弱光胁迫下鄞红葡萄生理生化特性及相关基因的研究
2019-01-09何静雯吴燕燕沈梓力吴月燕
何静雯,李 晨,邱 甜,吴燕燕,沈梓力,吴月燕
(1.浙江万里学院 生物与环境学院,浙江 宁波 315100;2.上海海洋大学 水产与生命学院,上海 201306)
葡萄(Vitisvinifera)为多年生木质藤本植物,是世界上种植最广泛的果树树种之一[1]。而鄞红葡萄为宁波地区设施栽培特有品种,由优良变异单株选育而来,属欧美杂交种[2-3]。该葡萄品种的栽培主要采用简易型日光温室,可抵御环境污染和台风等。但由于设施条件的限制和季节性不良气候的影响,温室环境通常具有透光量低、透光分布不均、光质差、紫外线含量低等缺点,会造成温室内部分葡萄采光不足。光照不足会严重影响葡萄生理生化过程及形态建成,导致产量和品质下降[4]。植物在长期进化过程中会形成一系列的自我保护机制,以抵御胁迫环境的伤害[5],弱光环境下植株生长受阻、但会通过叶面积增加来抵御弱光胁迫[6]。另外,植物细胞保护酶活性的高低以及渗透调节物质的合成状况更是植物对胁迫环境响应的重要指标[7]。通常植物细胞内的超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)会在一定的胁迫程度内升高,丙二醛(MDA)、脯氨酸(PRO)含量也会对胁迫环境显著响应[8-9]。近年来,植物抗逆基因的研究也成为众多植物领域的热门,MYC2转录因子(MYC2 BHLH protein)与h型硫氧还原蛋白(Thioredoxin h-type)在胁迫环境下反应活跃,能够调节许多相关因子以抵御胁迫环境[10-11]。由此可见,弱光响应的理化分析与相关基因表达量的测定是植物弱光胁迫研究的关键。目前,国内外有关弱光的研究多是从弱光胁迫对植物的生理生化、光合作用、同化力的形成等方面入手,且主要集中在辣椒、番茄、茄子等蔬菜作物上[12-13],对葡萄生理生化以及相关基因的研究相对较少。
