广东花生黑腐病病原菌鉴定及防治药剂的室内毒力测定
2019-06-11李娜王泽钿何桂碧向梅梅张云霞
李娜 王泽钿 何桂碧 向梅梅 张云霞

摘 要 花生黑腐病是花生的重要病害,对生产造成严重影响,被列为我国进境植物检疫性病害。为明确花生黑腐病的病原菌种类和筛选有效防治药剂,采用形态学特征与分子生物学技术相结合的方法,对花生黑腐病害进行了病原菌鉴定,同时采用菌丝生长速率法测定了8种杀菌剂对花生黑腐病菌的抑制作用。结果表明:花生黑腐病害的病原菌为冬青丽赤壳Calonectria iliciola。8种杀菌剂室内毒力测定结果显示,10%戊唑醇EC对花生黑腐病菌的毒力最强,EC50值为1.12 μg/mL;300 g/L苯甲·丙环唑EC、29%吡萘嘧菌酯SC、25%咯菌腈SC、15%吡唑嘧菌酯SC和22.5%啶氧菌酯SC对花生黑腐病菌也有较好的抑制作用,EC50均低于10 μg/mL;30%精甲恶霉灵的抑制作用最差,EC50值为76.71 μg/mL。以上研究结果可为花生黑腐病的防治提供理论依据。
关键词 花生;黑腐病;杀菌剂;毒力测定
中图分类号 Q93 文献标识码 A
花生(Arachis hypogaea)是我国重要的油料作物,黑腐病是影响花生生产的重要病害之一。1965年,美国乔治亚州首次发现该病害,随后,迅速扩散至美国东南部的花生种植区,受害严重时损失率高达50%[1-2]。目前该病害分布于美国、日本、韩国、澳大利亚等10多个国家[3]。2009年,我国广东省首次发现花生黑腐病,随后,在福建、江西和云南等省先后发现该病害[4-7]。2010年,广东省农业厅将该病原菌增列入《广东省农业植物检疫性有害生物补充名单》。
花生黑腐病主要通过土壤和种子带菌传播,病害一旦发生,防控极为困难。目前花生黑腐病没有高抗的品种,农业防治极其困难,土壤熏蒸具有一定的效果,但费用高且易造成环境污染[8],迄今没有有效的防治措施。本文拟在病原菌鉴定的基础上,对病原菌进行杀菌剂的室内毒力测定和有效药剂的筛选,为生产上病害的防治提供科学依据。
1 材料与方法
1.1 材料
1.1.1 供试菌株 供试菌株于2017年5月采自广东省翁源县三华镇,从发病组织的根茎部分离获得病原菌。
1.1.2 供试培养基 病原菌分离用培养基为马铃薯葡萄糖培养基(PDA)、菌落形态鉴定培养基为麦芽汁培养基(MEA),显微形态观察培养基为康乃馨叶片培养基(CLA)。单菌丝分离培养基为水琼脂培养基(WA)。
1.1.3 供试药剂 供试杀菌剂共8种:30%精甲噁霉灵水剂、10%戊唑醇乳油,浙江禾本科技有限公司;22.5%啶氧菌脂酯悬浮剂,上海农化生化有限公司;20%噻夫吡唑醚悬浮剂、15%吡唑醚菌酯悬浮剂,北京燕化永乐生物科技有限公司;29%吡萘嘧菌脂悬浮剂、300 g/L苯甲·丙环唑乳油、25%咯菌腈悬浮剂,瑞士先正达作物保护有限公司。
1.2 方法
