比克氏棉色素腺体形态建成与棉酚代谢特性的研究
2019-01-31胡佳惠赵天伦李诚余静文陈进红祝水金
胡佳惠,赵天伦,李诚,余静文,陈进红,祝水金
(浙江大学农业与生物技术学院,浙江省作物种质资源重点实验室,杭州310058)
棉花是重要的经济作物,棉纤维是最主要的天然纤维来源。此外,棉花还为人们提供了大量优质的棉籽蛋白、棉籽饼和棉籽油[1]。由于棉籽中含有棉酚及其衍生物,对人和单胃动物有毒,限制了棉籽的使用[2-3]。但是,棉酚对棉花的自我保护起着重要作用[4]。因此,研究棉酚代谢有助于了解棉酚合成、转运、贮存特点,为提高植株棉酚含量和降低种子棉酚含量提供新的途径和方法。色素腺体是棉属及近缘植物特有的1种用来贮存棉酚及其衍生物的结构,通常呈褐色或黑褐色,遍布植株主要器官[5]。色素腺体是1个大的分泌空腔,外面围绕着1~3层的分泌细胞。前人研究表明,分泌细胞形成的空腔是细胞程序性死亡的结果[6]。澳大利亚的野生二倍体棉种,如澳洲棉(Gossypium australe)、比克氏棉(G.bickiiProkh.)和斯特提棉(G.sturtianum)等具有子叶色素腺体延缓形成性状,即休眠种子中无肉眼可见的色素腺体,检测不到棉酚;种子萌发后的子叶和下胚轴会陆续形成色素腺体,并检测到棉酚,且植株的根、茎、叶等主要器官均可观察到色素腺体[7-8]。这一特殊性状对于培育种子无色素腺体而植株有色素腺体棉花具有十分重要的价值。
Punit等[9-10]发现基因型在很大程度上影响色素腺体分布,且棉酚含量与色素腺体的数目有很强的相关性,因此常把二者一起研究。丁亮等[11]对种仁有色素腺体无酚型品种司笃克氏棉进行色素腺体形成与棉酚合成的动态研究。结果表明,在种胚发育过程中,棉酚的含量随发育进行呈现上升趋势,认为种胚发育过程中棉酚的含量与色素腺体的形成具有一定的关系。而在种子萌发过程中,其色素腺体密度和棉酚的含量呈负相关。目前,棉酚合成通路已明确[12],但并不清楚棉酚合成的关键酶是否参与色素腺体形态建成。马丹等[13]沉默了棉酚生物合成途径中的关键基因,其棉酚含量显著减少,但仍可形成正常的色素腺体。因此,棉酚与色素腺体之间的关系错综复杂,需要更多的证据加以澄清。
本研究以子叶色素腺体延缓发生的比克氏棉为材料,以陆地棉(G.hirsutum)有色素腺体品种标准系TM-1、陆地棉有色素腺体品种中棉所12及其无色素腺体近等基因系中棉所12无为对照,通过观察比克氏棉在种胚形成期和种子萌发期色素腺体形态建成,色素腺体形态与棉酚含量的关系,分析色素腺体形成与棉酚代谢的动态关系。通过分析色素腺体形成与棉酚合成关键酶基因在比克氏棉色素腺体形成过程中的表达特点,探究比克氏棉色素腺体形成的分子机理,为利用该性状提供依据。
