海南广藿香内生真菌分离鉴定及拮抗菌株筛选
2019-07-09易天凤吴友根于靖张军锋杨东梅
易天凤 吴友根 于靖 张军锋 杨东梅
摘 要 本研究从海南广藿香的根、茎、叶和花等组织中分离获得61株内生真菌,采用显微形态观察和ITS序列分析鉴定为15个属,其中优势菌群为炭疽菌属(Colletotrichum)和干酪酵母菌属(Meyerozyma)。以澳洲坚果叶枯病菌(Pestalotiopsis microspora)、西瓜枯萎病菌(Fusarium oxysporum)和火龙果溃疡病菌(Neoscytalidium dimi dia tum)为指示菌,采用平板对峙法进行拮抗菌株筛选,发现镰刀菌属(Fusarium)菌株PfuJ20、PfuG16和PfuG5对澳洲坚果叶枯病菌的抑制效果最好,棒孢属(Corynespora)菌株PfuH2对西瓜枯萎病菌抑菌效果较好。采用滤纸片扩散法进行各菌株对金黄色葡萄球菌、大肠杆菌、沙门氏菌、枯草芽孢杆菌等动植物病原细菌的抑菌作用评价,发现镰刀菌属PfuJ20、PfuG16对各细菌均有较强抑制效果,而链格孢属(Alternaria) PfuH1对金黄色葡萄球菌的生长抑制效果相对最强。结果表明,海南广藿香内生真菌具有一定的生物多样性,部分内生真菌菌株具有较强的抑菌活性。
关键词 广藿香;内生真菌;拮抗菌株
中图分类号 S476 文献标识码 A
Abstract In this study, 61 endophytic fungi were isolated from roots, stems, leaves and flowers of Hainan patchouli, and identified as 15 genera by ITS sequence analysis and morphological analysis. The dominant genus was the Colletotrichum and Meyerozyma. The inhibitory effect of Fusarium PfuJ20, PfuG16 and PfuG5 on Pestalotiopsis microspora was most obvious when Pestalotiopsis microspora, Fusarium oxysporum and