小穗基部1位籽粒粒柄维管束的横面面积略大于第3位籽粒, 明显大于第4位籽粒。形状上, TN18小穗基部第1位和第3位为近圆形, 第4位籽粒柄的横切面为扁长型。多穗型小麦品种SN20小穗第1位籽粒的籽粒柄的横面面积明显大于第3位籽粒, 小穗第4位籽粒柄横面面积略小于第3位籽粒。从微观上看随着粒位的上升, 组织空隙也加大了。小穗上位籽粒与籽粒柄连接处这种维管束横面面积变小及微观空隙变大会不利于同化物及生理活性物质的运输。

图6 TN18和SN20小麦小穗上不同粒位籽粒柄与籽粒连接处横切面的扫描电镜照片
1: 小穗基部第1位籽粒; 3: 小穗基部第3位籽粒; 4: 小穗基部第4位籽粒。从左到右依次放大50×、150×、500×、1500×。
1: Grains from the first kernel position in the base of the spikelet; 3: Grains from the third kernel position in the base of the spikelet; 4: Grains from the fourth kernel position in the base of the spikelet. Magnified 50×, 150×, 500×, 1500× from left to right.
3 讨论
小麦不同粒位籽粒的粒重受遗传因素[19]、营养物质供应水平[20]、外界环境因素[21]等的影响。小麦小穗上不同粒位籽粒发育的不均衡性与进入其中维管束分布的不平衡性密切相关。同一穗上不同籽粒间存在着同化产物的竞争效应, 而小穗轴的维管束组织结构差异是影响光合产物分配、籽粒充实度和结实特征的重要因素之一[22]。前人通过对小穗的组织解剖研究认为, 小穗基部第1、第2位小花维管束直接独立地来自小穗基部维管束, 通向其籽粒的维管束数较多且面积也较大, 第3位以上的小花的维管束主要来自其下位花的分支, 维管束数较少, 面积也较小[23-24]。所以导致小穗上位小花对有机营养的强烈竞争。黄升谋等[25]研究也表明, 水稻弱势粒结实率低与其源不足, 维管束负担过重, 有机物运输不畅有关。本试验研究结果与前人研究结果相似, 小穗基部第1、第2位籽粒柄的维管束横面面积大, 且组织微观结构排列紧密而整齐, 有利于营养物质的运输。不同的是大穗型小麦品种TN18中部小穗第3朵花的维管束部分直接来源于小穗基部维管束结节, 部分来自第2朵花维管束的分枝; 而多穗型小麦品种SN20小穗第3朵小花的维管束来自第2位小花。这种维管束结构差异可能是导致多穗型小麦SN20小穗3位籽粒粒重明显低于TN18小穗第3位籽粒粒重的原因。当然, 维管束系统作为整个运输过程的一个环节, 其差异只能是导致穗器官不均衡发育的原因之一。维管束的分化需要一定的营养物质, 特别是蔗糖和吲哚乙酸[26]。幼穗分化的早期养分分配的差异可能是导致维管束分化差异的原因之一, 这种差异反过来又促进穗器官发育的不均衡, 最终导致小穗上不同粒位间籽粒在物质转运中的差异。在小麦栽培管理中, 我们可以通过改进栽培措施, 培育壮苗, 改善植株的营养条件, 增加穗器官的养分供应, 提高籽粒的饱满度, 增加产量。
小麦的同一穗上籽粒大小不一的现象, 并不仅仅是由于养料供应的不足, 无论养料充足与否, 均会出现籽粒大小的差异, 这与籽粒在小穗上的位置有关。籽粒在小穗部的位置决定了籽粒结实与物质积累的空间分布特征。李明慧等[27]研究发现, 在同一小穗中, 基部第1、第2粒中可溶性糖含量较高, 第3粒中含量较低, 第4粒中含量极低; 本研究结果与其不同, 灌浆前期小穗上位(第3位和第4、第5位)籽粒可溶性总糖和蔗糖含量都高于基部籽粒(第1、第2位), 进一步证明了小麦小穗上位小花发育迟, 籽粒结实和灌浆启动晚。小麦籽粒灌浆前期籽粒中可溶性糖和蔗糖含量高, 灌浆过程中籽粒中蔗糖不断地向淀粉快速转化来促进籽粒胚乳的快速发育, 但是随着小麦籽粒胚乳中淀粉的不断合成积累, 糖含量不断“被稀释”, 其浓度也快速降低。
