酒酒球菌胁迫适应性机制的研究进展
2019-08-07赵红玉刘龙祥
赵红玉 李 华 刘龙祥 彭 帅 王 华
(西北农林科技大学葡萄酒学院 陕西杨凌712100)
苹果酸乳酸发酵 (Malolactic fermentation,MLF)是在酒精发酵结束后再发酵的过程,又称为“二次发酵”。发酵机理是在苹果酸乳酸酶催化下将苹果酸分解为乳酸和二氧化碳,其中,乳酸菌参与合成苹果酸乳酸酶[1-3]。MLF 被认为是酿造优质红葡萄酒及部分白葡萄酒中必不可少的环节,它的顺利进行会降低葡萄酒的酸度,提高葡萄酒的微生物稳定性,增加酒体的果香味,降低酒体的收敛性,丰富酒体的结构感,使其更加饱满、馥郁等[4-5]。目前,应用于葡萄酒使其进行MLF 的主要人为诱导方法是向酒体中加入乳酸菌直投式发酵剂[6-9]。参与MLF 的乳酸菌主要有:明串珠菌属(Leuconostoc)、片球菌属(Pediococcus)和乳杆菌属(Lactobacillus),其中,酒酒球菌是19 世纪60年代中期在16SRNA 序列分析的基础上被鉴定并重新命名的。它是使苹果酸乳酸发酵能够顺利进行的最重要的启动者,也是在葡萄酒的严峻生境中耐受性最强的菌种[10-13]。
在葡萄酒的酿造过程中,酒体中乙醇浓度、SO2含量、酒体pH 值、温度都是制约酒酒球菌生长的重要因素[14],从而对MLF 的顺利进行产生不利影响。葡萄酒微生物学家探究了酒酒球菌在葡萄酒中的适应性机制,同时,需要继续发展和完善直投式发酵剂的生产技术,以获得耐受性强、发酵活力高、活菌数高、MLF 效率高的菌株,在接种后可直接使用,无需中间继代培养和预培养的商业化产品。对酒酒球菌的胁迫适应性机制研究颇受关注[15~18]。本文主要从pH 值、乙醇浓度和温度3个方面对酒酒球菌的影响及其在葡萄酒恶劣生境中形成的适应性机制进行综述。
1 乙醇对酒酒球菌的影响及胁迫反应机制
1.1 乙醇胁迫对酒酒球菌的影响
早期有关酒酒球菌(Oenococcus oeni)抗逆性机理的研究主要在细胞膜结构和成分的变化上。酒酒球菌是革兰氏阳性细菌,没有完整的细胞结构,直接由细胞膜包被,所以细胞膜是其抵御不利环境的第一道屏障[17]。在受到胁迫反应时,为避免影响细胞新陈代谢的正常进行,细胞膜物理特性和流动性需要不断调整,以维持其屏障作用和酶活性。乙醇胁迫主要影响酒酒球菌细胞膜的结构和功能,研究显示,随着乙醇浓度的升高,细胞膜的流动性和通透性增加,质子或其它离子的透过率增加,将致使能量转化效率降低[19]。随着乙醇处理时间的延长,细胞存活率显著降低,不同乙醇浓度对细胞的影响出现明显差异。