Neoscytalidium dimidiatum were used as the indicator bacteria, and the activity of phytopathogenic fungi was determined by the plate sputum method. The activity of anti-phytopathogenic bacteria was determined by the filter paper diffusion method. Fusarium had strong inhibitory effects on the growth of Staphylococcus aureus, Escherichia coli, Salmonella and Bacillus subtilis and Alternaria had the strongest growth inhibitory effect on Staphylococcus aureus. The results showed that Hainan patchouli endophytic fungi had certain biodiversity and some endophytic fungal strains had more obvious antibacterial activity.
Keywords Pogostemon cablin; endophytic fungi; antagonistic strains
DOI 10.3969/j.issn.1000-2561.2019.08.021
廣藿香[Pogostemon cablin (Blanco.) Benth.]为唇形科刺蕊草属植物,原产于东南亚地区,如马来西亚、菲律宾、印度尼西亚等国,在我国引种可追溯到梁代或以前[1]。现主要分布于广东和海南2省,广东广州、肇庆、湛江以及海南万宁等地均有栽培。现今广藿香按产地可分为“广州石牌广藿香”(牌香)、“广东高要广藿香”(肇香)、“广东湛江广藿香”(湛香)和“海南广藿香”(南香)4种[2-3]。自古以来广藿香已广泛用作药材,在药用生产应用上有很大的前景,他具有芳香化浊、止呕及发表解暑之功效,常用于胸闷不舒、寒湿闭暑、腹痛吐泻、鼻渊头痛等方面疾病的治疗。现代研究表明广藿香具有抗菌、抗肿瘤、抗氧化、杀虫等作用[4],从广藿香中提取的精油在轻化工业上是重要的原料,可应用于化妆品、香水等的制作[5-6]。
