我国南亚热带森林群落先锋树种马尾松的水分利用特征
2019-03-06王立景邓永红曾小平刘世忠李跃林
王立景 ,邓永红 ,曾小平 ,刘世忠 ,孟 泽 ,李跃林
(1.中国科学院 华南植物园,广东 广州 510650;2.中国科学院生态环境研究中心,北京 100085;3.中国科学院大学,北京 100049)
蒸腾是植物将水分释放进入大气的重要的生理过程,影响着全球的水循环过程[1-2]。目前对于蒸腾的监测已有较多的研究方法[3-5],利用树干液流可精确的测量树木蒸腾[6],其中,热消散探针法以其结果精确以及成本较低等一系列原因得到广泛应用[7-8]。已有的研究发现,树干液流的日变化均为昼高夜低的变化趋势[9-12],且夜间树干液流速率不为零;树干液流速率受光合有效辐射辐射、水汽压亏缺、气温、湿度、风速和土壤含水量等环境因子深刻影响[13-17],其中光合有效辐射和水汽压亏缺是主要的影响因子[18-19]。
马尾松作为造林先锋树种,已经在中国南方生态恢复初期被广泛种植[19-20],针对该树种的研究目前也比较多,从水涵养功能[20]、碳储量[21-23]、土壤养分动态[24-25]到其繁育过程[26-27]都有大量研究,但是对马尾松水分利用特征的研究很少报道,且已有研究多集中在群落演替初期[19-20]。
在森林群落不同演替阶段,林木表现出不同的水分利用特征[28],这可能是导致其在不同演替阶段的森林群落中生态位差异的关键因素。以我国鼎湖山南亚热带森林群落演替中期的针阔叶混交林为研究对象,运用树干液流方法,对该林型中马尾松水分利用特征进行研究。阐述马尾松在演替中期的水分利用特征,对比分析马尾松在森林演替过程中的水分生理特征的变化,为我国南亚热带森林生态系统科学经营和管理提供科学依据。
1 材料与方法
1.1 样地概况
试验地位于鼎湖山国家级自然保护区的联合国教科文组织人与生物圈的定位研究站(112°30′39″~ 112°33′41″E, 23°09′21″~23°11′30″N),属南亚热带季风气候,年均气温21℃,年降水量1 956 mm,干湿季较明显,全年降水量的80%集中在湿季的4—9月,年均相对湿度80.8%。保护区面积约1 133 hm2,分布着多种森林类型。本文研究的森林群落为演替中期的针阔叶混交林,是广东省马尾松针叶林群落向针阔叶林群落演替的中间森林类型,在本地具有典型代表性。该群落在垂直结构上大致分为3层,乔、灌、草,此外还有少量藤本和附生等层间植物。组成种类以常绿树种占绝对优势,其中优势种有锥栗Castanopsis chinensis、木荷Schima superba、马尾松P.massoniana、黄果厚壳桂Cryptocarya concinna等,马尾松为优势种之一。
为研究马尾松在针阔叶林阶段的水分利用特征,于2010年选取代表性的林段600 m2建立样地,样地位于鼎湖山森林生态系统定位研究站客座公寓后面,坡度6°~8°,坡向南,海拔30~40 m,母岩为砂页岩,土壤为赤红壤。林木密度为25株/100 m2,林龄60~80 a,叶面积指数4.02,其中马尾松密度为2株/100 m2,马尾松胸径(DBH)最大为28.4 cm,最小为5.4 cm,平均DBH为23.12 cm,平均树高为16.64 m。