处理时间30 min 时,14%乙醇处理会使菌体细胞发生不可逆的硬化,而10%和12%乙醇处理发生的硬化是可逆的,当胁迫条件移除时细胞膜会恢复至初始状态。处理时间相同时,细胞存活率随着乙醇浓度的增加而降低,即乙醇浓度越高对细胞活力的影响越大[20]。也有研究指出,当环境中存在10%乙醇或更少量乙醇(3%~5%或7%)时,可促进菌体的生长,当酒精浓度高于12%时,将显著抑制酒酒球菌的生长[21]。
Nair 等[21]对酒酒球菌PSU-1 进行转录组学及蛋白质组学分析,发现在12%乙醇存在的条件下,与碳水化合物转移有关的基因表达会受到抑制,与细胞组件合成相关的基因和蛋白质表达量增加。此外,两个与乙酰转移酶相关的基因表达也受到显著抑制。由蛋白质组学分析可知,酒酒球菌PSU-1 在12%乙醇处理5 h 后,蛋白质含量及稳定性显著下降,如clpC、clpE 及参与胞壁质合成的蛋白质如MurC、MurD 和GlmS 等。
1.2 乙醇胁迫的适应性机制
细胞受到乙醇胁迫时,细胞膜组分会进行适当调整以适应不利环境。其中,较为明显的是环丙烷脂肪酸含量增加,环丙烷环状结构中酰基链比顺式的双键更稳定,从而增强菌体细胞对胁迫环境的耐受性[22]。同时,细胞膜中乳杆菌酸(lactobacillic acid,C19cyc11)含量增加,表示C19cyc11可能参与了胁迫反应,此机制仍需要进一步探究。乙醇可通过自由扩散进出细胞,因此环境中存在乙醇时会影响细胞膜的通透性。菌体细胞受到乙醇胁迫时,细胞膜上的不饱和脂肪酸含量增加使流动性增加,细胞膜通透性增加[23]。菌体细胞中的柠檬酸代谢可提供细胞生长存活所需的能量,当受到乙醇胁迫时,很多调控菌体细胞中柠檬酸盐代谢的基因会过量表达,这就表明柠檬酸盐代谢可能参与了胁迫反应[24]。进一步研究得到柠檬酸盐并不直接发挥其胁迫适应性作用,而是其代谢的产物,如乙酸、二乙酰发挥作用,且参与代谢的基因表达量与乙醇浓度有关[25]。其中,酒酒球菌SD-2a 在经乙醇胁迫处理后,其降解苹果酸的能力与细胞膜中C14:0 的含量之间存在明显的正相关性,即细胞膜中C14:0 可能参与了细胞的乙醇胁迫反应。
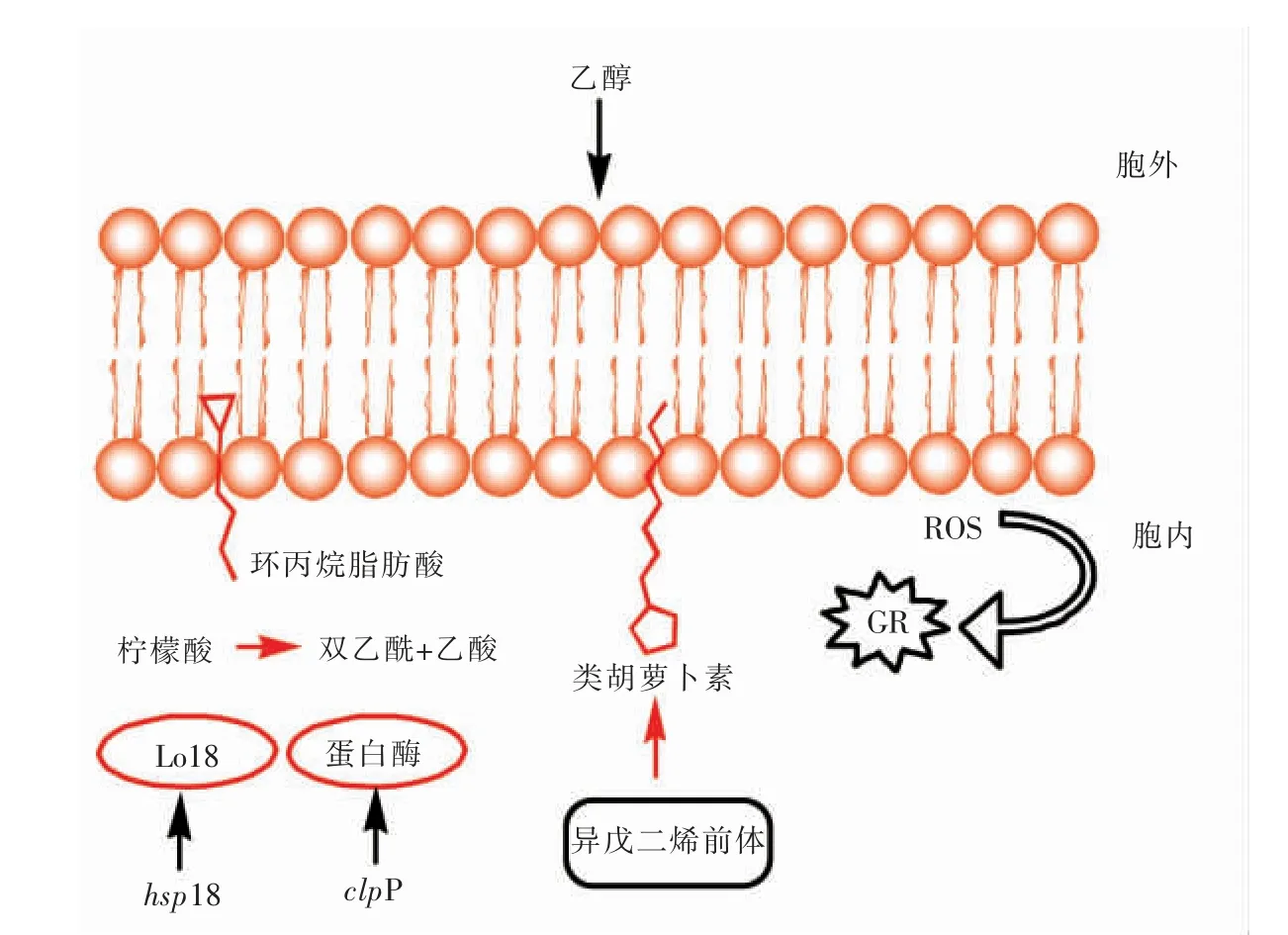
图1 乙醇胁迫后细胞适应性机制示意图Fig.1 Schematic diagram of cell adaptive mechanism after ethanol stress