本研究以鄞红葡萄幼苗为材料,研究了其在不同遮阴程度下的生长形态变化、细胞保护酶活性、渗透调节物质以及MYC2与TRX基因相对表达量,以阐释鄞红葡萄对弱光胁迫产生响应的机制,旨在为今后葡萄相关品种的光照管理奠定理论基础。
1 材料和方法
1.1 试验材料
供试品种为150株长势均匀的1年生鄞红幼苗。
1.2 试验设计
温度恒定在38 ℃,相对湿度65%。瓦盆高 28.5 cm, 内径 25 cm,人工基质培养。基础营养成分为有机质5 530 mg/kg、水解氮 43.6 mg/kg、速效磷 11.5 mg/kg、速效钾93.7 mg/kg。定植前施入复合肥(N∶P∶K =15∶11∶9),主梢长至50 cm时摘心,副梢留2叶摘心。以遮阳网和塑料薄膜为覆盖材料进行遮阴处理,遮阴程度设置5个水平,即 CK(遮光率为0、光照强度28 000 lx)、T1(遮光率25%、光照强度约22 000 lx)、T2(遮光率40%、光照强度约17 000 lx)、T3(遮光率70%、光照强度约10 000 lx)、T4(遮光率85%、光照强度约5 000 lx)。在遮阴处理后的0,10,20,30 d观察并采样,进行各种理化指标的测定,每组指标均重复3次并取平均值。
1.3 测定指标与方法
1.3.1 鄞红叶片生长指标的测定 每处理组随机选取5株,将根部和叶片完整剪下,采用LA-S全能植物图像分析系统(广州深华公司)测定根系长度、直径、表面积、叶面积。
1.3.2 鄞红叶片保护酶及渗透调节物质的测定 可溶性蛋白测定采用考马斯亮蓝G-250法[14];丙二醛测定采用硫代巴比妥法[15];脯氨酸测定采用茚三酮显色法[16];超氧化物歧化酶测定采用氮蓝四唑(NBT)法[15];过氧化氢酶测定采用紫外吸收法[17-18];过氧化物酶测定采用愈创木酚比色法[19]。
1.3.3 弱光胁迫下鄞红相关基因表达量测定 遮阴处理30 d后,测定葡萄叶片在不同遮阴程度下MYC2与TRX基因表达量的变化。按照植物总RNA提取试剂盒(OMEGA公司,R6827-01)说明书提取叶片总RNA。采用NanoDropTM 2000分光光度计(Thermo Fisher, USA)测定总RNA的质量和纯度,1.5%琼脂糖凝胶电泳鉴定其完整性。