1.2.1 病原真菌的分離和纯化 参照方中达[9]的组织分离法:新鲜病株的茎基部病组织切成3 mm×3 mm小块,依次用75%酒精消毒10~15 s、2.5%次氯酸钠消毒25~35 s,无菌水洗涤3次,再用无菌滤纸吸干水分,移至PDA平板、25 ℃培养。待菌落长出后,纯化并保存菌种。
单菌丝分离:挑取菌落边缘少量菌丝,放入WA培养基上,25 ℃培养24 h,于显微镜下切取单根菌丝,放入MEA培养基上,25 ℃恒温黑暗培养。
1.2.2 病原菌的致病性测定 参照袁汇涛[10]的方法:花生种子水培发芽后,选取长势好的幼芽,于菌丝悬浮液中浸泡10 min,以无菌水处理为对照,每个处理接种5株。将接种处理后的幼芽插入水培棉中,移入装有水的托盘中,25 ℃光照培养,观察发病情况。
1.2.3 病原真菌的形态学鉴定 参照Lombard等[11]的方法。在培养5 d后获得单菌丝菌落的边缘打孔,取菌饼置于新的MEA培养基上,于25 ℃条件下培养,观察菌落特征。取菌饼置于CLA培养基上,于25 ℃条件下培养,待产孢后于光学显微镜下观察其显微形态结构,拍照并测量分生孢子、产孢细胞和泡囊大小。
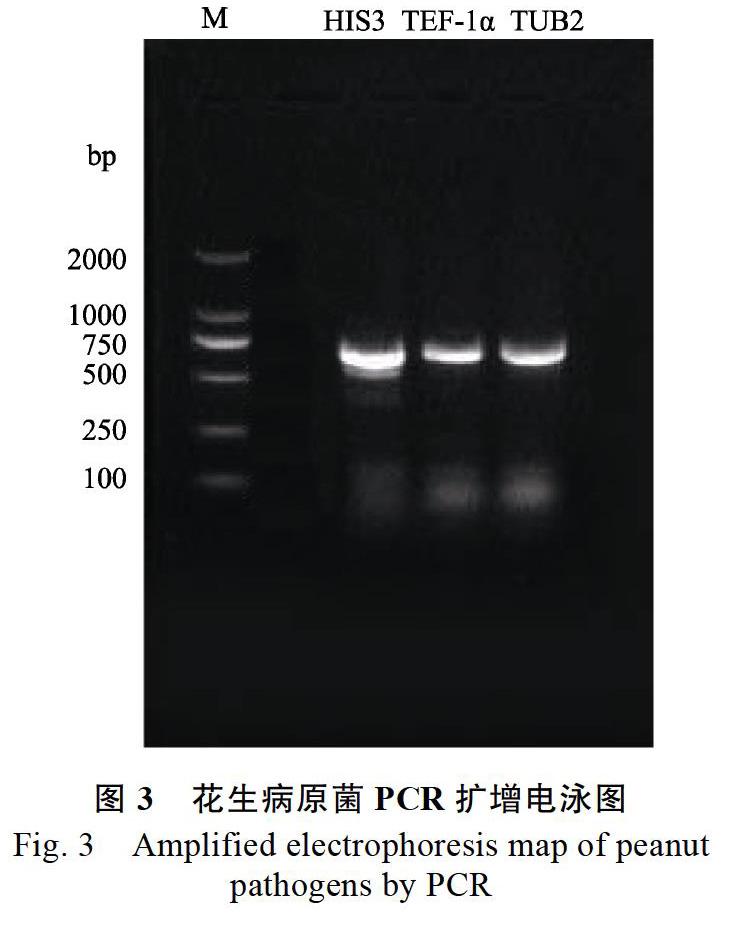
1.2.4 病原真菌的分子鉴定 采用CTAB法提取待鉴定菌株的DNA。参照Carbone等[12]的方法,选取组蛋白H3(histone H3,HIS3)、翻译延伸因子-1α(translation elongation factor 1-alpha,TEF- 1α)和β-微管蛋白(β-tubulin,TUB2)共3个基因序列进行PCR扩增。
特异性引物:HIS3选用引物CYLH3F和CYLH3R[13]。TEF-1α选用引物EF1-728F[12]和引物EF2[14]。TUB2选用引物CYLTUB1R[13]和T1[15]。
PCR反应体系(总体积25 μL):2×Taq PCR Green Mix 12.5 μL,DNA模板2 μL,10 μmol/L上游引物和下游引物各1 μL,ddH2O 8.5 μL。
His3 和TEF-1α的PCR扩增程序:94 ℃预变性3 min,94 ℃变性30 s,48 ℃退火30 s,72 ℃延伸1 min,35个循环;72 ℃终延伸10 min。TUB2的退火温度为50 ℃。
将得到的PCR扩增产物送至上海英潍捷基贸易有限公司测序,最后将获得的序列在NCBI网站上进行BLAST同源性比对分析。
1.2.5 室内毒力测定试验 含毒培养基的配制:每种供试药剂按有效成分含量分别用无菌水稀释成一定浓度梯度的母液,向PDA培养基中加入已配制好的杀菌剂母液,制成不同浓度梯度的含药平板。根据预试验结果每种药剂设置5个处理浓度(100、10、1、0.1、0.01 μg/mL)。CK加入等量的无菌水于PDA培养基作空白对照。