此外,菌体细胞受到乙醇胁迫时,流动性发生变化会触发信号传导,从而诱导胁迫基因的表达。如,胁 迫 基 因hsp18[26]、ggpps[27]、clpP 等 的 表 达。hsp18 合成胁迫蛋白Lo18,此蛋白可作用于菌体细胞膜以防止蛋白质聚集而发生变性[28],利于维持细胞膜的完整性,从而使得菌体细胞在乙醇环境中存活。ggpps 基因是参与类胡萝卜素代谢的基因,当它过量表达时,可促使类异戊二烯前体转化为类胡萝卜素以提高细胞膜的稳定性,从而参与胁迫适应性反应,且乙醇浓度越高ggpps 基因的表达量就越高,其机制有待进一步探究。在酒酒球菌受到胁迫时,常伴随着分子伴侣的表达,分子伴侣是一类在序列上虽没有相关性但有共同功能的蛋白质,它们在细胞内帮助其它含多肽的结构完成正确的组装,而且在组装完毕后与其分离,不构成这些蛋白质结构执行功能的组分。在受到乙醇胁迫时,分子伴侣适应性机制(如clpP 等)会被诱导表达[29],这对维持细胞内蛋白质的正确组装有重要意义。此外,活性氧(Reactive oxygen species,ROS)在生物体内持续的产生,可导致DNA、蛋白质和脂质损伤,过量的活性氧会导致细胞受到氧化胁迫[30]。细胞受到乙醇胁迫时细胞内的活性氧产生会增加,谷胱甘肽还原酶(Glutathione reductase,GR)的含量也会增加,谷胱甘肽还原酶是一种利用还原型NADPH 将氧化型谷胱甘肽催化反应成还原型谷胱甘肽的酶。因此,细胞中GR 对维持细胞内氧化还原电位和抵制氧化胁迫有重要作用[31]。
2 低温对酒酒球菌的影响及胁迫反应机制
2.1 低温对酒酒球菌的影响
葡萄酒进行MLF 的启动途径有以下几种:自然诱发MLF;利用正在进行MLF 的酒体接种;使用实验室制备的菌株或商业化菌株接种(即直投式发酵剂);运用固定化细胞或固定化酶。目前,应用最为广泛是第3 种诱发途径。葡萄酒生境复杂,MLF 自然触发与发酵时间都难以控制,其次,自然触发MLF 对酒体的影响是非定向的,这与诱发菌种特性有关,如果MLF 不能顺利进行可能会给酒体带来病害影响。人为诱发时,可选择诱发的菌种与时间,有利于MLF 的控制,用于进行MLF 的直投式发酵剂主要为酒酒球菌冻干粉,因此探究低温对酒酒球菌的影响,对提高酒酒球菌启动MLF的顺利进行非常重要[32]。细胞经低温处理后,细胞膜中不饱和脂肪酸/饱和脂肪酸(UFA/SFA)比率上升,而膜中环丙烷脂肪酸(CFA)的含量增加,同时,细胞膜的流动性大大降低。Chu 等[19]研究发现细胞在经低温处理后,细胞膜会发生明显的硬化现象,然而其细胞活力未受到明显影响,且温度越低其硬化现象越明显。细胞经短时间低温处理后,将温度回升至原来温度时,其细胞膜的流动性也会恢复至原来状态,且细胞膜及蛋白质组分并未发生明显变化。即低温胁迫会影响细胞膜中脂肪酸及蛋白质组分,却不影响细胞的存活率,且此过程是可逆的。低温不仅会影响细胞膜的流动性,更会使得细胞内的酶活性暂时降低,抑制了细胞内各种生化反应的进行,当温度回升,这种抑制作用便会解除。