植物内生真菌是指那些在其生活史的一定阶段或全部阶段生活于健康植物的各种组织和器官内部或细胞间隙的真菌,是植物微生态系统中的重要组成部分,部分菌株具有促进营养物质吸收、抗病虫害、抗逆境、促植物生长等作用[7]。目前,关于广藿香的报道多数集中在活性成分的研究上,包括Daldinia eschscholtzii、Cerrena sp. A593、Diaporthe longicolla A616和Bipolaris sorokiniana A606等内生真菌的活性成分已得到初步研究[8-11],但有关内生真菌在其与宿主植物组成的微生态系统中的作用机制还缺少研究。本研究选用海南广藿香(南香)作为研究材料,开展了内生真菌的分离鉴定及拮抗菌株筛选活动,为进一步开展相关内生真菌资源的收集与利用提供依据。
1 材料与方法
1.1 材料
1.1.1 植物材料 广藿香品种为南香,由海南大学热带农林学院提供,2017年2月种植于海南省儋州市(N20336,E1101925),常规管理,2018年5月取样。
1.1.2 培养基和试剂 孟加拉红平板培养基(用于分离内生真菌)、PDA固体培养基(用于保存内生真菌)、马铃薯葡萄糖肉汤(PDB)液体培养基(用于内生真菌发酵)、营养琼脂(NA)固体培养基(用于培养指示细菌),购自北京索莱宝Solarbio科技有限公司。真菌基因组DNA提取试剂盒、PCR 相关试剂等购买于北京索莱宝科技有限公司。引物由天一辉远生物科技有限公司合成(测序引物为ITS4:5′-TCCTCCGCTTATT MGA TA TGC-3′和ITS1f:5′-CTTGGTCATTTAGA GG AAG TAA-3′)。其他试剂均为国产分析纯。
1.1.3 供试菌株 真菌:澳洲坚果叶枯病菌,拟盘多毛孢(Pestalotiopsis microspora);西瓜枯萎病,尖胞镰刀菌(西瓜专化型)(Fusarium oxysporum);火龙果溃疡病,新暗色柱节孢菌(Neo scyt alidium dim dia tum)。细菌:金黄色葡萄球菌(Sta phylococcus aureus)、大肠杆菌(Escherichia coli)、沙门氏菌 、枯草芽孢杆菌(Bacillus subtilis)。供试菌株均由海南大学热带农林学院提供。