1.2 研究方法
1.2.1 边材面积的测定
在针阔叶混交林选取30 m×40 m样地,根据样地实际情况,选取3株树干圆满通直、生长状况良好且无病虫害的马尾松作为研究对象,样树的基本特征如表1所示。
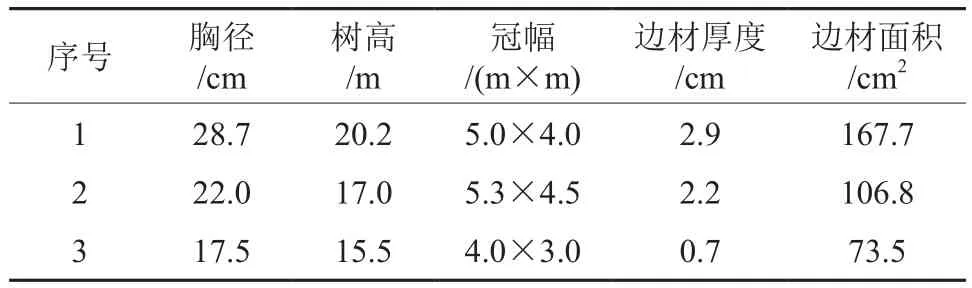
表1 样树基本特征Table 1 Basic characteristics of the sampling trees
在实际操作中,只能通过测定边材厚度求算边材面积。出于对自然保护区植被保护的原则,并避免对样树造成伤害,因此对研究地区采取抽样调查的办法(不包括待测树干液流的样树),选取8株,量取胸径,采用直径5 mm生长锥于树干北面胸高1.3 m处钻取木栓,考虑到林内郁闭度高,树干圆满通直,主要以北面木栓为主,量取胸高处边材厚度,计算边材面积并建立边材面积与胸径的关系式:

式(1)中:As代表边材面积(cm2),DBH为胸径,m、n为参数。
在同一地区所测马尾松边材面积与胸径关系为:y=0.84x2.29,R2=0.99[29]。
1.2.2 树干液流的测定
综合气象资料,采用Granier热扩散探针法于2010年7月至2012年6月对3株样树的树干液流速率进行连续测定。将一对20 mm探针上下相距10~15 cm安装于树干北面(避免阳光直射),胸高1.3 m处。为防止雨水接触探针,导致测定产生误差,在探针外覆盖泡沫盒,并包裹防辐射薄膜,减小温度波动。上探针供以12 V直流电压持续加热(0.2 W),下探针作为参照不加热。两探针之间的温差电势应用数据采集器DL2e(Delta-T Devices,England)自动记录和存储(每10 s测读1次数据,存储每30 min的平均值)。根据Granier[7]建立的经验公式将温差电势转化为树干液流速率:
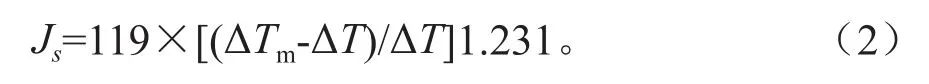
式中:ΔTm为上、下探针之间的最大昼夜温差(℃),ΔT为瞬时温差(℃),Js为瞬时树干液流速率(g·m-2s-1)。
1.2.3 整树蒸腾速率的计算
将树干液流速率测定值扩展到整树蒸腾的尺度水平,马尾松平均蒸腾速率Et(g·s-1Tree-1)由如下公式计算[30]:

式中:Js为瞬时树干液流速率(g·m-2s-1),As为样树边材面积(m2)。夜间树干液流速率主要用于补充贮水部位的水分亏缺,因此不计入整树蒸腾量估算。
1.2.4 气象因子的测定
微型气象观测仪安装于样地内高约10 m处的空旷地铁架上,以避免树木等障碍物影响数据准确性。无线电子测量记录器(Derelektronische Funk-Messlogger funky_Clima,德国)用于测量气温(T)和空气相对湿度(RH)。Li-cor光合有效辐射传感器(LI-COR Environmental,美国)用于对光合有效辐射进行连续监测。土壤10 cm和30 cm处安插Em50-ECH2O型土壤含水量传感器,环境因子数据采集和存储频率均与树干液流相同。为综合表达温度和空气相对湿度的协同效应采用VPD(vapor pressure deficiency)这一指标,采用下式计算VPD[31]:

式中:es(T)代表T温度下的饱和水汽压(kPa),a、b、c为参数,分别取值为:0.611 kPa、17.502、240.97 ℃,T为温度(℃),ea为实际水汽压(kPa),RH为相对湿度,VPD为叶片和空气之间的水汽压亏缺(kPa)。
1.2.5 数据处理
本文运用Excel 2016软件进行数据处理,用SPSS19.0统计软件中的Pearson偏相关分析对树干液流与环境因子进行偏相关分析,并进行回归分析,运用Sigmaplot12.5绘图。
2 结果与分析
2.1 微气象环境
华南地区每年4月至9月为湿季,10月至翌年3月为干季。该地区微气象因子特征如图1所示,2010年内的PAR与2011年和2012年同期的结果相近(P>0.05),湿季PAR均显著高于干季(P<0.001);T、RH和VPD的结果与PAR相同,2010—2012年同期的结果相近(P>0.05),三者在湿季的结果均显著高于干季(P<0.001)。2010年7月—2011年6月间,日均PAR最大值为 814.582 μmol·m-2s-1(8月3日),日均VPD最大值为2.446 kPa(5月9日),日均T最大值为32.84℃(7月1日)。
2.2 马尾松水分利用特征
温度、水汽压亏缺和光合有效辐射的日变化表现为昼高夜低的单峰曲线,相对湿度的日变化表现为昼低夜高的趋势,两者均为湿季大于干季。马尾松树干液流速率的日变化趋势也表现为昼高夜低,白天湿季树干液流速率大于干季,夜间湿季树干液流速率小于干季(图2)。华南地区植物树干液流的存在时间(12个月)长于北方地区(4—10月)[15];湿季树干液流速率的平均值高于干季,且峰值范围要宽于干季;白天胸径最大的树干液并未最高,胸径最小树干液流速率最快,湿季峰值为25.24 g·m-2s-1(图2左下图版),干季峰值最高达17.75 g·m-2s-1(图2右下图版);夜间马尾松有较低的树干液流速率,且变幅较小,基本稳定。
2.2.1 马尾松树干液流的变化与环境因子的关系
将树干液流速率与光合有效辐射、水汽压亏缺、温度、空气湿度的最大值进行边界线分析,如图3所示。随着PAR升高,树干液流速率升高,当 PAR 超过 1500 μmol·m-2s-1,树干液流速率在24.16 g·m-2s-1上下呈稳定波动状态,这可能是因为马尾松达到光饱和值,蒸腾达到相对稳定状态。随着VPD增大,树干液流速率呈增长状态,当VPD超过2.6 kPa时,树干液流速率从20.91 g·m-2s-1下降,这是因为植物在应对过高的VPD时,会关闭气孔以阻止过多水分流失。随着温度升高,树干液流速率整体呈增长趋势,低于14℃时,树干液流速率增长缓慢,在14~24 ℃时,树干液流速率在3.11 g·m-2s-1呈稳定波动趋势,当温度超过24 ℃,树干液流速率快速增长。随相对湿度的增加,树干液流速率整体呈下降趋势。