2.2 低温胁迫的适应性机制
细胞膜的流动性是生物膜正常功能的必须条件,如细胞物质运输、细胞信息的传导、膜上的酶活性等。受到低温胁迫时细胞膜组分调整是为了使其流动性保持在最适范围内,其最佳流动性约为水黏度的100 倍,相当于90%甘油水溶液的黏度。在该值附近,细胞膜可维持其结构的连续性,适合于膜蛋白的嵌入和运动。酒酒球菌细胞受到低温胁迫时,C19cyc11 的含量会增加,C16:0 的含量会降低,这样的脂肪酸组成有利于提高细胞膜的完整性和对抗胁迫环境的能力[33]。环丙烷脂肪酸的合成防止了细胞膜磷脂的堆积,且环丙烷脂肪酸比其它脂肪酸更加稳定,使细胞膜在低温下仍能保持较好的弹性和灵活性[24]。此外,酒酒球菌可产生胞外多糖(Exopolysaccharides,EPS),主要是荚膜类多聚体及葡聚糖。荚膜EPS 和可溶性EPS 的产生对酒酒球菌的细胞膜有保护作用,在低温胁迫时,这可对菌体细胞起到一定的保护作用,然而菌株所产生的EPS 对葡萄酒的质量有一定的影响[34-36]。总言之,低温胁迫对细胞的影响及机制的探究大多数停留在细胞膜组分的变化上,而并未进行深入的研究。
3 高温对酒酒球菌的影响及胁迫反应机制
3.1 高温对酒酒球菌的影响
高温会增加细胞膜的流动性及通透性,甚至导致细胞内代谢物流出,温度过高时会影响细胞膜中的酶活,进而影响细胞的存活率。在42 ℃条件下培养细胞时,菌体的生长明显受到抑制,且高温胁迫诱导了胁迫蛋白的合成[37]。这种机制使得酒酒球菌细胞能够存活并繁殖。并且酒酒球菌细胞经42 ℃处理后直投入葡萄酒中发酵可明显提高细胞在葡萄酒中的存活率以及诱发MLF 顺利进行的能力。酒酒球菌细胞受到高温胁迫时,细胞内的热休克蛋白含量增加,如Lo18[38-39]。且胁迫温度不同时,Lo18 在细胞上的分布存在差异。当温度升至42 ℃时,Lo18 大多数分布在细胞脂质体膜上,这表示Lo18 可直接作用于磷脂组分来增强分子顺序,防止蛋白质发生热聚集变性,保持细胞膜完整及稳定性。当处理温度高于33.8 ℃时,膜会发生硬化现象,当温度为15~33.8 ℃时,Lo18 对细胞膜没有显著影响。在不同胁迫温度下测定sHSP分子质量,当温度为4 ℃时,sHSP 的分子质量为1.5 Mu,30 ℃时sHSP 的分子质量为229 ku,40 ℃时为75 ku,45 ℃时为43 ku[38]。