1 材料与方法
1.1 试验材料
本研究供试的材料为野生二倍体棉种比克氏棉(G染色体组),对照材料包括陆地棉有色素腺体标准系TM-1、陆地棉有色素腺体中棉所12及其无色素腺体品种近等基因系中棉所12无,均为本实验室自交保存的种质系。
1.2 试验方法
1.2.1材料取样及密度大小测定。比克氏棉和陆地棉TM-1于开花期挂牌标记。取充分成熟的比克氏棉种子进行浓硫酸脱绒,在种子的合点处切口,28℃条件下浸泡12 h后剥去种皮,置于光照培养箱(培养条件:16 h光照,8 h黑暗;28℃)中发芽(种子浸泡12 h后发芽为种子萌发的时间起点),从种子萌发开始至种子萌发48 h,每隔2 h取子叶用于显微观察其色素腺体情况。在体式显微镜下观察种子萌发后不同时间点的子叶及开花当天各材料幼嫩的叶片、花瓣、萼片和苞叶等器官并拍摄照片,使用Image Pro Plus 6.0测定腺体密度和大小。每种材料3次重复。
1.2.2组织切片观察。比克氏棉开花后每天取比克氏棉种胚放在FAA(Formalin-aceto-alcohol)固定液里固定保存。分别取萌发0、2、4、6、8、10、12、14、16、18、20、22、24、36和48 h的 子 叶 固 定 于FAA溶液中用于石蜡切片(萌发条件同1.2.1)。同时取比克氏棉种子10粒,在完全黑暗条件下进行萌发,并观察在无光照情况下各个时间点子叶色素腺体的形态变化。本研究采用毛晓霞的石蜡切片方法[14],经脱水、包埋、切片、染色后置于体式显微镜下观察及拍照。
1.2.3棉酚含量测定。将比克氏棉、陆地棉有色素腺体棉中棉所12及其无色素腺体近等基因系中棉所12无进行开花期挂牌标记,各材料开花10 d以后至开花后50 d每天收取种胚,将胚珠放在烘箱(35℃)中烘干,烘干后将材料磨成粉并利用80目筛(孔径0.178 mm)过滤后低温避光备用。分别取萌发1、2、3、4、5、6、7、8和9 d的子叶放在纸袋中(萌发条件同1.2.2),置于35℃烘箱烘干,磨粉过80目筛备用。精确称取不同样品0.1 g,按照刘慧等的方法[15]进行棉酚提取及色谱测定。每种材料3次重复。
棉酚标准品购自于Sigma公司,左旋棉酚标准品购自山西慈缘生化科技有限公司。其他试剂购自天津四友精细化学品有限公司。
1.2.4SPSS统计分析。利用SPSS 19.0软件,分析种子萌发期色素腺体大小、密度与棉酚含量的差异性和相关性。