杨建昌等[9]研究表明, 弱势粒的灌浆启动迟、灌浆速率和生长终值均小于强势粒。郭文善等[24]研究认为, 小麦不同粒位籽粒的灌浆历期相近, 小穗上第3粒位籽粒的灌浆速率显著低于第1、第2位籽粒。有人认为延长灌浆时间是籽粒增重的有效因素[28]。Sayed等[29]认为粒重受灌浆持续期的影响不大。顾自奋等[30]认为灌浆强度与灌浆时间均组成对籽粒增重的同一型多元回归关系, 简单排斥二者中任一因素的作用是不恰当的。本试验结果认为粒重由灌浆速率和灌浆时间共同决定。小麦籽粒灌浆过程中, 营养物质遵循“就近供应”的原则来促进基部第1、第2位籽粒的发育来构建小麦胚体的形成和淀粉等储藏物质的积累, 但是当其积累到一定的程度时小麦籽粒会出现“饱和”状态, 此时就会通过一系列信号传导来迫使营养物质向上位籽粒运输来促进上位籽粒的生长。延长灌浆后期籽粒的有效灌浆时间可以提高小穗上位籽粒的粒重, 这在大穗型小麦品种中表现的更突出。
籽粒灌浆速率和最终粒重, 在很大程度上还受籽粒内激素的平衡和调节。强势粒高峰期GA含量高于弱势粒[9]。本试验结果表明小穗基部籽粒GA含量高于上位籽粒; 多穗型小麦品种SN20灌浆期籽粒GA含量高于大穗型小麦品种TN18; 田淑兰等[31]研究表明, 籽粒灌浆期间不同粒位间籽粒灌浆速率的变化趋势与其内源IAA含量的变化相似, 且在灌浆前期, 外施IAA可代替强势粒对弱势粒的这种抑制作用, 推测IAA通过调节有机养料的分配控制籽粒的发育。刘仲齐等[32]报道, IAA含量与小麦籽粒淀粉积累速率及蔗糖转化酶、蔗糖合成酶的活性呈极显著正相关。本研究认为: 小穗基部位籽粒IAA含量和GA含量高于上位籽粒, 影响养料的调配从而控制籽粒的生长发育, 使强势粒受精后蔗糖向淀粉的转化速率快, 灌浆启动早, 灌浆速率高。ABA处理麦穗阻止了小穗上位小花的结实, 但对小穗基部第1、第2小花影响不大, 表明小穗上位小花对ABA含量变化反应较敏感,这可能是由于ABA含量增加阻碍了通到上位小花的维管束的形成[33]。也有研究认为外源ABA可以提高库细胞对同化物的主动吸收能力[34]。还有研究发现, 小麦籽粒灌浆前期, 内源ABA的含量对籽粒灌浆速率的影响前期表现为正效应, 后期则为负效应。强势粒ABA含量的峰值先于弱势粒出现, 并且籽粒充实度好的组合强势粒中ABA含量高于充实度差的组合[9,35]。本研究结果与前人的研究结果不同, 灌浆过程中上位籽粒ABA含量高于基部强势粒, 且灌浆过程中ABA含量变化较大。认为籽粒ABA的含量主要受环境影响较大。所以ABA的生理功能有待进一步的研究。
4 结论
粒重由品种特性及其在小穗上的位置决定, 并受栽培措施的调控。花后籽粒灌浆过程中小穗基部籽粒中较高的GA和IAA含量可使籽粒蔗糖向淀粉转化开始早, 籽粒分化快, 灌浆速率高, 是小穗基部籽粒粒重高的生理机制。小麦籽粒灌浆初期小穗基部籽粒柄维管束的横面面积明显大, 组织微观空隙小且排列较规则, 有利于同化物和生理活性物质的运输, 是小穗基部籽粒灌浆初期籽粒粒重增长快、灌浆速率高的解剖学基础。
[1] Fu J, Huang Z H, Wang Z Q, Yang J C, Zhang J H. Pre-anthesis nonstructural carbohydrate reserve in the stem enhances the sink strength of inferior spikelets during grain filling of rice., 2011, 123: 170–182.
