三角鲂×翘嘴鲌、团头鲂×翘嘴鲌两种杂交后代微卫星遗传结构分析
2019-03-11苏晓磊郑国栋蒋霞云邹曙明
苏晓磊 郑国栋 蒋霞云 邹曙明
(上海海洋大学, 农业部团头鲂遗传育种中心, 农业部淡水水产种质资源重点实验室,水产科学国家级实验教学示范中心, 上海 201306)
鱼类的远缘杂交是一种育种的重要手段, 可以将亲本的优良性状结合, 增加遗传多样性, 从而获的优良的杂交新品种[1, 2]。远缘杂交已广泛应用于鱼类遗传育种, 自20世纪50年代末, 中国已做了100多个杂交组合, 包括3个目, 7个科, 共40多种鱼类[3]。有些组合具有杂种优势。目前对于团头鲂(Megalobrama amblycephala)和翘嘴鲌(Erythroculter ilishaeformis)、三角鲂(Megalobrama terminalis)和翘嘴鲌杂交研究还比较少。葛玲瑞等[4]通过翘嘴红鲌(♂)和团头鲂(♀)的杂交, 得到了二倍体与三倍体鲂鲌。在翘嘴红鲌(♂)×团头鲂(♀)杂交子代的含肉率方面, 金万昆等[5]也做过一些检测, 其氨基酸含量中很多指标都高于其他鱼类。鳊鲂杂交方面山西大学生物系[6]进行了简单的研究, 其杂种肉质鲜美, 生长较快。团头鲂(♂)×翘嘴红鲌(♀)杂种F1鳞片较大且不易脱落, 耐操作, 顾志敏等[7]在杂种F1形态特征及遗传分析方面的研究证实了这一点。郑国栋等[8]利用团头鲂(♀)和翘嘴鲌(♂)进行杂交, 杂交种有良好的生长优势, 且耐低氧和耐运输能力也比亲本有所提高。通过在营养价值方面对三角鲂 (♀)×翘嘴红鲌 (♂) F1的必需氨基酸和亲本的肌肉组成进行研究, 马恒甲[9]得出翘嘴红鲌(♂)×三角鲂 (♀) 的杂种F1为较优质鱼类, 但其并没有在遗传结构上进行分析。从这些研究可以看出,杂交后代确实有明显的杂交优势。团头鲂和三角鲂属鲂属, 而翘嘴鲌属鲌属, 它们在我国淡水养殖鱼类, 都占有很重要的地位, 但在形态结构上有着很大差异, 因此遗传结构也比较复杂。通过对团头鲂[10]♀×翘嘴鲌♂和三角鲂×翘嘴鲌♂的杂交后代的微卫星分析, 了解其遗传结构以及杂交优势, 为今后团头鲂×翘嘴鲌和三角鲂×翘嘴鲌的杂交育种提供了理论依据。
1 材料与方法
1.1 实验材料
杂交试验所用亲本为“浦江1号”团头鲂, 东江三角鲂和淀山湖翘嘴鲌。杂交试验利用团头鲂雌性亲本30尾与翘嘴鲌雄性亲本30尾进行杂交获得“团头鲂♀×翘嘴鲌♂”杂交种群体, 利用三角鲂雌性亲本30尾与与翘嘴鲌雄性亲本30尾进行杂交获得“三角鲂♀×翘嘴鲌♂”杂交种群体。2个杂交种群体及其亲本均保存于上海海洋大学水产动物遗传育种中心。3个亲本群体及2个杂交群体各随机剪取30尾鱼鳍, 共150尾, 放入95%的酒精中, 保存于-20℃备用。
1.2 微卫星引物