吸光度OD260/280值在1.9~2.1、电泳有3条亮带,且28S条带亮度大约是18S 的2倍,其总RNA为高质量RNA,可进行cDNA第1链合成试验。以提取的高质量总RNA为模板,使用SuperRT cDNA第1链合成试剂盒(SuperRT cDNA Kit CW0741, CWBIO,China)反转录合成cDNA第1链,具体操作步骤按说明书进行。
根据已报道的葡萄MYC2 (NC_012021.3)和TRX(NW_003724430.1)基因序列,采用Primer软件分别设计荧光PCR引物(表1)。数据处理采用相对值2-ΔΔCt进行计算,采用GraphPrism 7.0软件处理数据和作图,采用SPSS 23.0软件对数据进行方差分析,用LSD法进行多重比较。
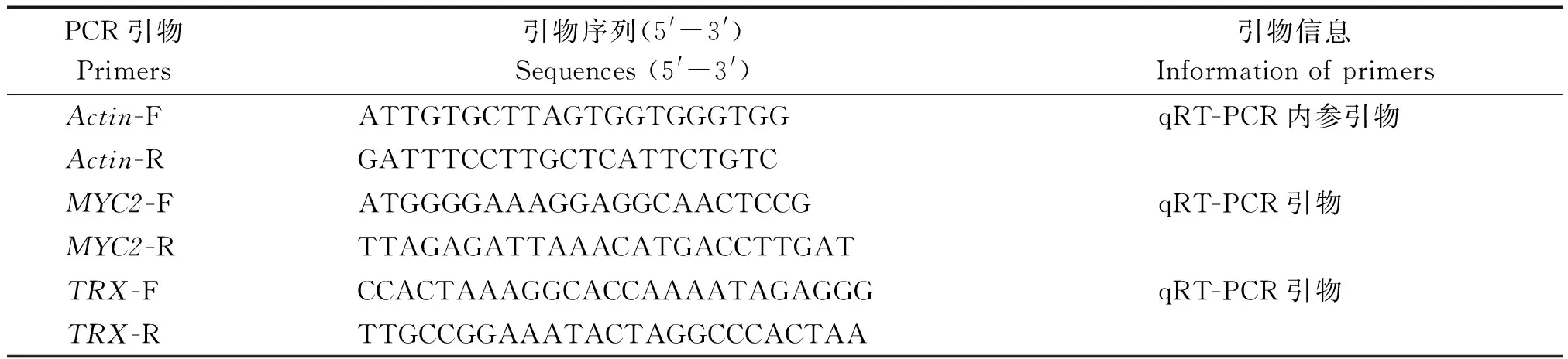
表1 荧光定量PCR基因扩增引物序列Tab.1 Primer sequences used for Q-PCR
2 结果与分析
2.1 弱光胁迫对鄞红叶片生长特性的影响
试验过程中与CK相比,T1叶片生长状况良好,T2叶片生长状况一般,T3生长状况下降明显,叶片出现一定的黄斑和脱落现象,与对照组相比差异显著,但仍然能够继续生长。其中,弱光环境下,T2的叶面积会随着弱光胁迫的增加而变大,与对照组相比差异显著(表2)。但随着遮阴程度的进一步加强,T4叶片出现大量黄斑和脱落,生长严重受阻,各项指标均低于对照组。因此,一定程度的遮阴有利于叶片生长,但遮阴过强会对植株造成不可恢复的伤害。
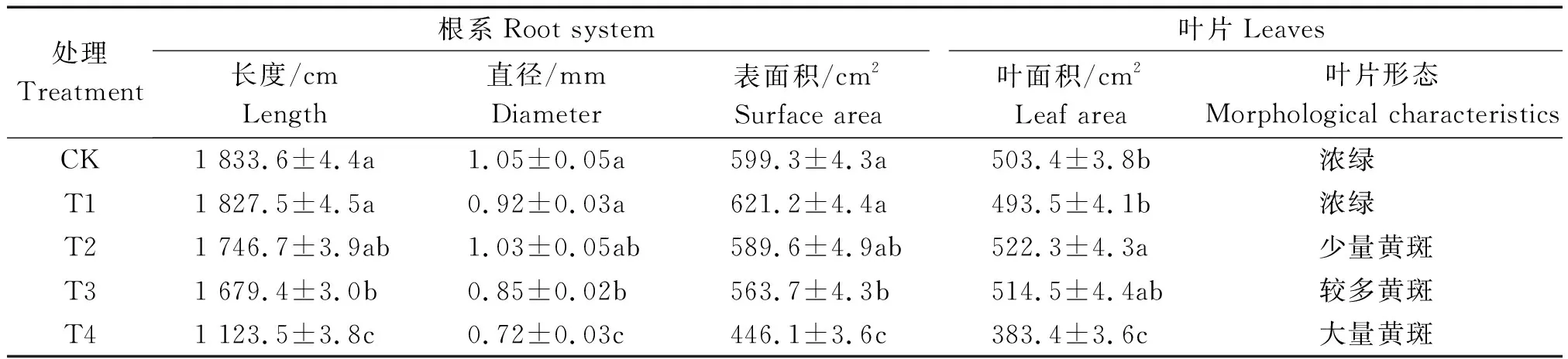
表2 弱光胁迫对鄞红叶片生长特性的影响Tab.2 Effects of low light stress on the growth characteristics of Yinhong grape
注:不同字母代表差异显著(P<0.05)。图1-4同。