3.2 高温胁迫的适应性机制
高温胁迫时,酒酒球菌细胞膜上的酰基链运动增加,利于保持较好的脂质顺序,以防止细胞膜流动性过高引起细胞内代谢物质流失。酒酒球菌可通过合成小热休克蛋白(Small heat shock protein,sHSP)来抵制高温胁迫环境,如Lo18[40]。在大多数分子伴侣上游都存在CtsR 基因,细胞膜流动性的变化会触发信号转导,从而诱导该调节子的表达,启动合成分子伴侣基因的表达[25]。细胞小热休克蛋白大多为分子伴侣,这种蛋白质广泛存在于可独立存活的微生物细胞中。受到高温胁迫时,随着温度的变化,Lo18 在细胞膜上的分布会发生相应的变化,这反映出不同温度对细胞膜的影响不同。Lo18 可根据细胞膜的变化调整自己在细胞膜上的分布情况,从而更有利于发挥其作用。温度较高时,Lo18 可直接作用于细胞膜上的脂质组分,来增强分子顺序防止蛋白质热聚集而变性,以维持蛋白质的正常功能,保证细胞在高温下的存活。此外,Lo18 的分子质量也根据温度的变化而有所改变,在一定范围内温度升高时,分子质量会降低,这样有利于更好的与细胞膜上的脂质分子相结合,更好的稳定细胞膜[40]。此外,在高温胁迫后菌体细胞内表达的多种胁迫蛋白被逐渐发现并了解,包括:ClpX,一种在指数生长期最先表达的热诱导蛋白,是调节ATP 酶亚基Clp-P 的分子伴侣[41];TrxA,类似于硫氧还蛋白,编码环蛋白,当环境中温度升高时其表达量会增加[42];FtsH,一种膜蛋白酶,在菌体受到高温胁迫或者精氨酸存在时表达量都会增加[43];OmrA,在高温胁迫下其表达量会增加[44]。
4 酸对酒酒球菌的影响及胁迫适应性机制
4.1 酸对酒酒球菌的影响
酒酒球菌生长的最适pH 值为4.8~5.5,而葡萄酒的pH 值通常低于3.5,即对酒酒球菌而言,葡萄酒是一个低pH 值的胁迫环境,pH 值的高低限制MLF 是否能够顺利进行[45]。当pH 值较低时,酒酒球菌细胞膜中乳杆菌酸的含量显著增加。Drici-Cachon 等[46]研究了3 株酒酒球菌在不同pH值条件下,膜脂肪酸的变化情况,结果表明pH 的变化对强嗜酸菌的膜脂肪酸影响较小,而对Lo107 和Lo8413(中性嗜酸菌)的影响幅度较大,且不饱和脂肪酸的不饱和度随着pH 的变化而变化。菌体细胞膜经pH 3.0 胁迫处理后细胞膜发生了硬化现象,pH 4.0 处理后的细胞膜流动性未出现明显变化,且酸胁迫对细胞膜的影响是不可逆的。不同pH 处理对细胞的存活率影响不同,经pH 3.0 处理30 min 后,细胞的存活率下降到75%,经pH 4.0 处理30 min 的细胞活力几乎保持100%。有研究对编码H+-ATP 酶的基因atp 进行分析,结果显示受到酸胁迫的菌体细胞其atp 的mRNA 含量要远高于未受到胁迫菌体细胞的mRNA 含量,受到胁迫后细胞中atp 基因表达量为正常环境中生存菌体细胞的3 倍[47]。