本研究从60对SSR引物中筛选出16对效果较好的引物。其中10对SSR引物是由Tang等[11]通过5′锚定PCR技术获得, 2对是Li等[12]通过磁珠富集的技术开发设计的SSR引物, 另外4对EST-SSR引物是Gao等[13]通过转录组测序分析而获得。SSR位点及引物信息见表 1。
1.3 基因组DNA的制备
使用北京天根生物科技有限公司生产的海洋动物组织基因组DNA提取试剂盒(离心柱型), 参照其说明书介绍的方法提取基因组DNA, 基因组DNA提取完成后, 用0.8% 的琼脂糖凝胶电泳和分光光度计检测DNA质量和浓度, -20℃保存备用。
1.4 PCR反应体系与扩增程序
参照张倩倩等[14]反应体系10 μL, 包含5 μL含染料的2×TaqPCR MasterMix (TaqDNA Polymerase: 0.1 U/μL; MgCl2: 4 mmol/L; dNTPs each:0.4 mmol/L), 上下游引物各0.5 μL (10 μmol/L),0.5 μL模板DNA (30—50 ng), 3.5 μL ddH2O。PCR反应Eppendorf Master cycler ep gradients型PCR仪上进行, 反应程序为94℃预变性5min, 94℃30s, 50—65℃(退火温度参照表 1中的引物信息进行调整) 30s, 72℃ 30s, 30 个循环, 最后 72℃延伸10min。

表 1 微卫星引物特征Tab. 1 Characteristics of microsatellite primers
1.5 PCR产物凝胶电泳检测
PCR产物电泳在8%的非变性聚丙烯酰胺凝胶上进行, 胶片规格为195 mm (长)×120 mm (宽)×1 mm (厚)。电泳条件: 电泳缓冲液为1×TBE, 电压200 V, 电泳1.5—2h。产物上样量均为1 μL, DNA Marker (50 bp DNA Ladder)上样量为0.5 μL。电泳完成后使用硝酸银染色, 参照高东等[15]的染色方法并稍加修改。最后将胶片置于灯箱, 进行拍照和保存。
2 结果
2.1 五个群体的SSR扩增结果
本研究筛选出多态性良好的16对微卫星引物,均能扩增出条带清晰、稳定的PCR产物, 且均表现为多态。并筛选出了引物TTF3、TTF4、TTF10以及Mam25, 4对可以区分5个群体的特异性引物。图 1为引物TTF3在5个群体中的扩增电泳图谱, 图 2为引物Mam25在5个群体中的扩增电泳图谱。通常利用多态信息含量、等位基因数和杂合度来衡量遗传标记的多态性[16]。衡量群体遗传变异通常用杂合度作为最合适参数[17]。