Note: The different letters indicate significant difference(P<0.05).The same as Fig.1-4.
2.2 弱光胁迫对鄞红葡萄叶片可溶性蛋白含量的影响
弱光胁迫下,T1与T2可溶性蛋白质含量的变化趋势与CK相似,均略微升高后维持稳定(图1),在30 d处理结束后分别比对照上升 2.5%和5.7%,与CK相比差异不显著(P>0.05)。但随着遮阴程度的增加,在T3与T4组中,叶片可溶性蛋白含量随遮阴时间增长呈逐渐下降的趋势,试验结束时分别下降22.7% 和 36.1%,均显著低于对照组(P<0.05)。
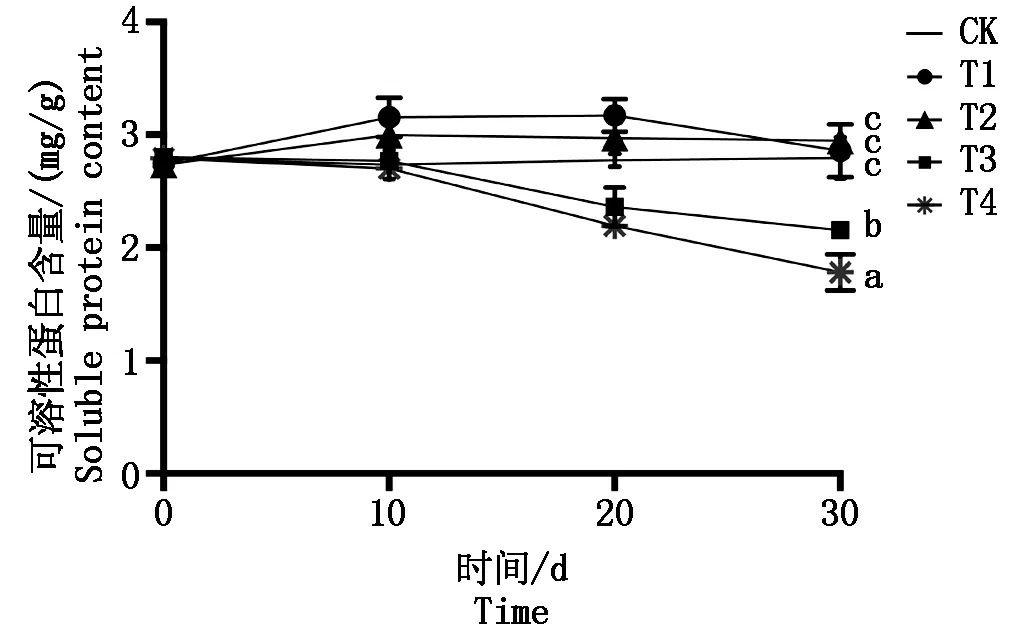
图1 弱光胁迫对鄞红葡萄叶片可溶性蛋白质含量的影响Fig.1 Effects of low light stress on the soluble protein content in Yinhong grape leaves
2.3 弱光胁迫对鄞红葡萄叶片丙二醛和脯氨酸含量的影响
遮阴结束后各处理组丙二醛、脯氨酸含量变化呈逐渐上升趋势(图2),其中丙二醛含量依次逐渐上升,从T1~T4相比CK分别上升了23.5%,34.8%,52.8%,98.5%,处理组与对照组相比除T1处理外差异显著(P<0.05)。脯氨酸的含量也随遮阴程度的增加而逐渐升高,试验结束时T1~T4分别比CK上升了27.1%,35.5%,63.6%,95.2%,处理组与对照组相比差异均显著(P<0.05)。
2.4 弱光胁迫对鄞红葡萄叶片保护酶活性的影响
随着遮阴程度的增强,叶片中SOD、POD、CAT活性均呈先升高后降低的趋势(图3)。但在遮阴处理结束时,鄞红叶片T1与T2组中SOD、POD、CAT含量相比CK分别下降 5.4%,4.8%,2.7% 和 17.1%,9.1%,3.9%,但差异不显著(P>0.05);但随着遮阴程度的增加,T3与T4组叶片保护酶活性呈持续降低状态,在处理结束时SOD、POD、CAT分别比CK下降了27.0%,30.5%,22.3% 和 52.8%,49.9%,48.7%,均显著低于对照组(P<0.05)。