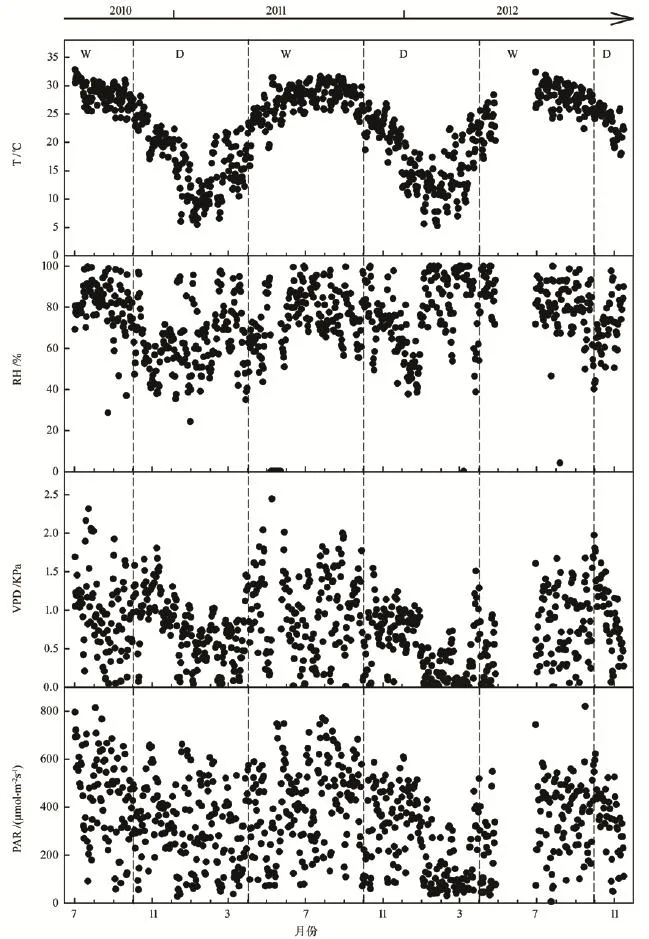
图1 2010年7月—2012年12月干湿季日平均温度、相对湿度、水汽压亏缺与光合有效辐射Fig.1 Daily mean air temperature (T), relative humidity(RH), vapor pressure deficit (VPD) and photosynthetic active radiation (PAR) from July 2010 to December 2012 (D:dry seasons; W:wet seasons)
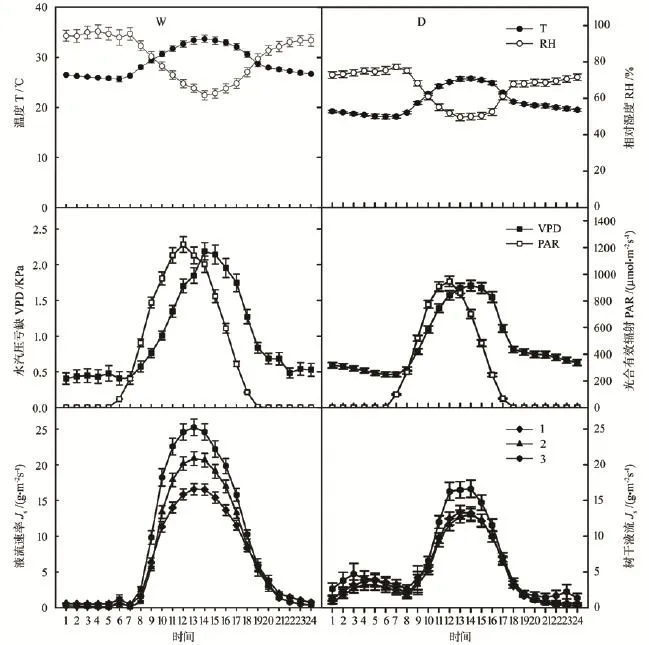
图2 在湿季(7月)和干季(11月)环境因子和马尾松树干液流速率的日变化Fig.2 Diurnal courses of air temperature, relative humidity, vapour pressure deficit, photosynthetically active radiation and sap flow velocity (D:dry seasons; W:wet seasons)
将月尺度上马尾松树干液流速率与同步监测的环境因子进行偏相关分析,结果如表2所示。从偏相关系数来看,在湿季(4—9月),马尾松树干液流速率主要与PAR呈极显著正相关,其中7月份(最热月)主要与T成极显著正相关;在干季(10月—翌年3月),树干液流速率主要与VPD呈极显著正相关,其中10月和11月树干液流速率主要与PAR呈极显著正相关。结果表明PAR与VPD为控制树干液流速率的主要环境因子。
通过错位对比法获得树干液流速率对PAR和VPD的时滞时间,如表3所示。结合图2进行分析发现,在湿季(7月),树干液流发生和峰值时间分别为7:00和13:00,滞后于光合有效辐射(6:00和12:00)1~2 h,提前于水汽压亏缺(5:00至6:00)1 h;在干季(11月),树干液流发生和峰值时间分别为8:00和14:00,滞后于光合有效辐射(7:00和12:00)2 h,提前于水汽压亏缺(7:00和14:00)小于1 h。

表2 年尺度上树干液流与PAR、VPD、RH、T间的偏相关分析†Table 2 Partial correlation analysis between sap flow and PAR, VPD, RH, T on a monthly scale

图3 日尺度上, 树干液流速率最大值对光合有效辐射、水汽压亏缺、温度、空气湿度的响应规律Fig.3 On a daily scale, the response laws of maximum sap flow rate to photosynthetic active radiation,water vapor pressure deficit, temperature and air humidity