1.2.5荧光定量PCR分析。利用艾德莱植物RNA提取试剂盒提取比克氏棉种子萌发0、12、24、36和48 h的子叶总RNA,利用PrimeScript First Strand cDNA Synthesis Kit(购自Takara公司)反转录试剂盒合成cDNA第1链,使用SYBER Premix Ex TaqTM(购自Takara公司)进行实时定量PCR(Polymerase chain reaction)分析。实时定量PCR引物利用Prime Premier 5.0设计,内参基因使用UBQ7,在擎科生物科技有限公司完成引物合成,进行3次生物学重复。实时定量PCR引物如表1所示。
2 结果与分析
2.1 色素腺体形成
2.1.1种胚发育过程中子叶色素腺体原的形成。图1A和1B是比克氏棉种胚形成过程的石蜡切片。在种胚形成前期(开花后15 d之前),比克氏棉子叶上未观察到成熟的色素腺体,但开花后15 d,其子叶上出现一些比正常细胞小、无规则排列的细胞群,即色素腺体原(图1A和1B)。这种色素腺体原随着种胚的发育略有变化,少量子叶边缘处的色素腺体原中间形成小腔,大多色素腺体保留无腔至种子休眠。因此,开花后15 d为比克氏棉子叶色素腺体原形成的时间节点(杭州,气温28~30℃)。

表1 荧光定量PCR的引物Table 1 All primers used in this study

图1 比克氏棉子叶色素腺体形态建成过程Fig.1 Morphogenesis of cotyledon pigment glands of G.bickii
2.1.2种子萌发过程中子叶色素腺体形态建成。比克氏棉的休眠种子中无成熟的色素腺体,但具有色素腺体原结构(图1C)。对其进行连续石蜡切片,光学显微镜下观察发现比克氏棉种仁中共存在2种色素腺体细胞群结构:1种是含有腔体的色素腺体原结构,另1种是比其他细胞相比略小并成团的细胞群结构,即没有腔体的色素腺体原。对萌发后不同时间的种子进行连续石蜡切片,观察发现,种子萌发10 h,子叶上的色素腺体细胞已看不清完整的细胞形态,出现了染色极深的厚壁(图1D)。种子萌发12 h,子叶中的色素腺体原开始出现空腔(图1E)。种子萌发24 h,子叶上的色素腺体原已基本形成成熟的色素腺体,且腔体内部出现内含物;此时肉眼也可以观察到色素腺体。然而,比克氏棉子叶上除了色素腺体原发育成的色素腺体外;还有新形成的色素腺体,这类色素腺体在形成过程中无色素腺体原阶段或色素腺体原阶段十分短,与其他器官所形成的色素腺体并无明显差异。