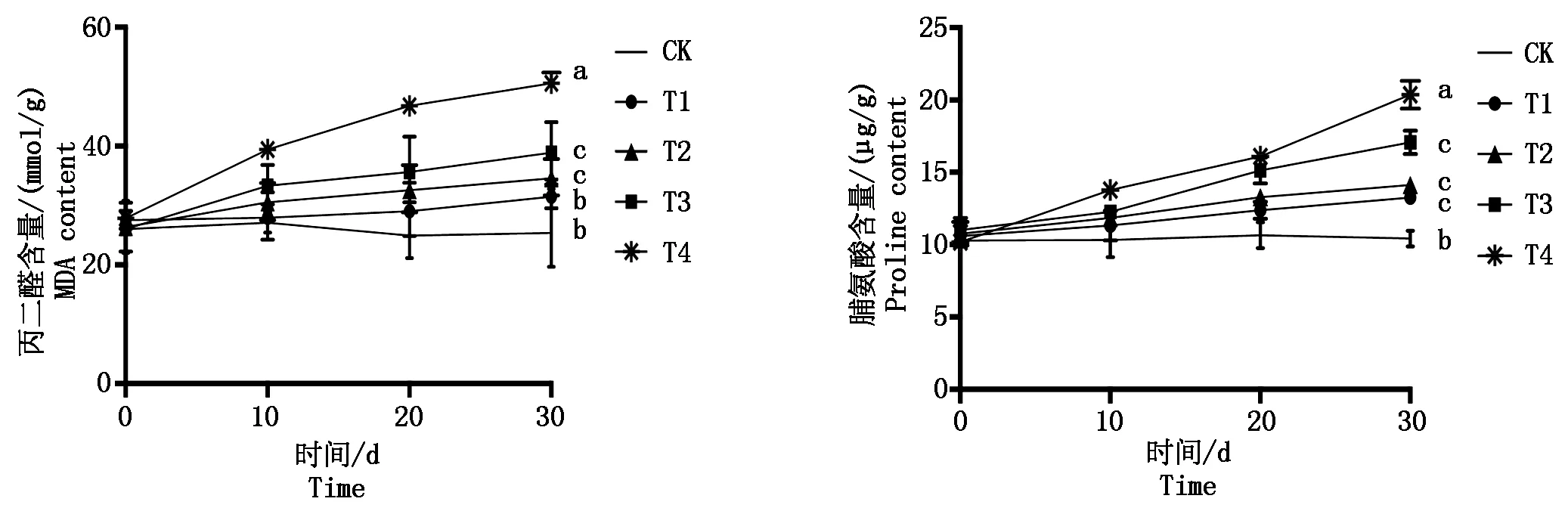
图2 弱光胁迫对鄞红葡萄叶片中丙二醛和脯氨酸含量的影响Fig.2 Effects of low light stress on the MDA content and proline content in Yinhong grape leaves
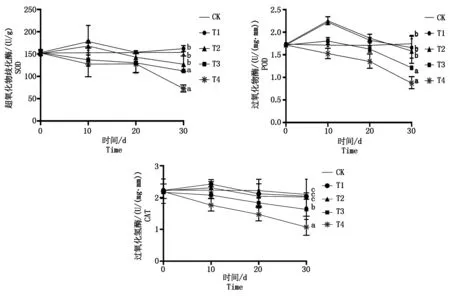
图3 弱光胁迫对鄞红葡萄叶片保护酶活性的影响Fig.3 Effects of low light stress on the protective enzyme activities in Yinhong grape leaves
2.5 MYC2与TRX基因表达量分析
30 d处理结束时,对鄞红葡萄叶片MYC2与TRX基因表达量的测定发现(图4),与CK相比MYC2与TRX基因的表达量均先升高后降低,这与CAT和可溶性蛋白的变化趋势大致相当。在T1处理下MYC2与TRX基因表达量显著上升,由此证明适度遮阴会通过二者上调表达来抵御胁迫环境,但随着遮阴程度的增强表达量又显著降低(P<0.05),重度弱光胁迫已经超出了抗逆体系的承受范围。
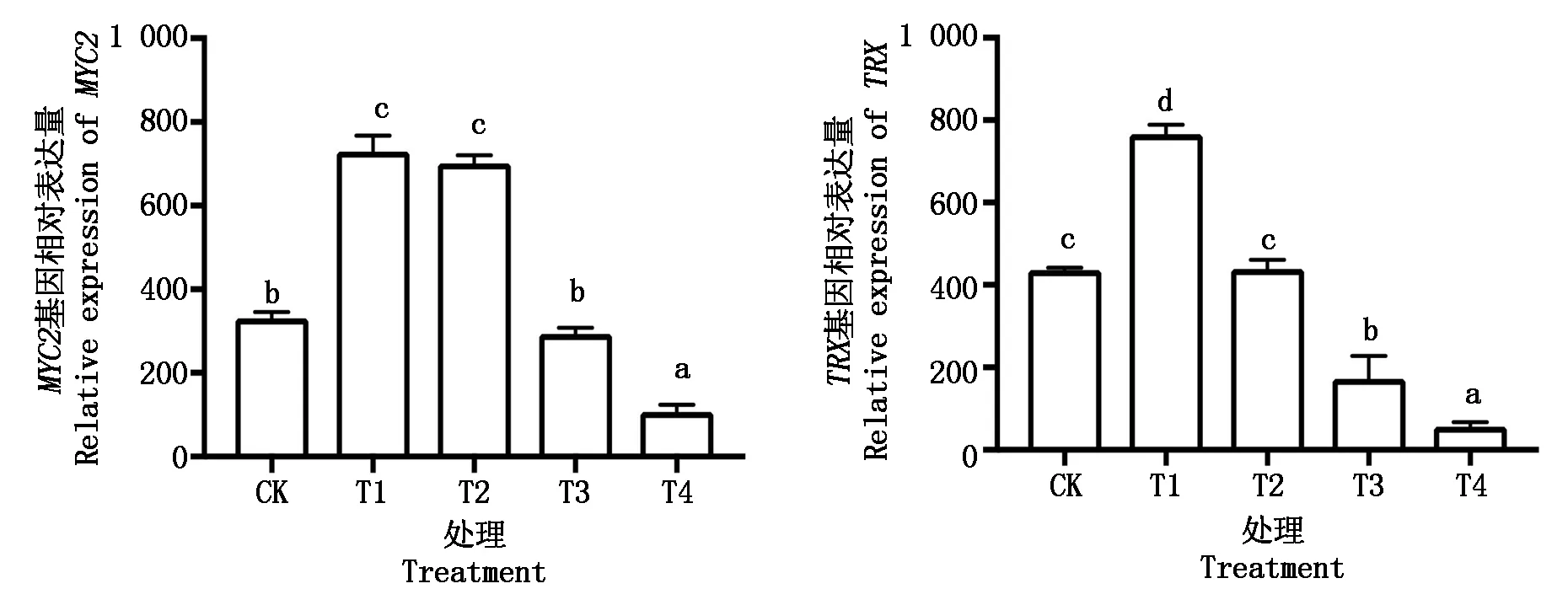
图4 弱光胁迫下MYC2与TRX基因表达量的变化Fig.4 Relative expression of MYC2 and TRX under low light stress
3 结论与讨论
光是植物生长发育过程中的重要因子,通常弱光处理下的植株根系受阻[2],同时会出现叶片变薄、叶色变浅、叶面积增加等现象[13]。唐韡等[20]研究发现,弱光胁迫下番茄根系活力下降。蒋晓婷等[21]对丝瓜研究发现,遮阴处理使植株根冠比显著下降、同化产物积累减少,植株干质量降低。丘立杭等[22]对甘薯的研究表明,弱光胁迫显著抑制甘蔗茎粗,使茎径不断减小。易金鑫等[7]研究发现,弱光胁迫下茄子叶面积增加,生长受阻。本研究中,鄞红葡萄在T1处理下生长状况良好,对弱光环境抗性较高,植株通过增加叶面积捕获更多光能制造有机物。但随着遮阴程度的增强,植株各项生长指标降低,发育减缓,这与常静等[8]对辣椒的研究结论大致相符。