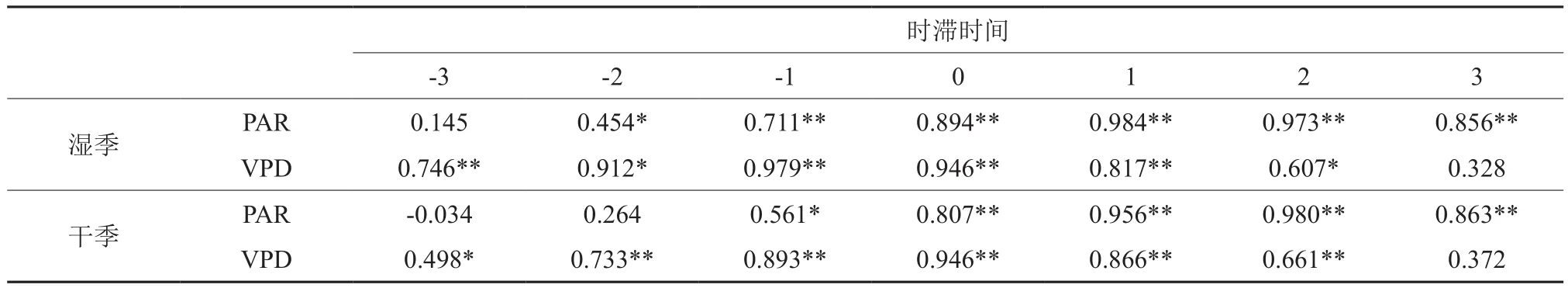
表3 湿季和干季马尾松树干液流与不同环境因子时滞相关系数†Table 3 Time delay correlation coefficients between sap flow velocity and environmental factors in wet season (Jul.)and dry season (Nov.)
2.2.2 马尾松蒸腾耗水量
马尾松在湿季、干季和年际的日均蒸腾耗水量如图4所示,样树的树干液流速率按胸径大小排序,在湿季分别为 22.9、16.2、10.3 kg·d-1,在干季分别为15.1、8.7、5.3 kg·d-1,大胸径个体蒸腾量高(P<0.001),湿季的蒸腾量显著高于干季(P<0.001)。
3 结论与讨论
3.1 结 论
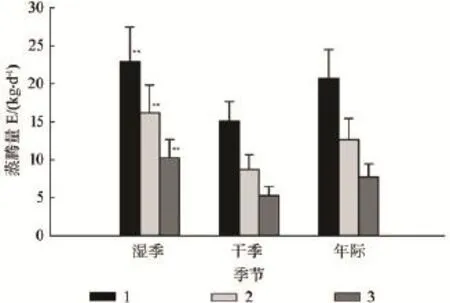
图4 马尾松干湿季和年日均蒸腾耗水量Fig.4 Average daily transpiration water consumption of P.massoniana during dry season, wet seasons and the whole year
1)鼎湖山地区马尾松全年均存在树干液流速率,其日变化呈昼高夜低的单峰型曲线,中午树干液流速率最高,湿季的峰值高于干季,夜间也存在微量树干液流,且基本稳定;
2)马尾松树干液流速率主要受光合有效辐射和水汽压亏缺的影响,湿季光合有效辐射起主要作用,干季水汽压亏缺起主要作用;在干湿两季,树干液流速率滞后于光合有效辐射,提前于水汽压亏缺;
3)马尾松在湿季的蒸腾量显著高于干季;与森林恢复初期的马尾松树干液流相比,本地区树干液流速率相对较低,表明随着演替的进行,马尾松树干液流速率下降。蒸腾耗水量的降低,可能是华南地区马尾松在演替后期被取代的重要原因。
3.2 讨 论
目前有较多研究发现,植物单日的树干液流速率呈现出昼高夜低的双峰或多峰曲线,如马占相思[11]、花旗松[32]、木荷[33],本研究中马尾松呈单峰型曲线,这主要是鼎湖山地区湿度相对较高,温度比城市略低,中午温度最高时未使气孔关闭。夜间,树干液流速率很微弱,但是都不为0,且干季变化幅度比湿季大,这与Resco[34]和赵晓伟等[35]的研究结果相似。
马尾松的树干液流速率变化主要受光合有效辐射和水汽压亏缺影响,这与已有的研究结果相似[11,36-37],已有的研究表明:不同月份,树干液流速率与不同环境因子呈极显著相关,卢志朋等[38]发现樟子松树干液流速率受土壤湿度的影响随月份不同而不同,各月水汽压亏缺与树干液流速率相关系数较大;Patankar等[39]发现在夏季(6—7月)黑云杉树干液流速率与水汽压亏缺和温度的相关性强;程静等[29]发现,马尾松7月份受水汽压亏缺的影响略大于光合有效辐射,与土壤含水量正相关关系不显著,11月份受光合有效辐射的影响大于水汽压亏缺,且与土壤含水量呈极显著负相关关系。本研究发现马尾松树干液流速率在湿季主要由光合有效辐射控制,在干季主要由水汽压亏缺控制。