2.1.3光照对种子萌发过程中色素腺体形态建成的影响。对置于完全黑暗条件下萌发48 h的比克氏棉种子进行连续的石蜡切片,研究光照对于比克氏棉子叶色素腺体形成的影响。结果(图1F)表明,黑暗条件下在光学显微镜中子叶仍可观察到色素腺体的结构,且与正常条件下形成的腺体结构无明显差异,只是黑暗条件下的子叶色素腺体结构在种子萌发48 h形成,其形成的时间明显晚于光照条件。说明光照不会影响子叶色素腺体的形成,但可能对色素腺体原转变成成熟的色素腺体有一定的延迟作用。
2.1.4子叶成长过程中色素腺体表现。比克氏棉种子在28℃条件下浸泡12 h后剥去种皮,置于28℃恒温箱中开始发芽(即种子萌发的时间起点)。种子萌发0 h无肉眼可见的色素腺体(图1G),萌发12 h前无肉眼可见的色素腺体(图1H),萌发24 h子叶有肉眼可见的色素腺体(图1I)。色素腺体的产生由子叶边缘向中间扩展,边缘的色素腺体密度高于中央。
对比克氏棉种子萌发不同阶段的子叶色素腺体密度和大小进行统计分析,结果表明,在种子萌发22 h后,子叶色素腺体数量开始增加,随后保持在252个·cm-2左右。萌发34 h继续增加至312个·cm-2。从色素腺体大小来看,种子萌发22 h开始,随着萌发时间的增加,色素腺体也呈增大趋势,至28 h,色素腺体直径略有减小,30 h时色素腺体直径最大可达75μm,随后降低保持在49 μm水平(表2)。比克氏棉子叶色素腺体密度及直径相关性分析结果表明,随着种子萌发时间的增加,子叶色素腺体密度和大小均有所增加,在种子萌发的不同时期,比克氏棉子叶色素腺体大小与密度无相关性,这与祝水金等[16]的结论一致。
2.1.5其他主要器官的色素腺体表现。对比克氏棉植株各主要器官的色素腺体密度和大小进行观察(表3)。比克氏棉休眠种子中无肉眼可见的色素腺体,而成熟的植株各器官可观察到色素腺体,说明比克氏棉具有种子无色素腺体,植株有色素腺体的性状。比克氏棉苞叶为针状形,无色素腺体;而花瓣、萼片、子叶、铃壳上色素腺体较稀疏;下胚轴和叶片上色素腺体较密集。从色素腺体大小来看,花瓣色素腺体最小,子叶、下胚轴、萼片的色素腺体大小适中,铃壳上的腺体最大。
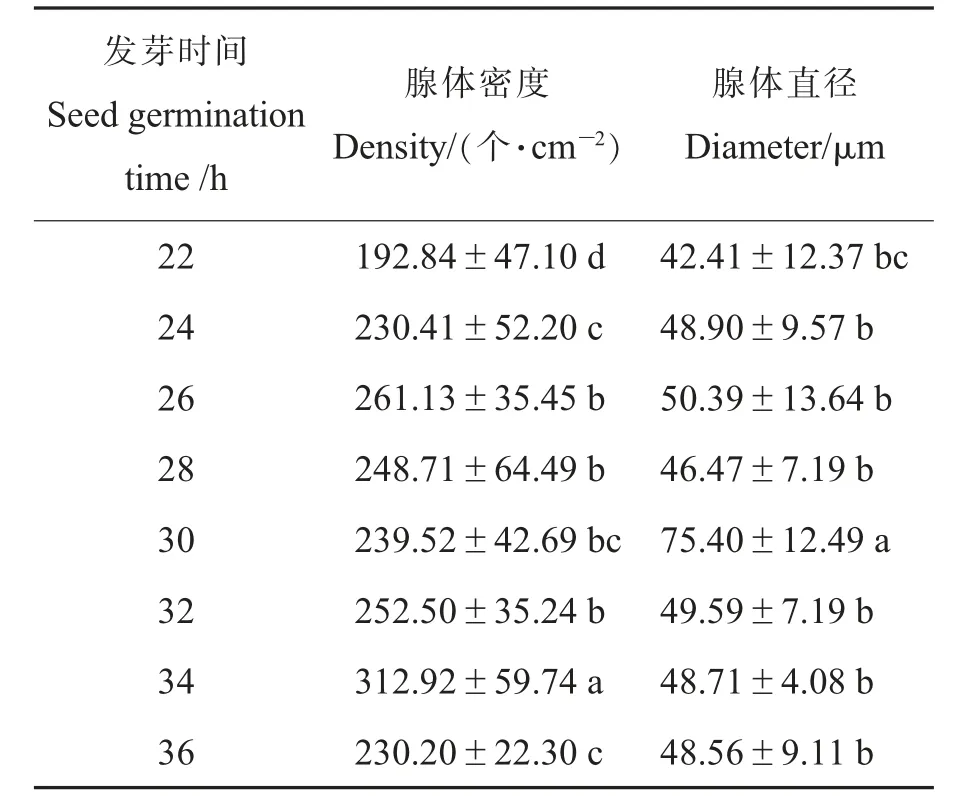
表2 比克氏棉种子萌发过程中子叶色素腺体密度和大小Table 2 The density and dimeter of the pigment glands during seed germination of G.bickii
2.2 棉酚代谢