弱光胁迫下葡萄叶片内可溶性蛋白、脯氨酸等渗透调节物质均会发生变化。一定胁迫程度下植物体内蛋白质合成受阻,但植物也会通过增加蛋白质的含量以适应胁迫环境[22]。脯氨酸作为重要的渗透调节物质之一,在胁迫环境下降低植物细胞水势、调节细胞微环境、提高胞内代谢反应[23-24]。候永平等[25]研究发现,低温弱光处理西葫芦后可溶性蛋白含量显著降低,这与迟伟等[26]对草莓的研究结果一致。本研究中,鄞红葡萄叶片可溶性蛋白先升高后降低,由此证明该品种对弱光具有一定的适应性;而脯氨酸含量随遮阴程度的增强逐级递增,以此表明叶片的受害程度不断增强,这与陈磊等[27-28]对于茄子的研究结果大致相当。
MDA是判断植物细胞膜脂过氧化程度的重要指标,胁迫环境会使MDA大量积累,从而导致细胞受损[29],而SOD、POD 和 CAT 等酶是细胞内清除活性氧和自由基的主要保护酶,这些酶与植物的抗逆系统密切相关[30]。其中,SOD会抵御逆境胁迫下植株内超氧阴离子的增加,POD与CAT参与多种抗氧化代谢,有控制植株生长以及分解 H2O2等作用,使植物细胞免受毒害[31]。任华中等[32]在番茄的研究中表明,细胞保护酶活性随光照强度的减弱而升高。刘永华等[33]对瓠瓜遮阴处理后研究发现,MDA与抗氧化酶活性与光照强度呈负相关。本研究中,MDA 含量随胁迫程度的增加而升高,SOD、POD 和 CAT 活性先升高后降低;但POD 变化趋势没有SOD 和CAT 明显,这可能是由于 POD 既参与自由基的消除,同时也参与活性氧的生成[34-35]。因此,适度弱光胁迫会激活植物体内抗逆系统,促使保护酶活性上升;但重度胁迫会导致细胞受损,从而无法合成细胞保护酶,此试验结果与宋金亮等[36]对西葫芦的研究结果一致。