4.2 酸胁迫的适应性机制
酸胁迫时,细胞膜中不饱和脂肪酸的含量会变化,以维持细胞膜流动性位于最适范围。其中,乳杆菌酸含量会增加,乳杆菌酸有利于维持细胞膜的完整性及稳定性。酒酒球菌在酸胁迫条件下时,能诱导膜上H+-ATP 酶的活性增强。该酶不仅在MLF 过程中发挥作用而且在抗酸机制中也发挥重要作用。H+-ATP 酶可耦合ATP 水解将质子排除细胞外,从而调节细胞内外的pH 值,以维持细胞膜特性,以保证细胞的正常生长[48]。同时,膜上H+-ATP 活性越强,MLF 活性也越强,这为菌株在葡萄酒恶劣生境中的生长提供了能量保障。此外,菌体细胞受到酸胁迫时会诱导精氨酸脱亚胺酶(Arginine deiminase,ADI)代谢,将精氨酸转化为碱性产物而升高外部pH 值,从而增强了菌体细胞的耐酸性[49]。此外,精氨酸代谢是一个产能过程,与细菌的生长相偶联[50],使酒酒球菌在胁迫条件下具有更高的存活率。
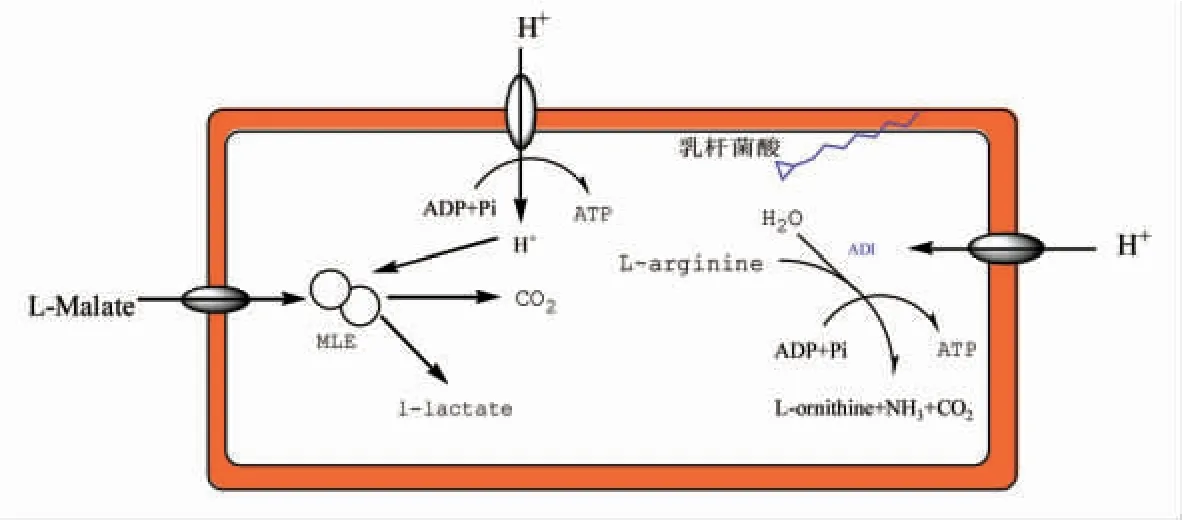
图2 酸胁迫后细胞适应性机制示意图Fig.2 Schematic diagram of cell adaptive mechanism after acid stress
5 存在问题与展望
随着生物信息学技术的发展,对酒酒球菌胁迫适应性机制逐渐转向基因组学、蛋白质组学及生物信息学研究,以期从基因表达方面了解胁迫反应的实质。在酒酒球菌受到胁迫后的生长过程中,参与胁迫反应的基因被发现并探究其发挥作用的机理。尽管很多胁迫基因已经被描述,但在恶劣的葡萄酒生境中,胁迫适应基因的调控机制,所参与的代谢通路仍需进一步研究[51]。例如,酒酒球菌对酒体中氨基酸和多肽的吸收至关重要,有利于其在胁迫环境中存活,然而目前对酒酒球菌在酒体中的氮代谢研究较少,如果能够探究得到菌体细胞在酸性环境下对多肽的利用机制,对进一步理解MLF 将会有非常重要的帮助[52]。
探究酒酒球菌胁迫反应基因的目标:明确菌体中胁迫反应基因的具体调控机制;对胁迫适应基因进行筛选、复制,研究其胁迫反应机理,并验证;发展生化手段筛选高耐受的菌株,以适应酒体的恶劣环境;发展生化或分子手段,对菌株在胁迫环境中的适应性进行评估;构建工程菌,使菌株具有抗酸,抗酒精,耐低温等特性,提高其在酒体中的存活率和MLF 能力。