2.2.1种胚发育过程中棉酚动态。色素腺体与棉酚的合成有一定的关系。植株各主要器官的棉酚含量和色素腺体的密度呈正相关,因此一些学者认为棉酚可作为色素腺体的生化指标[17]。但棉酚含量与色素腺体密度并无直接关系。对比克氏棉种胚发育过程中的棉酚含量进行测定,并与陆地棉有色素腺体(中棉所12)和无色素腺体近等基因系中棉所12无进行对照。从表4可以看出,有色素腺体的中棉所12及其无色素腺体的近等基因系中棉所12无在种子成熟后期(开花后25 d)均可检测到棉酚,只是无色素腺体棉种子成熟后几乎检测不到棉酚,而有色素腺体棉种子中的棉酚含量一直呈上升趋势,到种子成熟后期(开花后50 d)达到0.922 0%。说明有色素腺体棉和无色素腺体棉在种胚发育过程中棉酚动态积累存在差异,由于有色素腺体棉在种胚发育后期色素腺体腔的形成使得棉酚大量积累,因此有色素腺体棉种子为高酚型,而无色素腺体棉在种子发育过程中不形成色素腺体,因而棉酚含量较低,为低酚型。比克氏棉在种胚发育过程中均未检测到棉酚,说明比克氏棉与无色素腺体棉种发育过程前期棉酚动态积累类似,因未形成色素腺体导致棉酚无法积累。比克氏棉和无色素腺体棉中棉所12无的成熟种子中棉酚含量均低于国家卫生标准(0.04%),说明比克氏棉与无色素腺体棉种相同,种子为低酚类型。

表3 比克氏棉、TM-1植株主要器官的色素腺体密度和大小Table 3 The density and dimeter of the pigment glands in the main organs of G.bickii and TM-1
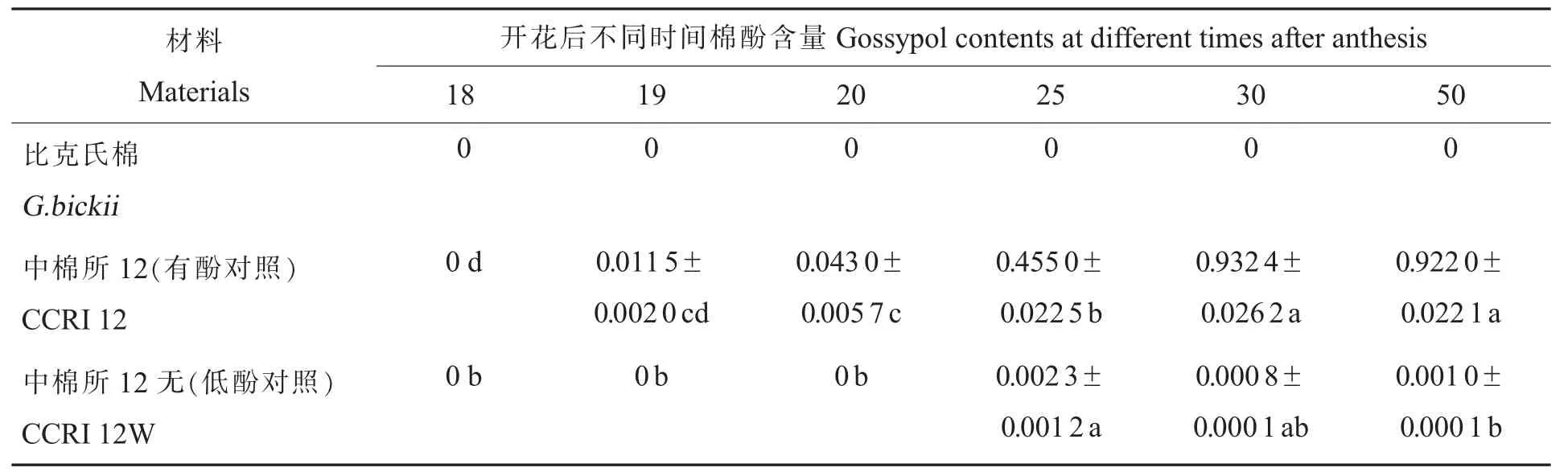
表4 不同棉种种子成熟各阶段的棉酚含量Table 4 Gossypol contents during seed maturation of different species of cotton %
2.2.2子叶色素腺体形态建成过程中的棉酚动态。对比克氏棉种子萌发不同阶段的子叶进行棉酚含量测定,结果(图2)表明,休眠种子中未检测到棉酚,种子萌发12 h开始,子叶中可以检测到棉酚,但非常低,只有0.045 0%。种子萌发24 h,子叶棉酚含量略有下降,保持在0.045 0%左右的水平。直到种子萌发72 h,子叶棉酚含量开始急剧增加至0.075 5%,随后棉酚含量趋于稳定,至种子萌发第8 d,棉酚含量达到最大值0.085 5%。比克氏棉种子萌发的不同阶段,子叶棉酚的动态积累与无色素腺体棉相似,但与有色素腺体棉相反[20]。