利用这16对引物对这5个群体共150个样进行扩增分析, 团头鲂、三角鲂、翘嘴鲌、团头鲂♀×翘嘴鲌♂、三角鲂♀×翘嘴鲌♂5个群体分别扩增出57、58、55、64、69个等位基因, 每个位点检测到1—5个不等的等位基因数,平均等位基因数(Na)分别为3.56、3.63、3.44、4.00和4.31, 平均观测杂合度(Ho)分别为0.3510、0.3757、0.3175、0.3818和0.4079, 平均期望杂合度(He)分别为0.6182、0.6290、0.5921、0.6490和0.6825, 平均多态信息含量(PIC)分别为0.5354、0.5367、0.5258、0.5785和0.6067。具有较高的多态性(PIC>0.5000), 能够在分子层面上反应出各个群体间的遗传关系(表 2、表 3)。从这些遗传参数中可得出, 遗传多样性高低依次为三角鲂♀×翘嘴鲌♂>团头鲂♀×翘嘴鲌♂>三角鲂>团头鲂>翘嘴鲌,即三角鲂♀×翘嘴鲌♂的最高, 翘嘴鲌的最低。
2.2 遗传多样性分析
5个群体间的遗传距离和Nei’s遗传相似性(表4)利用Popgene (Version1.32)软件计算, 5个群体间的Nei’s遗传相似系数为0.4207—0.9183, 遗传距离为0.0853—0.5204。其中团头鲂和三角鲂相似性系数最高, 为0.9183; 团头鲂和翘嘴鲌相似性系数最低, 为0.4207。图 3为5个群体间的UPGMA聚类关系, 利用MEGA5.1根据5个群体间Nei’s遗传距离所构建, 结果显示团头鲂与三角鲂首先聚类; 团头鲂♀×翘嘴鲌♂与三角鲂♀×翘嘴鲌♂首先聚类, 然后两大类聚为一支, 最后与翘嘴鲌聚为一支。其中, 团头鲂和三角鲂遗传距离最近, 为0.0853; 团头鲂与翘嘴鲌遗传距离最远, 为0.5204。
3 讨论
通过部分不同鱼类属间或种间的亲本杂交, 可得到具有亲本优良性状的杂种子代[1]。据目前统计, 有关团头鲂、三角鲂以及翘嘴鲌的杂交组合主要有团头鲂(Megalobrama amblycephala)(♀)×鳜(Siniperca chuatsi)(♂)[18]、鳙(Aristichthy nobilis)(♀)×团头鲂(♂)及其反交[19]、草鱼(Ctenopharyngodon idellus)(♀)×团头鲂(♂)[20, 21]、草鱼(♀)×三角鲂(M. terminalis)(♂)[22, 23]、鲢(Hypophthalmichthys molitrix)(♀)×团头鲂(♂)及其反交[24, 25]、鲤(Cyprinus carpio)(♀)×团头鲂(♂)[26—28]、长春鳊(Parabramis pekinensis)(♀)×三角鲂(♂)[6]、团头鲂(♀)×长春鳊(♂)[29]、红鳍鲌(Culter erythropterus)(♀)×团头鲂(♂)[30]、广东鲂(Megalobrama hoffmanni)(♀)×团头鲂(♂)[31, 32]、三角鲂(♀)×团头鲂(♂)及其反交[33]、海南红鲌(E. pseudobrevicauda)(♂)×翘嘴红鲌(♀)[34]、翘嘴鲌(♀)×团头鲂(♂)[7]、团头鲂 (♀)×黄尾密鲴(Xenocypris daviodi)(♂)[35]、团头鲂(♀)×丁鱥(Tinca tinca)(♂)[36]及其反交等。