菌落测定方法:供试菌株培养5 d后,用直径5 mm的打孔器在菌落边缘打孔,取菌饼接种于含毒培养基中央,每个处理5个重复。接种后的平板置于25 ℃培养箱中培养。待对照菌落长满培养皿时,采用“十”字交叉法测量各处理的菌落直径,计算平均值,得出抑菌率。抑菌率=(对照组菌落直径药剂处理组菌落直径)/(对照组菌落直径菌饼直径)100%。
以药剂浓度对数值为横坐标(x),以校正抑菌率几率值为纵坐标(y),得到线性回归方程y=a+bx和相关系数r值。计算供试药剂对花生黑腐病菌的抑菌中浓度EC50及EC95值,比较各供试药剂的抑制效果及花生黑腐病菌对各供试药剂的敏感性。
2 结果与分析
2.1 花生黑腐病菌的病原菌鉴定
2.1.1 花生黑腐病的症状及致病性测定 花生幼苗期和成株均可受害,受害茎基部和果针上出现黑色的凹陷病斑,荚果和根受害后期会变黑腐烂,植株叶片变黄、萎蔫,严重时枯死。在潮湿状况下,发病部位产生红色的子囊壳(图1)。
健康的花生幼苗在接种5 d后接种部位开始出现褐色小病斑,病斑逐渐扩大蔓延至整条根变褐色,后期植株萎蔫。接种发病症状与田间病状一致。对照组未发病。
从接种发病的植株进行再分离,得到的菌落形态和显微形态与接种菌株一致,说明分离菌株为花生黑腐病的病原菌。
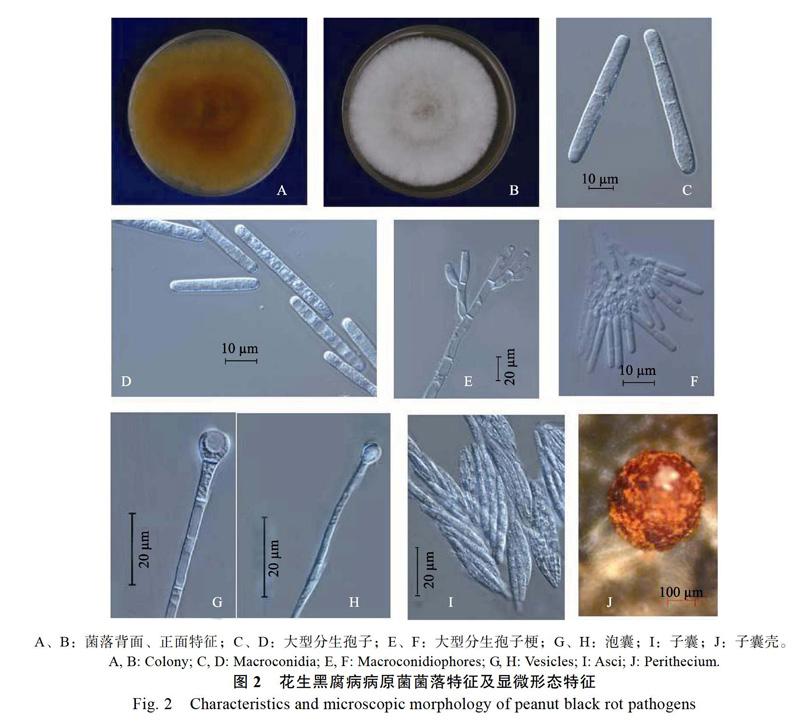
2.1.2 花生黑腐病病原菌的形态学鉴定 MEA培养基上菌落初期为白色,逐渐变为灰白色至浅褐色,菌落背面中央红褐色,边缘白色。气生菌丝绒毛状,茂盛,菌落边缘较整齐(图2A,图2B)。
在CLA培养基上培养,分生孢子梗多级帚状分枝,分枝末端产生2~4个瓶梗,大小为(10~25)μm(3~6.5)μm;末端泡囊球形或近球形,大小为(15~28)μm(5.5~15)μm,延伸梗具隔膜,大小为(200~280)μm(3.5~6.5)μm;大型分生孢子直,圆柱形,无色透明,一端稍窄,大小为(60~75)μm(7.5~8.0)μm,1~3个隔膜,隔膜处缢缩不明显。子囊壳卵球形或近球形,红褐色,单生或聚生,大小为(300~600)μm(300~500)μm;子囊棍棒状,每个子囊含8个子囊孢子;子囊孢子纺锤形,直或稍弯,无色透明,两端钝圆,1~3个隔膜,隔膜处稍有缢缩,大小为(30~85)μm (5~9.5)μm(图2C~图2J)。其显微形态结构与Pan等[4]报道的冬青丽赤壳Calon ectria ilicicola基本一致。
2.1.3 花生黑腐病病原菌的分子鉴定 利用翻译延伸因子-1α(TEF-1α)蛋白H3(HIS3)和β-的DNA序列在NCBI网站上进行BLAST比对,实验菌株ws(NCBI登录号为MK120359、MK120360、MK120361)与Calonectria ilicicola(NCBI登录号为AY725728.1、GQ267256.1、EF159730.1)的相似度最高,都為99%。因此,根据形态学特征和DNA序列分析,将分离的花生黑腐病的病原菌鉴定为C. ilicicola。