1.2 方法
1.2.1 海南广藿香内生真菌分离鉴定 参照马文婷等[12]的方法进行内生真菌分离。
显微形态观察:菌株接种培养基平板后,将盖玻片45~50插入固体培养基,待菌丝扩展至盖玻片处时取下,滴加棉兰染色剂,放在载玻片上用光学显微镜进行观察。
ITS序列分析:参照说明书用试剂盒提取各真菌菌株的基因组DNA,基于核糖体ITS区域的序列分析鉴定内生真菌[12]。PCR扩增产物送往铂尚生物技术(上海)有限公司进行测序。
1.2.2 内生真菌对植物病原真菌的拮抗作用评价 平板对峙法参照张冬静[13]的方法进行筛选。抑菌活性测试公式:
生长抑制率=(1-r/r0)100%式中,r为处理组病原菌生长半径;r0为对照组病原真菌生长半径。
1.2.3 内生真菌对动植物病原细菌的拮抗作用评价 发酵物制备:挑取5个纯化的内生真菌菌饼接入装有PDB培养基此处应补充说明培养基的体积(121 ℃、20 min灭菌)的250 mL锥形瓶中,放入摇床25 ℃恒温黑暗培养7 d左右。发酵液经两层滤纸片过滤,分别收集菌液与菌丝体,将乙酸乙酯加入菌液(1∶1,V/V)与干燥后粉碎的菌丝体,充分混匀后存放1 d,同样过程将菌液与菌丝体萃取3次收集有机相旋转蒸发至浸膏状,加甲醇配至20 mg/mL溶液,得到提取液。
抑菌测试:将浓度为(1105~1 107)CFU/mL的指示菌悬液均匀涂抹于营养琼脂固体培养基平板表面,再将浸泡于提取液24 h的滤纸片放置在平板表面,3次重复,以甲醇溶液为对照,37 ℃恒温培养1 d后测量抑菌圈直径。
1.3 数据处理
采用Excel软件处理数据,SAS 8.0软件进行方差分析。
2 结果及分析
2.1 海南广藿香内生真菌分离结果
通过平板分离法对广藿香内生真菌进行分离纯化,采用漂洗液和组织印记法检测表面消毒情況显示对照组无明显菌落长出。表明试验材料表面已消毒彻底,分离获得的菌株来自海南广藿香组织内部。从海南广藿香中总共分离出61株内生真菌,其中根部分离出14株,茎、叶、花分别分离出20、19、8株(表1)。代表性的海南广藿香内生真菌形态特征如表2所示。
2.2 海南广藿香内生真菌鉴定结果分析
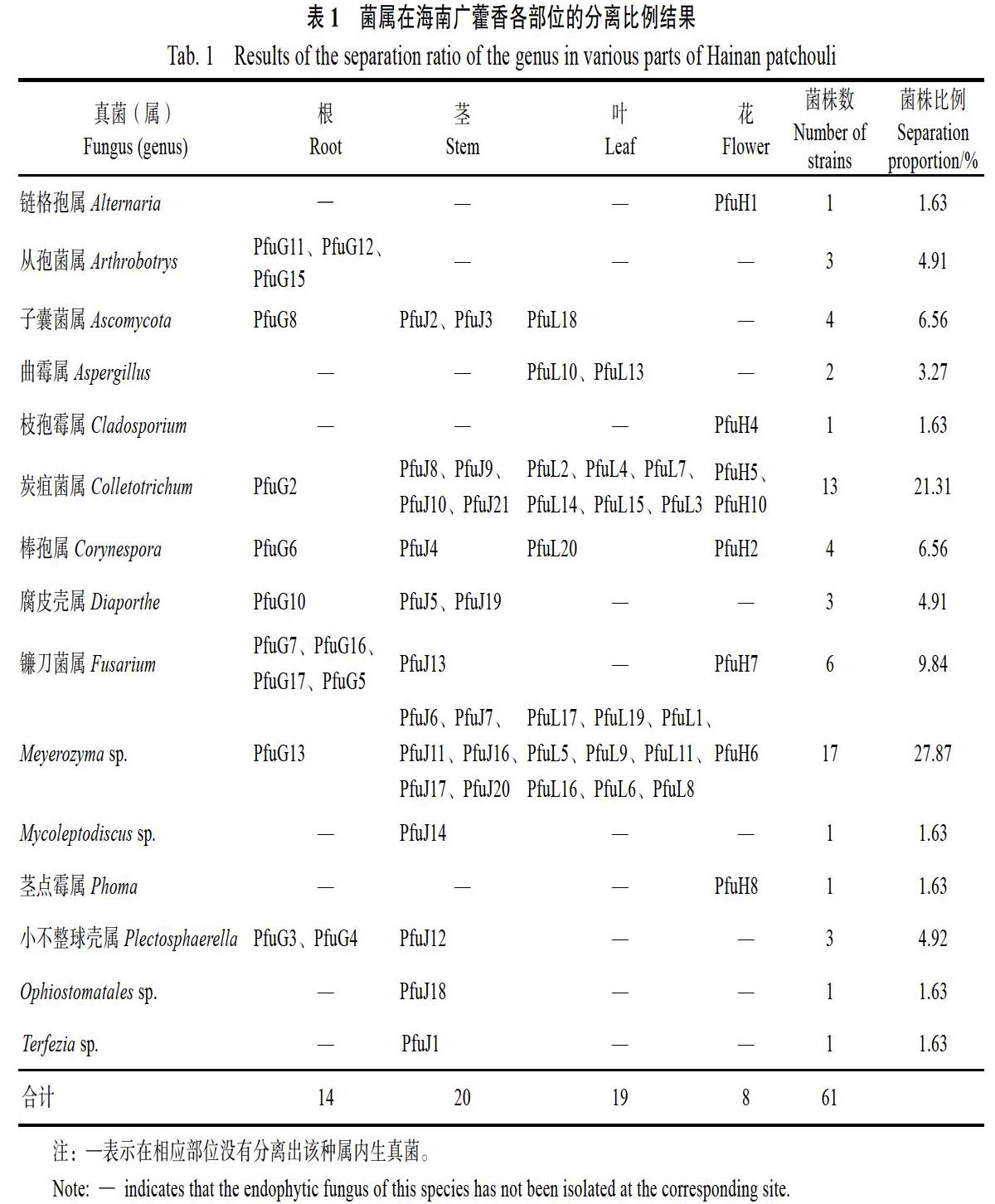
