骨髓间充质干细胞旁分泌HGF体外调控肝星状细胞
2019-02-14李静郑雪丁新向辉陈卫刚
李静,郑雪,丁新,向辉,陈卫刚
肝脏疾病是我国常见病、多发病,肝硬化和肝癌是肝脏疾病重要的致死原因。肝纤维化是肝硬化和肝癌的中间环节,而抑制肝星状细胞的激活和增殖是缓解肝纤维化的策略之一。骨髓间充质干细胞(BMSCs)是骨髓中的非造血组织,因具有自我更新、多向分化、免疫抑制等功能而成为临床研究热点[1]。现越来越多的动物实验及临床试验表明BMSCs可治疗骨或软骨损伤、心脏疾病、中枢神经系统损伤、脊髓损伤[2]。近年来,BMSCs在肝纤维化方面的治疗也取得了一定的进展[3],但其具体机制还不明确。本实验将骨髓间充质干细胞与肝星状细胞(HSCs)建立间接共培养体系,初步探讨骨髓间充质干细胞缓解肝纤维化的机制。
1 材料与方法
1.1 材料 清洁级SD雄性大鼠,体质量120~180 g,购自新疆维吾尔自治区疾病预防控制中心;大鼠肝星状细胞(上海富衡生物科技有限公司);胎牛血清(美国BI公司)、L-DMEM培养基(美国Gibco公司),0.25%胰蛋白酶(美国Hyclone公司),CD29-FITC抗体、CD45-FITC抗体、CD90-PE抗体(eBioscience公司);MTT(Solarbio公司);肝细胞生长因子ELISA试剂盒(中国Elabscience公司);c-met抗体(MCE公司);兔抗α-肌动蛋白(α-SMA)单克隆抗体(ab32575,Abcam公司);荧光二抗(山羊抗兔IgG-FITC,北京中杉金桥生物科技有限公司);PI(美国Sigma公司);Transwell insert半透膜(0.4µm,美国Corning Costar公司);Annexin V-PE细胞凋亡试剂盒(中国联科生物公司);流式细胞仪(美国BD公司);酶标仪(美国BIO-RAD公司);激光共聚焦显微镜(德国ZEISS公司)。
1.2 方法
1.2.1 细胞的提取、培养和鉴定 利用全骨髓贴壁培养法提取大鼠双下肢骨髓间充质干细胞,收集第2代对数生长期细胞,流式鉴定表面分子CD29、CD90、CD45;复苏新购的HSCs,取第3代生长活跃的细胞,倒置相差显微镜下观察活体细胞形态学改变,采用荧光免疫法检测α-SMA的表达。
1.2.2 细胞共培养体系的建立及实验分组 6孔板半透膜(直径0.4µm)上层每孔接种1×105个细胞,下层接种5×104个细胞/孔。实验分组:(1)H组,HSCs单独培养(半透膜上层只有培养基)。(2)H-H组,HSCs和HSCs共培养。(3)M-H组,BMSCs与HSCs共培养。(4)M-H-C组,c-met抑制剂2 mmol/L预先封闭HSCs表面肝细胞生长因子(HGF)受体,4 h后BMSCs与HSCs共培养;48 h后倒置相差显微镜下动态观察活体细胞形态学改变。
1.2.3 MTT法检测HSCs增殖抑制率 各组细胞共培养48 h后,抽取上清液保留备用,0.25%胰酶消化贴壁细胞,用各组共培养上清液分别重悬各组细胞,以3 000个细胞/孔均匀接种在96孔板上,孵育4 h,向各孔加10µL MTT液,4 h后弃上清,各孔加100µL二甲基亚砜(DMSO),在酶标仪上测定各孔在450 nm处的光密度(OD)值;细胞生长抑制率=(H组平均OD值-各处理组平均OD值)/H组平均OD值×100%。