[2] Jiang D, Cao W, Dai T, Jing Q. Activities of key enzymes for starch synthesis in relation to growth of superior and inferior grains on winter wheat (L.) spike., 2003, 41: 247–257.
[3] Mohapatra P K, Patel R, Sahu S K. Time of flowering affects grain quality and spikelet partitioning within the rice panicle., 1993, 20: 231–242.
[4] 梁太波, 尹燕枰, 蔡瑞国, 闫素辉, 李文阳, 耿庆辉, 王平, 王振林. 大穗型小麦品种强、弱势籽粒淀粉积累和相关酶活性的比较. 作物学报, 2008, 34: 150–156. Liang T B, Yin Y P, Cai R G, Yan S H, Li W Y, Geng Q H, Wang P, Wang Z L. Starch accumulation and related enzyme activities in superior and inferior grains of large spike wheat., 2008, 34: 150–156 (in Chinese with English abstract).
[5] Yang J. C. Abscises acid and ethylene interact in wheat grains in response to soil drying during grain filling., 2006, 171: 293–303.
[6] Zhu J X, Jang D, Cao W X, Dong T B. Relationships of endogenous plant hormones to accumulation of grain protein and starch in winter wheat under different post-anthesis soil water statuses., 2003, 41: 117–127.
[7] Liu Y E, Liu P D, Zhang S T, Wang J. Hormonal changes caused by the Xenia effect during grain filling of normal corn and high-oil corn crosses., 2010, 50: 215–221.
[8] 王瑞英, 于振文, 潘庆民, 许玉敏. 小麦籽粒发育过程中激素含量变化. 作物学报, 1999, 25: 227–232. Wang R Y, Yu Z W, Pan Q M, Xu Y M. Changes of endogenous plant hormone contents during grain development in wheat., 1999, 25: 227–232 (in Chinese with English abstract).
[9] 杨建昌, 刘立军, 王志琴, 郎有忠, 朱庆森. 稻穗颖花开花时间对胚乳发育的影响及其生理机制. 中国农业科学, 1999, 32: 44–51. Yang J C, Liu L J, Wang Z Q, Lang Y Z, Zhu Q S. Effects of flowering time of spikelets on endosperm development in rice and its physiological mechanism., 1999, 32: 44–51 (in Chinese with English abstract)
[10] 杨卫兵, 王振林, 尹燕枰, 李文阳, 李勇, 陈晓光, 王平, 陈二影, 郭俊祥, 蔡铁, 倪英丽. 外源ABA和GA对小麦籽粒内源激素含量及其灌浆进程的影响. 中国农业科学, 2011, 44: 2673–2682. Yang W B, Wang Z L, Yin Y P, Li W Y, Li Y, Chen X G, Wang P, Chen E Y, Guo J X, Cai T, Ni Y L. Effects of spraying exogenous ABA or GA on the endogenous hormones concentration and filling of wheat grains., 2011, 44: 2673–2682 (in Chinese with English abstract).
[11] 戴忠民. 喷施6BA和ABA对小麦籽粒胚乳细胞增殖及淀粉积累动态的影响. 麦类作物学报, 2008, 28: 484–489. Dai Z M. Effects of 6BA and ABA on endosperm cell propagation and starch accumulation in grains of winter wheat., 2008, 28: 484–489 (in Chinese with English abstract).
[12] Travaglia C, Cohen A C, Reinoso H, Castillo C, Bottini R. Exogenous abscisic acid increases carbohydrate accumulation and redistribution to the grains in wheat grown under field conditions of soil water restriction., 2007, 26: 285–289.
[13] 张晓融, 王世之. 小麦穗、小穗及籽粒差异的解剖结构及生理原因的研究. 作物学报, 1993, 19: 103–109. Zhang X R, Wang S Z. Studies on the anatomical and physiological factors influencing the unbalanced development of the spikes and grains of wheat., 1993, 19: 103–109 (in Chinese with English abstract).