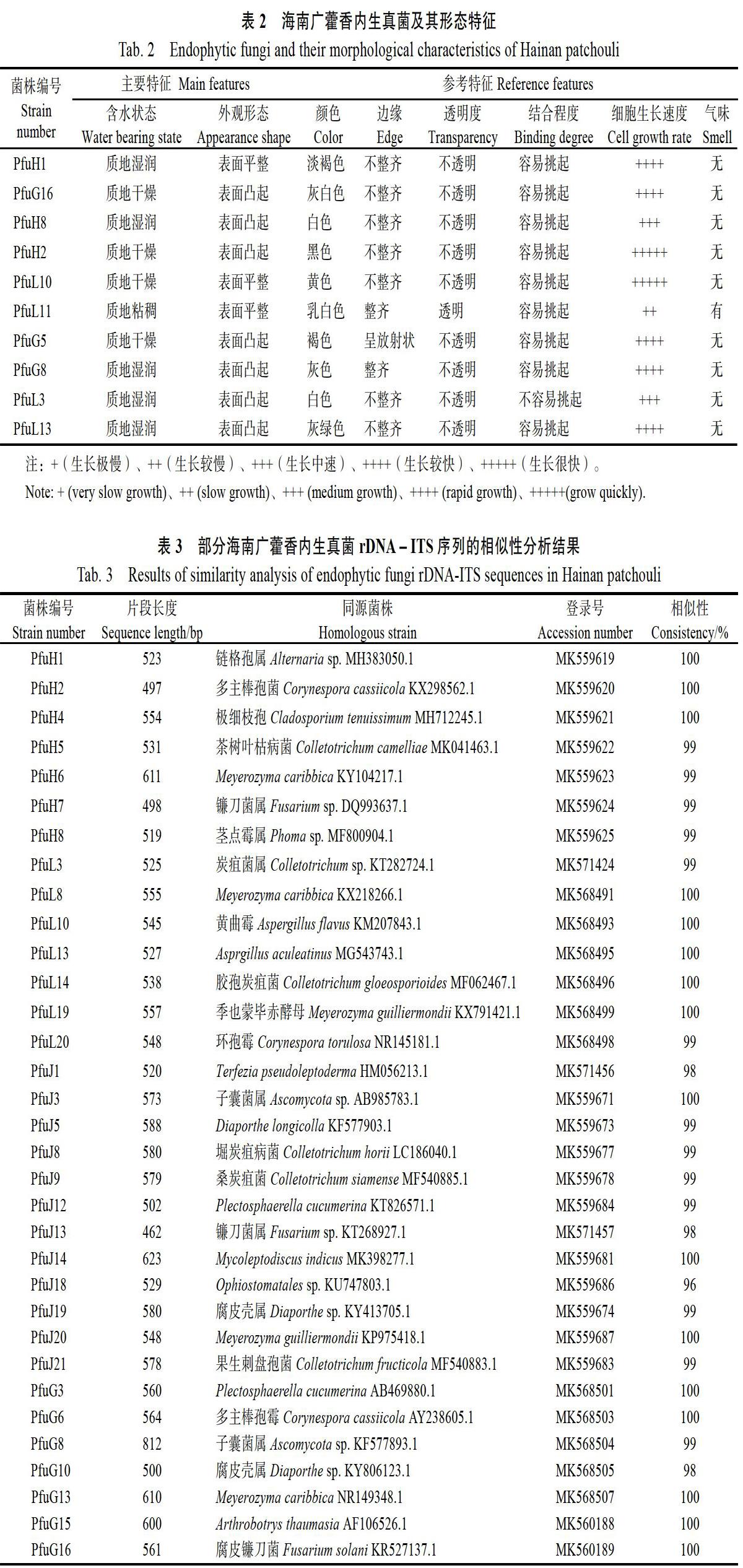
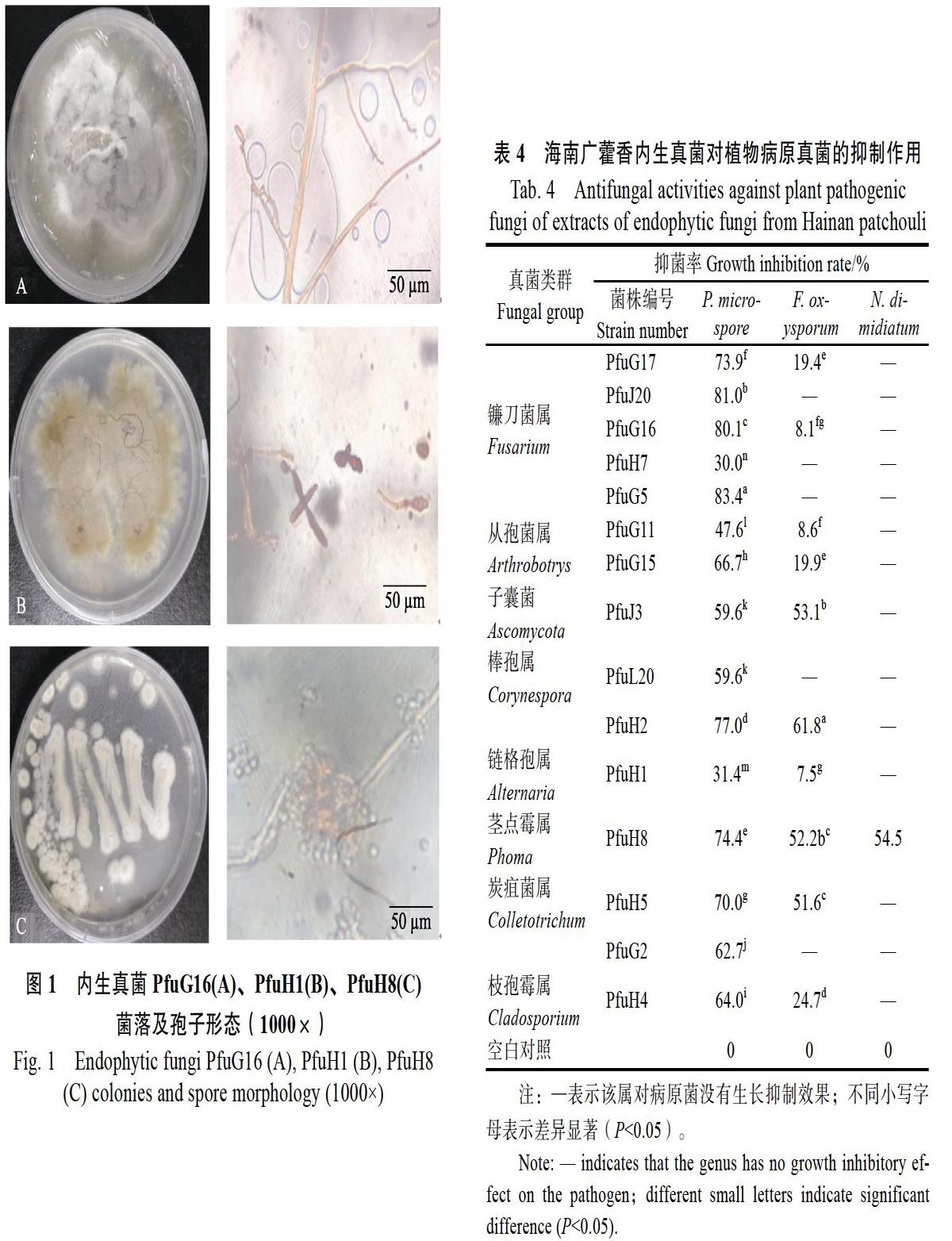
将分离得到的广藿香内生真菌进行DNA提取及rDNA-ITS序列PCR扩增,1.0%琼脂糖凝胶电泳显示,扩增序列条带在600 bp左右。部分海南广藿香内生真菌rDNA-ITS序列的相似性分析结果如表3所示,所获菌株序列长度在500 bp左右,且与同源菌株一致性都在90%以上。部分显微形态观察如图1:PfuG16菌丝有隔,分枝,分生孢子呈纺锤形,可初步判断为镰刀菌属;PfuH1分生孢子呈暗色的链状,且有明显的隔,则可鉴定为链格孢属;PfuH8分生孢子器球形,分生孢子单生于梗的末端呈椭圆形,可鉴定为茎点霉属。
本研究通过显微形态观察及分子鉴定的方法,分析出61株内生真菌为15个属,主要有炭疽菌属Colletotrichum、子囊菌属Ascomycota、棒孢属Corynespora、间座壳属Diaporthe、镰刀菌属Fusarium、从孢菌属Arthrobotrys等,表明海南广藿香内生真菌具有一定的生物多样性。通过对海南广藿香不同部位内生真菌的分离,发现不同组织部位的内生真菌种属分布不同,同一菌属也可存活在不同组织。炭疽菌属Colletotr ichum、棒孢属Corynespora在根、茎、叶、花中都有分布;链格孢属Alternaria、茎点霉属Phoma和枝孢霉属Cladosporium仅在花部分布;曲霉属Aspergillus仅分布于叶部;分离到的Plectos phaerella sp.、Terfezia sp.、Mycoleptodiscus sp.和Ophiostomatales sp.只分布于茎部。结果表明,海南广藿香不同部位分离的内生真菌种属存在差异,其中优势菌属为炭疽菌属(比例=21.31%)和Meyerozyma sp.(比例=27.87%)。
2.3 拮抗性內生真菌筛选
2.3.1 内生真菌对植物病原真菌的拮抗作用评价经拮抗作用评价后,得到15株对靶标病原真菌具有抑制效果的菌株(表4)。该15株内生真菌对P. microspore生长均有抑制作用,抑菌率都在30%以上,其中镰刀菌属的PfuJ20、PfuG16和PfuG5的抑菌效果最为显著,抑菌率均在80%以上;如图2可见,PfuL20拮抗P. microspora、PfuJ20拮抗P. microspora效果明显。子囊菌PfuJ3、棒孢属PfuH2、茎点霉属PfuH8和炭疽菌属PfuH5对F. oxysporum有较好的抑菌作用,抑菌率为50%以上,而内生真菌中仅茎点霉属PfuH8对N. dimidiatum有较好的抑菌作用(抑菌率为54.5%)。