这些杂交组合均可受精并获得鱼苗, 但子代的可育性以及成活率有所差异。总之, 鱼类有着相对复杂遗传结构, 很难全部了解, 因此, 鱼类的远缘杂交具有良好的发展前景, 可以利用更多的鱼类进行远缘杂交, 获得更多具有杂种优势的良种。

图 1 TTF3在5个群体中的PAGE图谱Fig. 1 PAGE analysis by primers TTF3 in 5groups

图 2 Mam25在5个群体中的PAGE图谱Fig. 2 PAGE analysis by primers Mam25 in 5 groups
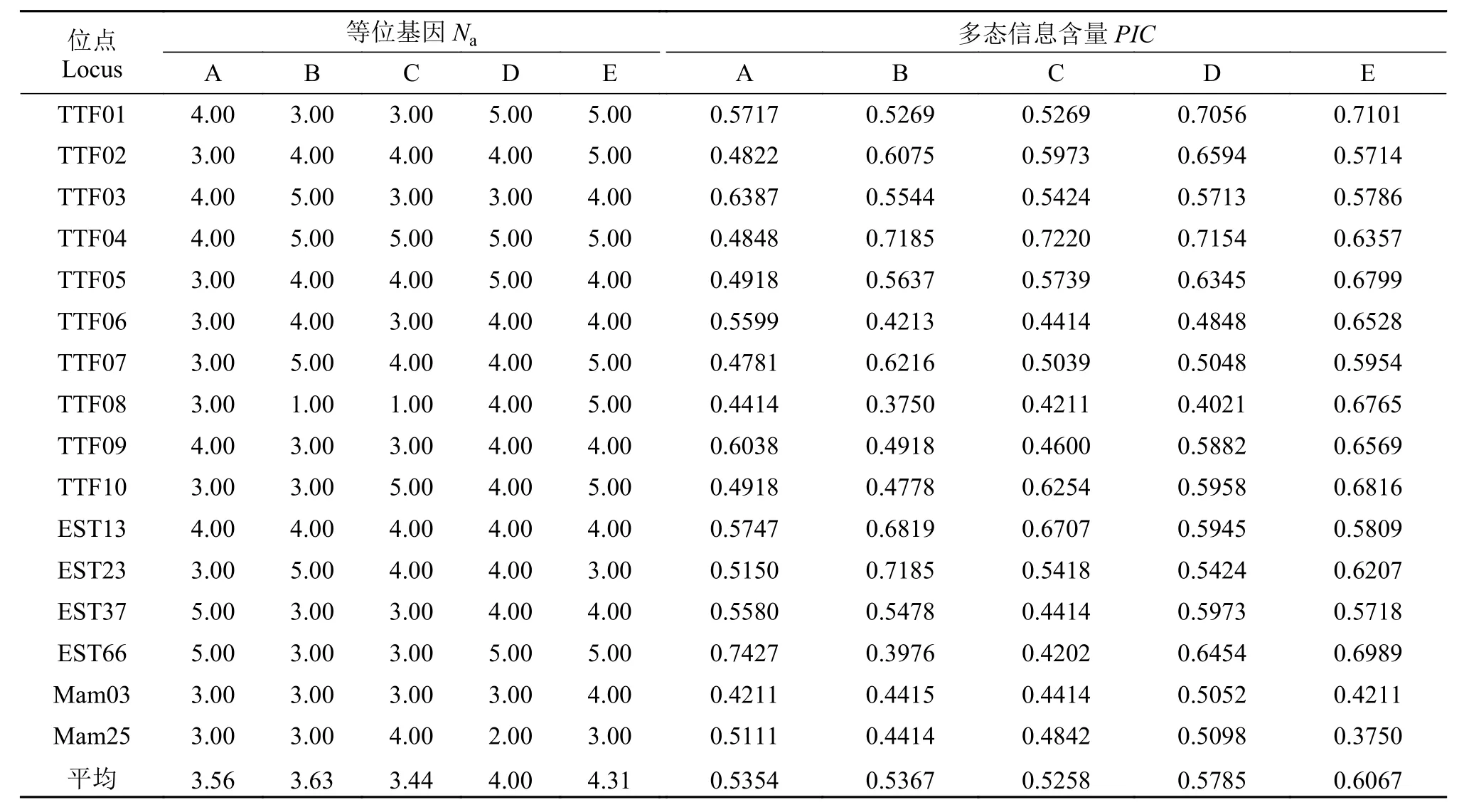
表 2 五个群体的微卫星位点的等位基因数、多态信息含量Tab. 2 Number of alleles (Na) and the polymorphism information content (PIC) of microsatellite loci
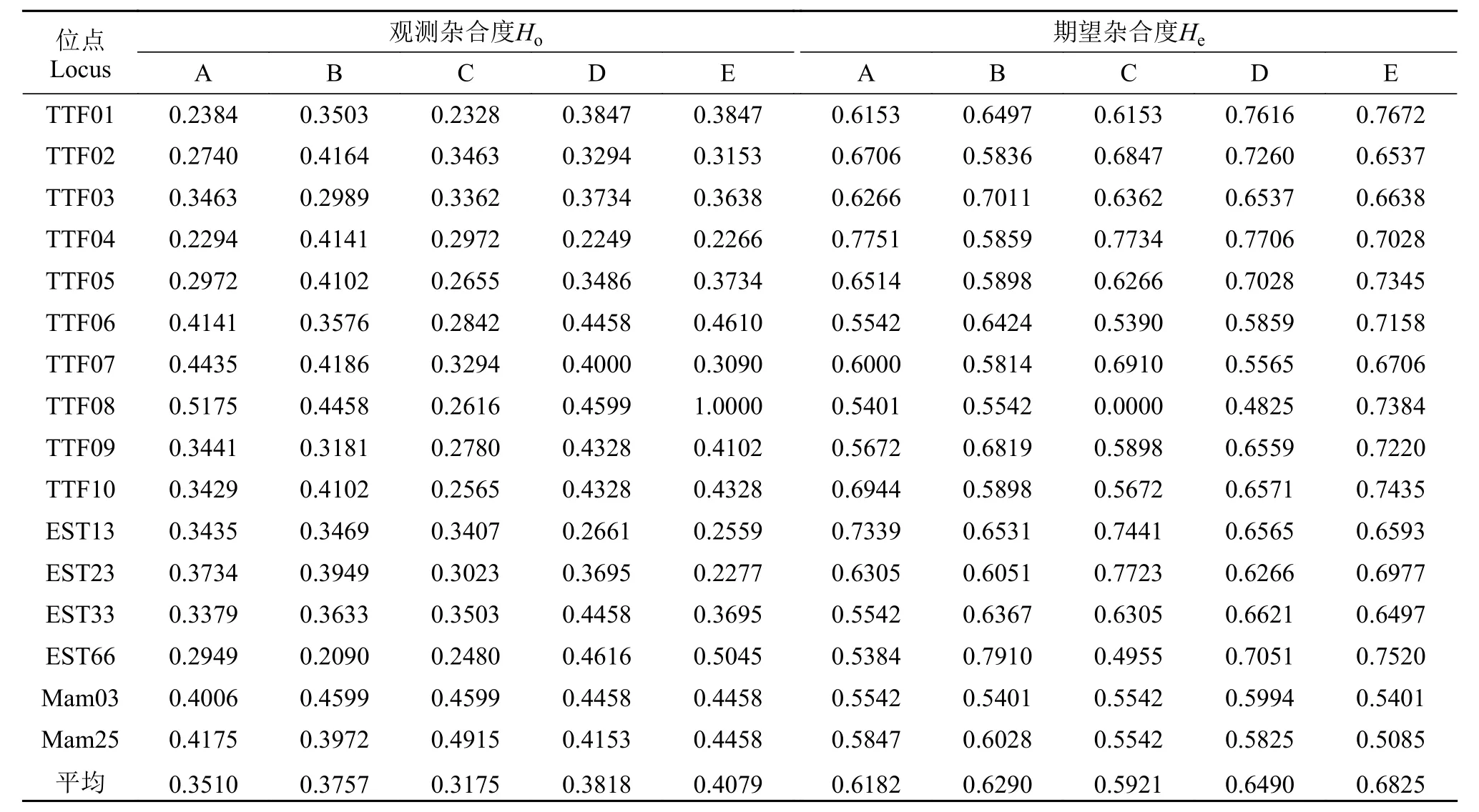
表 3 五个群体的微卫星位点的观测杂合度、期望杂合度Tab. 3 Observed heterozygosity (Ho) and expected heterozygosity (He) of microsatellite loci

表 4 不同群体的Nei’s遗传相似性(对角线上方)和遗传距离(对角线下方)Tab. 4 Nei’s genetic identity (above diagonal) and genetic distance(below diagonal)

图 3 不同群体的UPGMA聚类图Fig. 3 Dendrogram of among various populations using UPGMA clustering method
本研究表明, 遗传多样性和遗传潜力可通过期望杂合度(He)、观测杂合度(Ho)、等位基因的数目(Na)和多态性信息含量(PIC)等遗传参数反映出来[37]。李思发等[38]研究表明杂交种表现性状会产生杂交优势, 杂交鱼具有比双亲更为丰富的多态性。不同标记数和样本数量对遗传参数的影响很大[39], 本研究5个群体在相同样本数量的条件下, 平均等位基因数(Na)分别为3.56、3.63、3.44、4.00和4.31, 平均观测杂合度(Ho)分别为0.3510、0.3757、0.3175、0.3818和0.4079, 平均期望杂合度(He)分别为0.6182、0.6290、0.5921、0.6490和0.6825, 平均多态信息含量(PIC)分别为0.5354、0.5367、0.5258、0.5785和0.6067。