[14] Wang E T, Wang J N, Zhu X D, Hao W, Wang L Y, Li Q, Zhang L X, He W, Lu B R, Lin H X, Ma H, Zhang G Q, He Z H. Control of rice grain-flling and yield by a gene with a potential signature of domestication., 2008, 40: 1370–1374.
[15] Murty P, Murty K. Spikelet sterility in relation to nitrogen and carbohydrate contents in rice., 1982, 25: 40–48.
[16] 姜东, 于振文, 李永庚, 余松烈. 高产小麦强势和弱势籽粒淀粉合成相关酶活性的变化. 中国农业科学, 2002, 35: 378–383. Jiang D, Yu Z W, Li Y K, Yu S L. Dynamic changes of enzyme activities involving in starch synthesis in superior and inferior grains of high-yield winter wheat., 2002, 35: 378–383 (in Chinese with English abstract).
[17] Richards F J. A flexible growth function for empirical use., 1959, 10: 290–301.
[18] Fales F. The assimilation and degradation of carbohydrates by yeast cells., 1959, 10: 290–301.
[19] Daniel J M, Gustavo A S. Individual grain weight responses to genetic reduction in culm length in wheat as affected by source-sink manipulations., 1995, 43: 55–62.
[20] Walley F, Pennock D, Solohub M. Hnatowich. Spring wheat () yield and grain protein responses to N fertilizer in topographically defined landscape positions., 2001, 81: 505–514.
[21] Chen Y, Yuan L P, Wang X H, Zhang D Y, Chen J, Deng Q Y, Zhao B R, Xu D Q. Relationship between grain yield and leaf photosynthetic rate in super hybird rice., 2007, 33: 235–243.
[22] Mohapatra P K, Patel R, Sahu S K. Time of flowering affects grain quality and spikelet partitioning within the rice panicle., 1993, 20: 231–242.
[23] Whingwiri E E, Kuo J, Stern W R. The vascular system in the rachis of a wheat ear, 1981, 48: 189–202.
[24] 郭文善, 彭永欣, 封超年, 严六零, 高年春. 小麦不同花位籽粒重差异及其原因分析. 江苏农学院学报, 1992, 13(3): 1–7. Guo W S, Peng Y X, Feng C N, Yan L L, Gao N C. Weight differences of grains at different positions and Its reason., 1992, 13(3): 1–7.
[25] 黄升谋, 邹应斌. 亚种间杂交水稻5460S/广抗粳2号充实度的研究. 中国农学通报, 2002, 18(5): 11–15.Huang S M, Zou Y B. Study on grain-filling rate of intersubspecific hybird combination 5460S/Guangkanggeng 2., 2002, 18(5): 11–15 (in Chinese with English abstract).
[26] Shininger T L. The control of vascular development., 1979, 30: 313–332.
[27] 李明慧, 张庆琛, 李刚, 沈光辉, 徐士库, 刘亚平, 韩战敏. 小麦穗与小穗及籽粒差异的解剖结构和生理原因研究. 安徽农业科学, 2008, 36: 938–940. Li M H, Zhang Q S, Li G, Shen G H, Xu S K, Liu Y P, Han Z M. Research on the relationship between the development imbalance of wheat ear, Spikelet and grain and vascular bundle system and Its physiological reason., 2008, 36: 938–940 (in Chinese with English abstract).
[28] Pinthus M J, Sar-shalom Y. Day matter accumulation in the grains of wheat (L.) cultivars differing in grains weight., 1978, 42: 469–471.
[29] Sayed H I, Gadallah A M. Variation in day matter and grain filling characteristics in wheat cultivars., 1983, 7: 61–71.
[30] 顾自奋, 李金诚, 翁训珠. 灌浆强度和时间对大麦籽粒增重的影响. 作物学报, 1987, 13: 288–295. Gu Z F, Li J C, Weng X Z. Effect of filling intensity and duration on increasing grain-weight in barley., 1987, 13: 288–295 (in Chinese with English abstract).
[31] 田淑兰, 王熹. IAA与亚种间杂交稻籽粒发育的关系及烯效唑的调节. 中国水稻科学, 1998, 12: 99–104. Tian S L, Wang X. The relationship between IAA and grain development ofhybrid rice and regulation with S-07., 1998, 12: 99–104 (in Chinese with English abstract).