不论在湿季还是干季,马尾松树干液流速率均滞后于PAR而提前于VPD,且不同季节的时滞和提前时间不同,在湿季,树干液流滞后于光合有效辐射1~2 h,提前预水汽压亏缺1 h;在干季,则滞后于光合有效辐射2 h,提前于水汽压亏缺小于1 h。与其他研究结果相似,Ewers[40]等发现火炬松树干液流速率与VPD间的时滞达30 min;王华等[41]发现马占相思在湿季和干季对PAR的滞后时间为60和140 min,对VPD的提前时间为80和50 min;赵春彦等[8]发现胡杨在6月对PAR时滞时间小于1 h,在5、7、8、9月时滞约1 h,5、6、8月对VPD提前1 h,7月提前3 h,9月提前时间小于1 h。
在演替初期,马尾松树干液流速率的变化趋势基本相同,郑怀舟等[18-19]研究表明演替早期的马尾松单日树干液流速率呈单峰型曲线,凯旋等[42]研究表明重庆近郊马尾松经营林分(演替早期)的树干液流速率为单峰型,本研究中演替中期的马尾松单日树干液流速率呈单峰型曲线。树干液流速率的峰值和日均值分别为17.56和5.64 g·m-2s-1,低于郑怀舟等所得出的24.06、20.60 g·m-2s-1和 7.08、6.11 g·m-2s-1, 鲁 小珍[43]所得出的峰值22.06 g·m-2s-1。本研究湿季日均蒸腾量10.3~22.9 kg·d-1,干季为5.3~15.1 kg·d-1,同时期凯旋所得马尾松日均蒸腾量为53.224 kg·d-1和 26.136 kg·d-1,对比分析表明,从演替初期到中期,马尾松树干液流速率下降,这主要与演替不同阶段物种多样性造成的竞争及林内微环境因子的变化有关。
陈彪等[44]对樟子松的研究表明,不同胸径樟子松单株耗水量差别较大,除了大的树干液流速率会导致较高的蒸腾量外,树木具有较大的边材面积也会导致其耗水量较多;赵平等[11]对马占相思的研究表明,胸径稍小的树木具有较高的单位基面积日树干液流量(kg·dm-2d-1),胸径越大树木整树蒸腾越高(P<0.001)。本研究中胸径最大的马尾松树干液流速率不是最高,但是其边材面积大,导致其蒸腾量大,按样树胸径大小排序,样树年日均蒸腾量顺序为20.7、12.6、7.7 kg·d-1。本结果是对马尾松作为先锋树种在演替中期的针阔叶林阶段的水分传输能力的解释,联系全球变化情形下我国南亚热带水热格局的变化,将有利于定量我国南亚热带华南地区森林生态系统的水分过程,为研究森林生态系统演替的水分过程及评估生态系统服务功能提供了理论依据。
本研究集中探讨了月尺度上环境因子对马尾松树干液流速率的影响。有研究表明,年尺度上,土壤含水量(SWC)是树干液流速率主要的影响因子。而在全球变化条件下,位于我国南亚热带的鼎湖山降水量虽然没有显著改变,但是土壤含水量显著减小,全年无雨日增加,暴雨的频率增加[45],这种变化趋势必然对先锋种马尾松的水分活动产生影响,其影响结果可做深入的探讨。