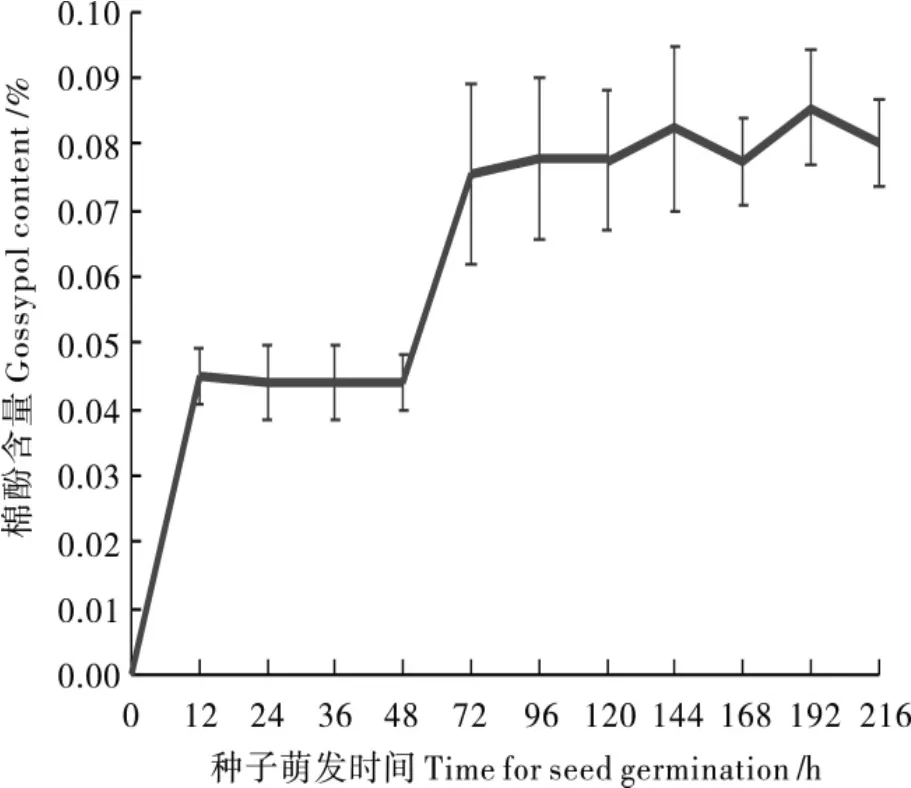
图2 比克氏棉种子萌发过程中的棉酚含量Fig.2 The contents of gossypol during seed germination of G.bickii
比克氏棉种子萌发12 h已形成色素腺体结构,同期可检测到棉酚的存在,但棉酚含量极低,说明棉酚合成与腺体形成在时间上是同步进行的,该发现与之前的结论有所差异[18]。在种子萌发48 h,比克氏棉的腺体结构已完全形成,此时棉酚含量开始急剧增加,这证明了色素腺体是棉酚的贮藏器官。
2.2.3比克氏棉植株主要器官的棉酚含量。由表5可以看出,有色素腺体棉(中棉所12)及其无色素腺体的近等基因系(中棉所12无)在植株的主要器官中均可检测到棉酚,且棉酚含量均高于国家卫生标准(0.04%)(除中棉所12无的种仁外),说明有色素腺体棉(中棉所12)及其无色素腺体的近等基因系(中棉所12无)植株均为有酚型。植株的各器官棉酚含量有所差异,中棉所12种仁棉酚最高含量为0.964 1%,子叶和花瓣棉酚含量较高分别为0.350 5%和0.223 0%,其余器官棉酚含量保持在0.096 6%~0.149 1%。中棉所12无子叶中棉酚最高含量为0.170 3%,叶片、苞叶、萼片、铃壳棉酚含量在0.120 0%左右,花瓣中含量较低为0.046 7%,种仁中棉酚仅为0.005 1%。比克氏棉植株除种仁、苞叶(比克氏棉苞叶为针型,无法检测棉酚含量)外的主要器官中也可检测到棉酚,且棉酚含量高于国家卫生标准,因此比克氏棉植株也为有酚型。比克氏棉萼片棉酚最高含量达0.312 1%,铃壳、叶片及子叶棉酚含量较高均超过0.180 0%,花瓣棉酚最低含量为0.054 1%,种仁中无法检测到棉酚。因此比克氏棉属于种子低酚型、植株有酚型品种。

表5 不同棉种各器官的棉酚含量Table 5 Gossypol contents in the main organs of G.bickii and G.hirsutum %
2.3 色素腺体形成和棉酚合成相关基因的表达特点
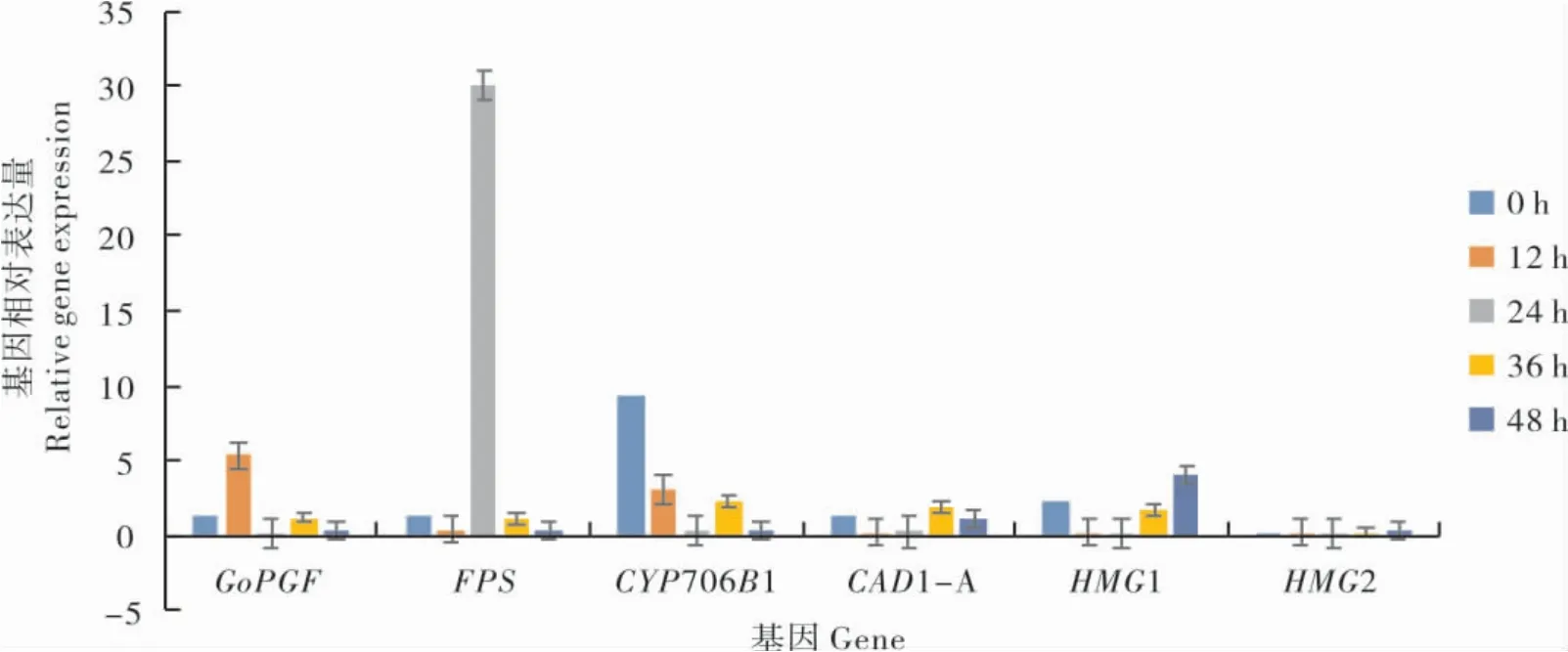
图3 色素腺体形成及棉酚合成基因相对表达量Fig.3 Relative expression of pigment gland formation and gossypol biosynthesis genes