1.2.4 共聚焦显微镜检测HSCs中α-SMA的表达 各组细胞共培养48 h后,弃上清,4%多聚甲醛固定细胞15 min,0.2%Triton-X-100透膜3 min,5%BSA封闭,孵α-SMA抗体,4℃过夜,次日暗室加荧光二抗,37℃孵育1~2 h,PI染核1 min,PBS洗4次,抗荧光淬灭剂封片,即刻采用激光共聚焦显微镜观察。
1.2.5 流式细胞仪检测HSCs凋亡率 各组细胞共培养48 h后,用不含EDTA的胰酶消化细胞,收集各组HSCs并计数,按照Annexin V-PE/7-ADD细胞凋亡试剂盒说明书操作,流式细胞仪检测细胞凋亡率。
1.2.6 ELISA检测共培养上清液HGF浓度 各孔中加入不同浓度的标准品或上下两层共培养上清液,按照肝细胞生长因子ELISA试剂盒说明书严格操作,用酶标仪在450 nm波长处测量各孔OD值。以浓度为横坐标,OD值为纵坐标,在双对数坐标纸上绘制四参数逻辑函数的标准曲线,根据标准曲线计算各组细胞上清液中HGF浓度。
1.3 统计学方法 应用SPSS 22.0软件对数据进行统计分析,所有计量资料以均数±标准差(±s)表示,组间比较采用单因素方差分析(one-way ANOVA),组间多重比较采用LSD-t法,P<0.05为差异有统计学意义。
2 结果
2.1 细胞形态特征及鉴定情况 原代BMSCs以集落为中心呈放射状向周围生长,培养至5~7 d时,细胞呈现明显的旋涡状。BMSCs流式鉴定结果显示,99.3%的细胞表达CD29,98.9%的细胞表达CD90,0.8%的细胞表达CD45,表明提取的原代细胞为纯化的BMSCs。HSCs在倒置显微镜下呈星形或多边形伸展状态,共聚焦结果显示活化后的HSCs胞膜及胞浆表达α-SMA,符合实验要求,见图1。
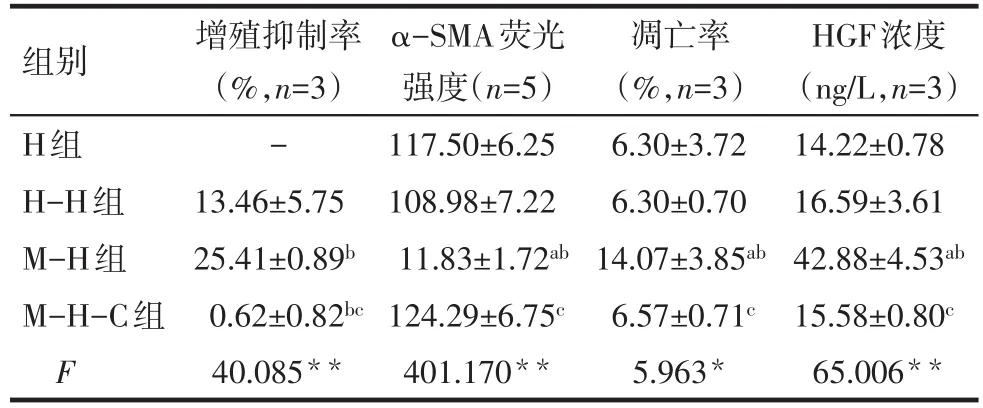
2.2 BMSCs抑制肝星状细胞的增殖 各组细胞共培养48 h后显示,BMSCs对HSCs的增殖抑制率最大,HSCs对HSCs的增殖抑制率次之,M-H-C组几乎无抑制,M-H组与H-H组、M-H-C组相比均有统计学意义(P<0.01),见表1。
Tab.1 Effects of BMSCs on HSCs after 48-hour co-culture表1 共培养48 h后BMSCs对HSCs的影响 (±s)
Tab.1 Effects of BMSCs on HSCs after 48-hour co-culture表1 共培养48 h后BMSCs对HSCs的影响 (±s)
*P<0.05,**P<0.01;a与H组比较,b与H-H组比较,c与M-H组比较,P<0.05