MYC 2转录因子与硫氧还蛋白(TRX)都与胁迫环境密切相关。其中MYC 2转录因子是植物 bHLH 转录因子家族中的一员,该转录因子是茉莉酸(JA)信号途径中的核心因子,在生物或非生物胁迫下通过激活相关转录因子的活性来抵御逆境胁迫[37]。Sasaki-Sekimoto等[38]研究发现,JA能够诱导拟南芥抗坏血酸合成途径中关键酶基因的表达,证明MYC 2转录因子在植物细胞抗氧化中正调控。Yadav等[39]研究表明,拟南芥MYC 2转录因子能结合Z-box与G-box光应答结构,调节光调控基因的表达。Gangappa等[10]验证了双突变拟南芥中MYC 2能结合SPA1 (SUPPRESSOR OF PHYTOCHROME1)启动子抑制光形态建成。硫氧还蛋白(TRX)是一种在植物体高度保守的低分子量酸性蛋白,能够清除细胞内活性氧(Reactive oxygen species, ROS),从而对抗逆基因进行代谢调控[11]。 Laloi等[40]研究发现,TRX h过表达会促进拟南芥CAT的活性升高,以抵御逆境胁迫。Broin等[41]研究表明,与TRX活性中心类似的CDSP32是植物抵御光合作用膜脂过氧化过程的重要组成部分。另外,TRX h还通过二硫键的氧化还原反应调节蛋白质结构导致其变性[42-43]。通过对MYC2和TRX在鄞红葡萄中基因表达量测定中发现,随着弱光胁迫程度的加强,二者表达量均先升高后降低,与抗氧化酶活性的研究结果大致相同。这可能是因为植株在适度弱光下,通过MYC2和TRX表达上调来清理体内多余的ROS;其中,TRX h与可溶性蛋白的变化也基本一致,进一步证明了植物在胁迫环境下会通过合成与分解可溶性蛋白减少细胞受损。但高度弱光胁迫下细胞受损会导致基因调控失衡,这与前人对拟南芥的研究结果基本一致[38]。
由于宁波地区夏季光照充足、温度恒定,本研究证明适度遮阴较有利于鄞红葡萄的生长,适度弱光下细胞保护酶活性与渗透调节物质的合成状况良好。今后需验证更多与鄞红有关的应答基因,以确认该基因是否受到弱光胁迫的进一步诱导。另外,在弱光胁迫的基础上,可以加入高温、高湿、重金属胁迫等双重或多重胁迫因子,结果可为多抗葡萄品种的选育提供理论支持。