[32] 刘仲齐, 吴兆苏, 俞世蓉. 吲哚乙酸和脱落酸对小麦子粒淀粉积累的影响. 南京农业大学学报, 1992, 15(1): 7–12. Liu Z Q, Wu Z S, Yu S R. Effects of IAA and ABA on starch accumulation in wheat grain., 1992, 15(1): 7–12 (in Chinese with English abstract).
[33] Radley M. Effect of abscisic and gibberellic acid on grain set in wheat., 1980, 95: 409–414.
[34] Sehussler J R, Brenner M L, Brun W A. Abscisic acid and its relationship to seed filling in soybeans., 1984, 76: 301–306.
[35] 柏新付, 蔡永萍, 聂凡. 脱落酸与稻麦子粒灌浆的关系. 植物生理学通讯, 1989, (3): 40–41. Bai X F, Cai Y P, Nie F. Relationship between absciaic acid and grain filling of rice and wheat., 1989, (3): 40–41 (in Chinese with English abstract).
Difference of physiological characteristics of grain weight at various kernel positions in wheat spikelets
LI Yan-Xia1, YANG Wei-Bing1,2, YIN Yan-Ping1, ZHENG Meng-Jing1, CHEN Jin1, YANG Dong-Qing1, LUO Yong-Li1, PANG Dang-Wei1, LI Yong1,*, and WANG Zhen-Lin1,*
1Agronomy College, Shandong Agricultural University / State Key Laboratory of Crop Biology, Tai’an 271018, Shandong, China;2Beijing Academy of Agriculture and Forestry Sciences, Beijing Engineering Research Center for Hybrid Wheat, Beijing 100097, China
The object of the experiment was to explore the main physiological mechanism of heterogeneous development of grains located at the various kernel positions in wheat spikelets, further to reveal the factors restricting grain filling of inferior grains located on the distal kernel positions. Two varieties with different spike sizes of winter wheat (a large-spike cultivar Tainong 18 and a multiple-spike cultivar Shannong 20) were used to as the experimental materials. We have studied the dynamic changes of concentrations of endogenous hormones, soluble sugar and total nitrogen in grains located at different kernel positions in wheat spikelets during grain-filling period, and the microstructure of vascular bundle junction on kernel handle and kernel weight. Correlation analysis showed that the grain filling rate was significantly correlated with the contents of GA and IAA in grains. Higher GA and IAA contents in proximal kernels could make the conversion from sucrose to starch start earlier, the seed differentiate rapidly and the grain-filling rate higher,which is the physiological mechanism of the higher weight of proximal kernel in wheat spikelets. The micrograph showed that the cross-sectional area of vascular bundle of seed stalk on proximal kernels at the early stage of grain-filling period was significantly larger than that of distal kernels, and the microcosmic space was small and orderly, which is conducive to the transport of assimilates and physiological active substances, showing an anatomical basis for the rapid increase of seed weight and higher grain-filling rate in the proximal kernels of spikelets.
kernel position; kernel weight; endogenous hormone; soluble sugar; total nitrogen content
本研究由国家自然科学基金项目(31271661, 30871477), 国家重点基础研究发展计划(973计划)项目(2015CB150404), 北京市农林科学院青年基金(QNJJ201629), 国家重点研发计划项目(2016YFD0300400), 山东省高等学校科技计划项目(J14LF12), 山东省泰山产业领军人才项目和山东省农业重大应用技术创新课题项目资助。
The work was supported by the National Nature Science Foundation of China (31271661, 30871477), the National Basic Research Program of China (973 Program, 2015CB150404), the Youth Fund Project from Beijing Academy of Agricultural and Forestry Sciences (QNJJ201629), the National Key Research and Development Plan (2016YFD0300400), the Shandong Province Higher Educational Science and Technology Plan (J14LF12), the Shandong Province Mount Tai Industrial Talents Program, and Shandong Province Key Agricultural project for Application Technology Innovation.
王振林, E-mail: zlwang@sdau.edu.cn; 李勇, E-mail: woooowo@126.com
E-mail: liyanxia429@126.com
2019-01-10;
2019-06-22;
2019-07-15.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20190712.1624.008.html
10.3724/SP.J.1006.2019.91004