2.2 花生黑腐病菌毒力测定
2.2.1 供试药剂对花生黑腐病菌的毒力活性 EC50值的大小是衡量杀菌剂对病原菌毒力作用的重要指标。8种供试药剂对花生黑腐病菌表现出不同的毒力作用,抑菌效果存在明显的差异(表1)。其中,戊唑醇对花生黑腐病菌毒力最高,EC50为1.12 μg/mL,其次为苯甲·丙环唑、吡萘嘧菌酯、咯菌腈、吡唑嘧菌酯、啶氧菌酯,EC50均低于10 μg/mL,精甲噁霉灵对花生黑腐病菌毒力最低,EC50为76.71 μg/mL。
2.2.2 花生黑腐病菌对杀菌剂敏感性比较 毒力测定试验中,回归方程的斜率越大说明病原菌对药剂的反应敏感性越强。由表1可知,花生黑腐病菌对吡萘嘧菌酯最敏感,其次为戊唑醇、精甲噁霉灵、苯甲·丙环唑、噻夫吡唑醚,花生黑腐病菌对吡唑嘧菌酯和啶氧菌酯敏感性较低。
3 讨论
本研究通过对标准条件下菌落特征、形态学特征及3个基因的DNA序列分析,明确了采自广东省翁源县的花生黑腐病菌为冬青丽赤壳(Calonectria iliciola)。该结果和Pan等[4]的研究一致。前人研究中,花生黑腐病菌多以其无性态命名,为寄生帚梗柱孢菌(Cylindrocladium parasiticum),按照真菌分类命名的最新法则[16],该属真菌应以有性态命名,所以花生黑腐病菌应订正为冬青丽赤壳(Calonectria iliciola)。
已有研究表明,三唑类的杀菌剂对Calonectria引起的植物病害具有较好的效果。马海宾等[17]在研究杀菌剂对桉树焦枯病菌(Cy. quinqueweptatum)毒力作用时发现,供试的6种杀菌剂中戊唑醇对桉树焦枯病菌的毒力最高,EC50为4.1237 mg/L。在林地防效中,丙环唑使桉树林地的病情指数下降33%。本研究中,戊唑醇对花生黑腐病菌毒力最高,其次为苯甲·丙环唑,研究结果与其一致。由于杀菌剂的室内毒力测定结果不能完全反映其大田的防效,因此戊唑醇对花生黑腐病菌的田间防效还需要进一步大田验证。
花生黑腐病菌在我国南方地区发生普遍,生产中应引起重视。我国从2009年在广东首次发现该病害后,相继在福建、江西和云南等地发现有该病害危害[4-7]。本文作者前期调查发现,该病害在广东省的大部分花生产区均有发生。目前对花生黑腐病的研究主要局限于病原菌上,而对该病的防治技术研究较少,仅袁汇涛[10]和蓝国兵等[18]通过人工接种方法对花生黑腐病的抗病品种进行了筛选,对花生黑腐病药剂方面的防治还未见报道。本研究结果可为花生黑腐病的药剂防控提供一定的参考。
参考文献
Bell D K, Sobers E K. A peg, pod and root necrosis of peanuts caused by a species of Calonectria[J]. Phytopathology, 1966, 56(12): 1361-1364.