组别H组H-H组M-H组M-H-C组F增殖抑制率(%,n=3)-13.46±5.75 25.41±0.89b 0.62±0.82bc 40.085**α-SMA荧光强度(n=5)117.50±6.25 108.98±7.22 11.83±1.72ab 124.29±6.75c 401.170**凋亡率(%,n=3)6.30±3.72 6.30±0.70 14.07±3.85ab 6.57±0.71c 5.963*HGF浓度(ng/L,n=3)14.22±0.78 16.59±3.61 42.88±4.53ab 15.58±0.80c 65.006**
2.3 BMSCs抑制HSCs的活性 与M-H组比较,其余3组HSCs胞膜和胞浆α-SMA表达量均增高,差异有统计学意义(P<0.005),而其余3组间比较差异无统计学意义,见表1、图1。
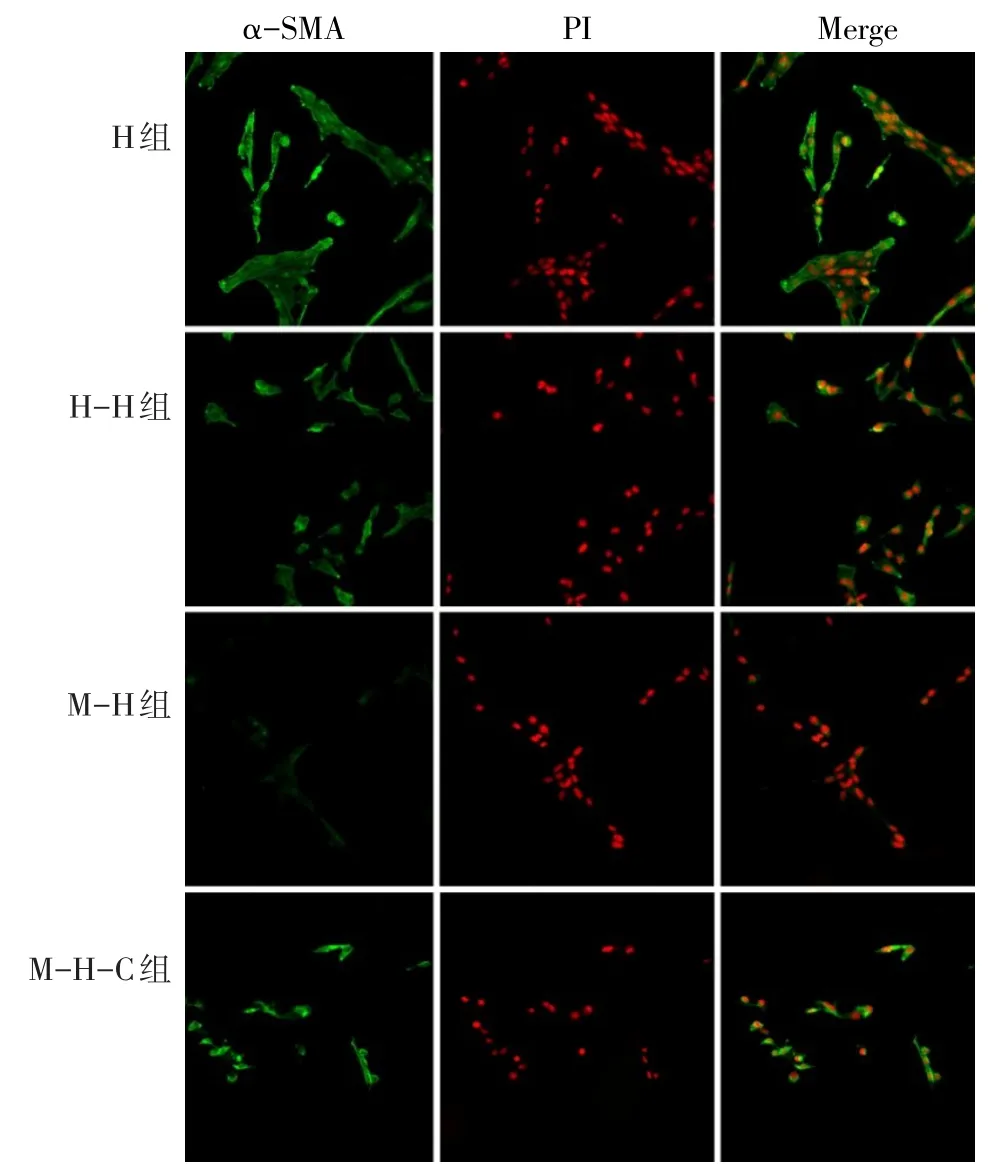
Fig.1 α-SMA expression of hepatic stellate cells in each group(×200)图1 各组肝星状细胞α-SMA表达量(×200)
2.4 BMSCs促进肝星状细胞的凋亡 流式结果显示,H组、H-H组、M-H-C组HSCs凋亡率较低,且3组差异无统计学意义,M-H组HSCs凋亡率明显高于H组、H-H组、M-H-C组,差异有统计学意义(均P<0.05),见图2、表1。
2.5 共培养上清液中HGF的浓度 ELISA结果显示,绘制的标准曲线与标准相符(R2=0.99),共培养48 h后,M-H组上清液HGF浓度明显高于H组、HH组、M-H-C组,差异均有统计学意义(均P<0.05),见表1。

Fig.2 The image of apoptosis in hepatic stellate cells图2 肝星状细胞凋亡图
3 讨论
目前肝硬化患者尚缺乏有效的治疗方法,干细胞因具有免疫原性低、易体外扩增、多向分化潜能,给肝硬化患者带来了新的希望。越来越多的研究表明干细胞移植治疗可以逆转肝纤维化和肝硬化,但具体机制尚不明确[4-5]。体内实验发现,BMSCs经尾静脉注射入四氯化碳(CCl4)诱导的肝纤维化大鼠体内,肝纤维化程度降低,HSCs的数量较对照组明显减少,凋亡率明显增高,这为BMSCs治疗肝纤维化的机制提供了新的思路[6]。HSCs经体外传代可被激活,活化HSCs细胞外基质的合成和降解失衡,导致细胞外基质增多,从而形成纤维化。
以往研究发现,间充质干细胞(MSCs)通过定向迁移到肝纤维化组织[7],分化为肝样细胞[8],并通过抗肝细胞凋亡改善肝功能[9-10],α-SMA是HSCs活化的标志,MSCs可以抑制成纤维细胞的增殖和α-SMA的表达,并促进基质金属蛋白酶的分泌[11]。Parekkadan等[12]在 BMSCs与HSCs共培养中发现,BMSCs来源的白细胞介素(IL)-10和肿瘤坏死因子(TNF)-α可抑制HSCs的增殖和胶原合成。本研究发现BMSCs与HSCs共培养48 h,BMSCs会明显抑制活化态HSCs的增殖并促进其凋亡,且作用和上清液中HGF的浓度有关,在HGF浓度最高的M-H组中,HSCs的增殖率最低,而凋亡率最高;当封闭HGF受体(c-met)后,增殖率升高同时凋亡率下降,说明BMSCs可能通过分泌的HGF抑制HSCs的增殖并促进其凋亡。