杂交群体的平均多态信息含量均大它们的亲本群体, 这可能是由于杂交子代整合了亲本的基因组。杂交群体平均多态信息含量(PIC)>0.5000。在通常情况下,PIC>0.5000为高度多态, 0.2500<PIC<0.5000为中度多态,PIC<0.2500为低度多态[40]。综合各个遗传参数可以得出, 杂交群体具有较高的遗传多样性。
利用UPGMA法对5个群体进行聚类分析, 结果显示团头鲂与三角鲂首先聚类, 团头鲂♀×翘嘴鲌♂与三角鲂♀×翘嘴鲌♂首先聚为一支, 这2大支聚类后又与翘嘴鲌聚类。通过聚类分析可以看出团头鲂与三角鲂遗传距离最近, 2种杂交子代遗传距离最近, 而翘嘴鲌与其他群体遗传距离较远。团头鲂和三角鲂属鲂属, 而翘嘴鲌属鲌属, 属内遗传距离要近于属间, 这一点与结果相符。2种杂交子代聚类后先与母本聚类后的一支聚类, 这是因为远缘杂交获得的子代都会有一定的形态差异[33, 34, 41], 双亲提供的遗传物质不相等。团头鲂♀×翘嘴鲌与母本团头鲂的遗传相似系数为0.7653, 与父本翘嘴鲌的遗传相似系数为0.6566; 三角鲂♀×翘嘴鲌♂与母本三角鲂的遗传相似系数为0.7224, 与父本翘嘴鲌的遗传相似系数为0.6213。2种杂交鱼的在遗传上都有些偏向母本, 但亲本偏向性都不算很大, 整合了亲本的优良基因[42], 但继承母本的遗传物质较多一些, 具有母本效应[38], 这为定向育种提供了一定帮助。
孟德尔分离定律指出在生物的体细胞中, 控制同一性状的遗传因子成对存在, 不相融合; 在形成配子时, 成对的遗传因子发生分离, 分离后的遗传因子分别进入不同的配子中, 随配子遗传给后代。通过引物TTF3在5个群体中的扩增电泳图谱以及引物Mam25在5个群体中的扩增电泳图谱可以看出, 杂交子代均可扩增出清晰条带。通过对杂交子代和亲本的基因型分析, 杂交子代的等位基因均来源于亲本, 杂交子代通过杂交组合产生了新的基因型, 符合孟德尔分离定律。
鱼类种质资源的研究对生物多样性的保护具有重要意义, 实验筛选出具有特异性的TTF3、TTF4、TTF10以及Mam25四对引物, 对于群体的鉴定、种质资源保护和有效的开发利用提供了一定的参考。目前, 有研究表明鳊鲂杂种较双亲有明显生长优势[6], 本实验数据显示三角鲂×翘嘴鲌和团头鲂×翘嘴鲌的杂交群体具有较高的遗传多样性, 可以使鱼类遗传育种的基因库得到一定的拓展, 使原种的有利基因得到充分利用, 对于原种来说具有明显杂交优势。本实验对三角鲂×翘嘴鲌和团头鲂×翘嘴鲌的杂交育种实践应用提供了理论依据。