2.3.2 抗动植物病原细菌活性结果分析 经过初筛得到10株对病原菌抑制效果明显的内生真菌,包含了8个属。由表5可知,抗S. aureus的菌株有8株,分别是PfuG8、PfuJ19、PfuH4、PfuG17、PfuG7、PfuL11、PfuJ6、PfuH1,占分离菌株总数的13.1%;抗E. coli的菌株有7株,分别是PfuG8、PfuJ3、PfuH4、PfuG17、PfuL11、PfuJ6、PfuH1,占分离菌株总数的11.4%;抗B. subtilis的菌株有4株,分别是PfuG8、PfuH4、PfuG17、PfuH1,占分离菌株总数的6.5%;抗S. enteritidis的菌株有5株,分别是PfuG8、PfuJ3、PfuL15、PfuG17、PfuH1,占分离菌株总数的8.2%;分别鉴定为镰刀菌属Fusarium与链格孢属Alternaria的PfuG17和PfuH1对供试菌株的生长抑制最为显著:PfuG17对供试细菌的抑菌圈直径可达到15 mm以上;H1对供试细菌的抑菌圈直径也可达11 mm以上,尤其对金黄色葡萄球菌(S. aureus)的抑制圈直径高达24 mm。图3为PfuH1和PfuG17菌株对病原细菌的拮抗作用,表明该菌株有明显的抗菌效果。
3 讨论
相关研究表明,内生真菌是普遍存在于植物组织内,且表现出多样性,大多数植物内生菌在植物不同的组织和器官内均有分布,而且它们的分布情况由宿主植物本身以及内生菌的种类所决定,在宿主植物不同器官的分布存在差异[14-15]。本研究在海南广藿香不同组织分离鉴定得到61株内生真菌,鉴定为15个属,且不同组织的分布存在差异,可能和不同组织提供的生长条件不同所造成的,吴婧波从杧果组织中分离得到53株内生真菌,其中胶孢炭疽菌(Colletotrichum gloeosporioides)、杧果球座菌(Guignardia mangiferae)、葡萄座腔菌(Botryosphaeria dothidea)为优势种,且各属在组织中分布也存在差异[16]。炭疽菌属内生真菌在药用草本植物中广泛分布[17],其在海南广藿香组织中同样属于优势菌属。谢华蓉的试验也发现广藿香内生真菌炭疽菌属(11.82%)为优势类群,链格孢属(占28.75%)、拟茎点霉属(占23.00%)的比例也高,存在这种差异可能是由于样品品种和环境条件的差异所造成的[18]。