Wright L P, Andrew J D, Wingfield B D, et al. Population structure of Cylindrocladium parasiticum infecting peanuts in Georgia, USA[J]. European Journal of Plant Pathology, 2010, 127(2): 199-206.
潘汝谦, 徐大高, 邓铭光, 等. 外来入侵花生黑腐病菌在中国的风险性评估[J].中国农业科学, 2012, 45(15): 3068-3074.
Pan R, Guan M, Xu D, et al. Cylindrocladium black rot caused by Cylindrocladium parasiticum newly reported on peanut in China[J]. Plant Pathology, 2009, 58(6): 1176.
Pan R, Deng Q, Xu D, et al. First report of Cylindrocladium black rot of peanut caused by Cylindrocladium parasiticum in Fujian province, Eastern China[J]. Plant disease, 2012, 96(4): 583.
Gai Y, Deng Q, Pan R, et al. First report of Cylindrocladium black rot of peanut caused by Cylindrocladium parasiticum (teleomorph Calonectria ilicicola) in Jiangxi province, China[J]. Plant disease, 2012, 96(4): 586.
Pei W H, Cao J F, Yang M Y, et al. First report of black rot of Medicago sativa Caused by Cylindrocladium parasiticum (teleomorph Calonectria ilicicola) in Yunnan Province, China[J]. Plant Disease, 2015, 99.
Phipps P M. Control of Cylindrocladium black rot of peanut with soil fumigants having methyl isothiocyanate as the active ingredient[J]. Plant Disease, 1990, 74: 438-441.
方中達. 植病研究方法[M]. 北京: 中国农业出版社, 1998.
袁汇涛. 花生黑腐病抗病品种筛选及抗病机制初步研究[D]. 广州: 仲恺农业工程学院, 2016.
Lombard L, Crous P W, Wingfeld B D, et al. Phylogeny and systematics of the genus Calonectria[J]. Studies in Mycol ogy, 2010, 66(3): 31-69.
Carbone I, Kohn L M. Method for designing primer sets for speciation studies in filamentous ascomycetes[J]. Mycologia, 1999, 91(3): 553-556.
Crous P W, Groenewald J Z, Risede J M, et al. Calonectria species and their Cylindrocladium anamorphs: species with sphaeropedunculate vesicles[J]. Studies in Mycology, 2004, 50: 415-430.
Donnell K O, Kistler H C, Cigelnik E, et al. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitocho ndrial gene genealogies[J]. Proceedings of the National Academy of Sciences, 1998, 95(5): 2044-2049.
Donnell K O, Cigelnik E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous[J]. Molecular Phylogenetics and Evolution, 1997, 7(1): 103-116.
Wingfield M J, De Beer Z W, Slippers B, et al. One fungus, one name promotes progressive plant pathology[J]. Molecular Plant Pathology, 2012, 13 (6): 604-613.
马海宾, 康丽华, 崔宁宁, 等. 杀菌剂对桉树焦枯病菌的毒力测定及林地防病效果[J]. 热带作物学报, 2010, 31(11): 2006-2009.
蓝国兵, 何自福, 佘小漫, 等. 广东花生主要品种对黑腐病菌的抗性水平鉴定[J]. 生物安全学报, 2016, 25(3): 214-217.