韦柳萍等[13]检测了BMSCs与HSCs共培养24、48、72 h后HSCs的凋亡率,发现随着培养时间的延长,HSCs的凋亡率逐渐增加,且在72 h时达到最高。然而,本研究在前期预实验过程中发现,H组在培养72 h后,HSCs的凋亡率也明显增高,说明此时HSCs的高凋亡率并不是由BMSCs引起的,可能是培养体系中未更换新的培养基而无法满足HSCs的生长要求,亦或是HSCs分泌的因子引起的。Wang等[14]将BMSCs与LX2在transwell小室共培养后发现,LX2中α-SMA的表达量明显降低,当敲低c-met后,BMSCs不能改变活化LX2中α-SMA的表达量。本研究结果同样也显示,在BMSCs与HSCs共培养48 h后,BMSCs使活化态的HSCs活性降低,当封闭HGF/c-met信号通路后,效应消失,说明BMSCs可通过分泌的HGF抑制HSCs的活化。
MSCs可以旁分泌多种细胞因子,如HGF、IL-10、胰岛素样生长因子-1、神经生长因子、血管内皮生长因子、肿瘤坏死因子等。HGF是一种多效生长因子,在细胞的分裂、存活、迁移、形态发生等方面发挥着重要的作用。本研究在M-H组上清液中检测到高浓度的HGF,说明BMSCs可分泌HGF。而HSCs活化后可表达 HGF 受体 c-met[15],c-met受体属于酪氨酸激酶家族,HSCs通过分泌TNF-α、IL-6等因子刺激MSCs分泌HGF、IL-10等,HGF与HSCs表面的c-met受体结合后激活c-met位点,通过HGF/c-met信号通路对HSCs产生影响。本实验发现,在HGF浓度最高的M-H组中,HSCs的凋亡率最高,HSCs的增殖和活化最低,当封闭HGF/c-met信号通路后,效应消失,表明BMSCs对HSCs的影响可能是BMSCs分泌HGF通过HGF/c-met信号通路起作用的。然而,HGF/c-met下游有Akt/PKB、PI3K、JNK、ERK/MAPK、PIP3等通路,具体是哪条通路起主要作用还有待进一步研究。
此外,MSCs还可通过免疫调节和炎症抑制的作用,抑制T、B淋巴细胞的增殖,促进抑制性细胞因子分泌,减少炎症性细胞因子产生,从而减轻肝脏炎症反应和肝细胞的破坏[16-17]。随着外泌体的提出,越来越多的学者认为MSCs改善肝纤维化与外泌体有关[18-19]。Schorey等[20]认为外泌体为 MSCs与 HSCs间传递通讯的载体。Li等[21]认为来源于人脐带MSCs的外泌体通过抑制肝细胞的上皮-间充质转化、显著恢复血清天冬氨酸转氨酶活性,并通过降低Ⅰ/Ⅲ型胶原、转化生长因子-β1(TGF-β1)以及Smad2的磷酸化,使TGF-β1/Smad信号通路失活,从而改善肝纤维化。
综上所述,BMSCs可使HSCs的增殖、活化受到抑制,凋亡增加,其机制可能是BMSCs旁分泌HGF通过HGF/c-met信号通路起作用的,这也可能是BMSCs移植治疗肝纤维化的机制之一。