内生真菌可产生多种活性物质,其具有抗病原微生物的作用。本研究通过抑菌测试实验发现镰刀菌属对澳洲坚果叶枯病(拟盘多毛孢)的生长抑制率高达80%,镰刀菌属在植物内生真菌中很常见,不少研究也发现该属产生的代谢产物对病原菌有一定的抑制作用。在广藿香中分离的内生真菌对荔枝霜疫霉霉(Peronop htho ra litchic)的抑制率高达95%,其中镰刀菌属抑制效果突出[19],其提取物浓度为50 g/L,试验结果差异说明抑菌效果与提取物浓度有密切关系。在进行海南广藿香内生真菌抑菌性测试时发现茎点霉属对澳洲坚果叶枯病的生长抑制显著,科研人员在进行该属内生真菌相关试验时也发现:从薄荷 (Mentha haplocalyx Briq. )中分离到的拟茎点霉属内生真菌能产生一种具有临床应用前景的新抗生素dicera ndorls[20];而且从银杏中分离的拟茎点霉属内生真菌中提取的杜松烷倍半萜也具有较强的抗菌作用[21]。这为海南广藿香拟茎点霉属内生真菌产生的活性物质研究提供了方向。在海南广藿香中分离得到的链格孢属对金黄色葡萄球菌也有显著的生长抑制效果,冯成亮等[22]从无花果中分离的链格孢属内生真菌可分离出对Valsa mali生长有明显抑制作用的次生代谢产物。由此可见,本研究对下一步内生真菌次生代谢产物的研究提供了试验基础。
当今已经有大量的研究工作者对内生真菌活性成分进行深入探索,此次研究在国内首次对广藿香花的内生真菌进行探索,研究结果表明,广藿香内生真菌在组织内表现出多样性,且从广藿香中分离的内生真菌部分具有较强抑菌活性,因此研究广藿香内生真菌的抑菌性为开发有效化合物及广藿香资源利用提供了理论依据,对于工业生产具有重要意义。
参考文献
吴友根, 郭巧生, 郑焕强. 广藿香本草及引种历史考证的研究[J]. 中国中药杂志, 2007, 32 (20): 2114-2117, 2181.
吴友根, 郭巧生, 郑焕强.广藿香种植土壤和药材中有机氯农药及重金属残留分析[J]. 中国中药杂志, 2008, 33 (13): 1528-1532.