利用实时定量PCR技术检测色素腺体形成基因(GoPGF)、棉酚合成基因(FPS、CYP706B1、CAD1-A、HMG1、HMG2)在比克氏棉种子萌发的不同时期(0、12、24、36和48 h)的表达情况。从图3可以看出,与色素腺体形成相关的GoPGF基因在种子萌发12 h时的相对表达量显著高于0 h和36 h,但在24 h及48 h几乎不表达,说明比克氏棉子叶色素腺体形态建成过程中,GoPGF基因参与了色素腺体原发育成成熟的色素腺体的过程。棉酚合成过程中的关键基因,FPS基因在24 h相对表达量最高,其他时期几乎无表达。CYP706B1基因在0 h相对表达量较高,在12 h和36 h有一定程度的表达;HMG1基因在48 h相对表达量较高;而CAD1-A及HMG2基因在种子萌发各个时期几乎不表达,说明这两个基因与比克氏棉种子色素腺体形成无密切关系。
3 讨论
由于植棉收益大幅下降,棉花种植面积下滑已成为棉花生产的严重问题。棉籽中含有高质量的蛋白质和脂肪,通过开发利用棉籽,可将棉花应用于工业、畜牧业及食品行业,从而提高棉花的综合利用价值,对棉花产业的可持续发展有重要的意义[19]。多年来,棉花育种家致力于通过传统育种技术培育全株有色素腺体的低酚棉品种,以充分利用棉籽蛋白质和油脂,并取得了长足的进展[20-21]。然而,由于棉花全株无色素腺体缺乏对多种逆境有拮抗作用的棉酚及其衍生物,限制其生产应用。比克氏棉等澳洲野生棉种具有1种特殊的子叶色素腺体延缓形成性状,即种子无色素腺体而植株有色素腺体,对于棉籽综合利用具有重要意义[7]。然而,由于遗传亲缘关系较远,加之目标性状与棉酚代谢之间的关系及目标性状的调控机制不清楚等原因,直接杂交转育难以成功[22-24]。低酚棉品种具有高蛋白、高油脂和高综合利用率等优点,但棉酚含量较低,对昆虫的防御能力较弱。种子无棉酚、植株有棉酚的棉花品种不仅具有低棉酚品种的优点,又具有抗虫性。如能培育出植株高棉酚,种子低棉酚的棉花品种并利用该品种进行选育,不仅可以实现棉花的综合利用,而且还可以实现其重大的社会价值及经济价值。因此种子无棉酚、植株有棉酚的品种是育种家工作的重中之重。但棉酚与色素腺体之间的关系错综复杂,需要更多的研究。
本研究结果表明,比克氏棉的色素腺体与有酚棉色素腺体的形成方式不同。比克氏棉种胚发育期(开花后15 d)前期,已形成色素腺体原,并保持不分化状态,大部分色素腺体为无腔体结构,仅子叶边缘的很少一部分色素腺体原形成小腔结构。由于无腔体结构,色素腺体原无法贮藏棉酚及其衍生物,因此其休眠种子无肉眼可见的色素腺体,检测不到棉酚的存在,为无酚棉种子。绝大多数色素腺体原在比克氏棉种子萌发后12 h,中间细胞解体成腔体,开始分化形成成熟的色素腺体。