Ito K, Akahoshi Y, Ito M, et al. Sedative effects of inhaled essential oil components of traditional fragrance Pogostemon cablin leaves and their structure-activity relationships[J]. Journal of Traditional and Complementary Medicine, 2016, 6(2): 140-145.
Chen Y, Wu Y, Xu Y, et al. Dynamic accumulation of sesquiterpenes in essential oil of Pogostemon cablin[J]. Revista Brasileira de Farmacognosia, 2014, 24(6): 626-634.
何國振, 李锦坤, 高 伟, 等. 广东省广藿香种植业发展策略[J]. 中国农学通报, 2012, 28(31): 288-292.
陈蔚文, 徐鸿华. 南药资源的保护与可持续利用研究[J]. 广州中医药大学学报, 2009, 26(3): 201-203.
Li Y, Ma C, Qian M, et al. Butein induces cell apoptosis and inhibition of cyclooxygenase-2 expression in A549 lung cancer cells[J]. Molecular Medicine Report, 2014, 9(2): 763-767.
卢梦梦, 陈玉婵, 刘洪新, 等. 广藿香内生真菌Daldinia eschscholtzii A630次级代谢产物及其细胞毒活性研究[J].天然产物研究与开发, 2018, 30(7): 1176-1180.
谢佳倩, 陈玉婵, 孙章华, 等. 广藿香内生真菌Cerrena sp.中倍半萜类成分及其细胞毒活性研究[J]. 中国中药杂志, 2017, 42(9): 1693-1698.
王 沫, 陈玉婵, 孙章华, 等. 广藿香内生真菌Diaporthe longicolla A616细胞毒活性次级代谢产物研究[J]. 中国中药杂志, 2016, 41(11): 2112-2117.
王 沫, 陈玉婵, 李浩华, 等. 广藿香内生真菌Bipolaris sorokiniana的次级代谢产物及其抗肿瘤活性研究[J]. 广东药学院学报, 2015, 31(5): 576-580.
马文婷, 吴友根, 胡征波, 等. 诺丽内生真菌的分离鉴定及其次生代谢产物的抗氧化活性[J]. 热带生物学报, 2017, 8(4): 424-430.
张冬静. 药用植物内生真菌的分离及其抑菌活性筛选[D].长春: 吉林大学, 2015.
刘 波, 郑雪芳, 孙大光, 等.柑橘黄龙病株不同部位内生细菌群落结构的多样性[J].生态学报, 2011, 31(24): 7325-7342.
严 亮, 杨瑞娟, 王桥美.云南铁皮石斛内生菌的分离与鉴定[J]. 云南农业大学学报(自然科学), 2015, 30(5): 760-765.
吴婧波, 詹儒林, 柳 凤, 等. 杧果内生真菌分离鉴定及多样性分析[J]. 果树学报, 2013, 30(6): 1030-1039.
谭小明, 周雅琴, 陈 娟, 等. 药用植物内生真菌多样性研究进展[J]. 中国药学杂志, 2015, 50(18): 1563-1580.
谢华蓉, 徐在超, 刘 军, 等. 广藿香内生真菌多样性及其对青枯菌的拮抗活性[J]. 微生物学通报, 2017, 44(5): 1171-1181.
王 营, 李浩华, 谭国慧, 等. 广藿香内生真菌类群分析及其抗菌活性研究[J]. 中国中药杂志, 2017, 42(4): 657-662.
Wagenaar M M, Clardy J. Dicerandrols. new antibiotic and cytotoxic dimers produced by the fungus Phomopsis logicolla isolated from an endangered mint[J]. Journal of Natural Products, 2001, 64(8): 1006-1009.
Silva G H, Teles H L, Zanardi L M, et al. Cadinane sesquiterpenoids of Phomopsis cassia, an endophytic fungus associated with Cassia spectabilis (Leguminosae)[J]. Phytochemistry, 2006, 67(17): 1964-1969.
冯成亮, 马养民. 无花果内生真菌Alternaria sp. FL25次生代谢产物的分离及对植物病原菌的抑制活性[J].应用与环境生物学报, 2010, 16 (1): 76-78.