之后,随着种子萌发,子叶生长,子叶上又开始出现部分色素腺体,这一类色素腺体与其它棉种及比克氏棉其它器官的色素腺体相同,不存在色素腺体原结构,或色素腺体原期十分短暂。值得提出的是,种子萌发12 h,通过基因表达产生的酶类十分罕见。可见,分解比克氏棉色素腺体原的酶可能并不是由于种子萌发而激活基因表达而产生的,而是在种子中或色素腺体原细胞中就已存在,种子萌发只是使其激活而已。暗处理发芽延缓了色素腺体原腔体形成,但未能完全抑制腔体形成。因此,激活色素腺体原细胞解体的因素还有待进一步的研究。
比克氏棉种子萌发后12 h的子叶上可以检测到棉酚,而这一时期正是色素腺体形态建成的关键时期,即色素腺体原形成腔体,棉酚及其衍生物转运至腔体内。然而,休眠种子并没有检测到棉酚,而且棉花种子萌发12 h还未有合成棉酚的能力。因此,这部分棉酚可能是以另一种不能被相应的光谱所检测到的方式存在,腔体形成后即逆转成能被其光谱检测到的棉酚,并存存于色素腺体内。由此也说明棉酚是一种不稳定的化学物质,只有在色素腺体内才稳定储存。
比克氏棉在种子萌发后12 h开始形成色素腺体厚壁,种子萌发后24 h已形成完整的色素腺体腔且腔内有内含物,种子萌发36 h肉眼可观察到较多的色素腺体。48 h时棉酚含量开始稳定上升,因此种子萌发48 h,比克氏棉色素腺体形态已经建成且表达充分。对比克氏棉种子萌发不同时期的子叶进行qRT-PCR表达谱分析结果表明,GoPGF基因在比克氏棉种子萌发12 h相对表达量较高,说明色素腺体腔的形成需要GoPGF基因的参与。
棉酚主要是在根部合成的[25]。然而,比克氏棉种子萌发过程其棉酚合成相关基因CYP706B1、FPS和HMG1基因均有不同程度的表达。其中FPS基因在24 h相对表达量最高,其它时期几乎无表达,推测该基因与棉酚的转运有关系。而CAD1-A及HMG2基因在种子萌发期几乎不表达,这明这两个基因与比克氏棉种子色素腺体形成无密切关系。
4 结论
本研究以具有子叶色素腺体延缓发生性状的比克氏棉为材料,探究色素腺体形态建成特点及色素腺体与棉酚含量之间的相关性,研究发现种子萌发后的12 h为比克氏棉子叶色素腺体形态建成的关键时期,且色素腺体的形态建成与棉酚出现具有同步性。这有助于了解色素腺体形成与棉酚代谢的动态关系。GoPGF基因与色素腺体原形成色素腺体有关,FPS基因参与色素腺体的形成和棉酚向子叶色素腺体转运,CYP706B1基因和HMG1基因与种子萌发后色素腺体腔的形成有关,并参与棉酚的贮运。这些基因对培育棉花新品种提供重要的理论依据。
