黑穗醋栗果实生长发育过程中抗坏血酸含量及相关酶活性的变化
2019-01-09孙小娟刘庆帅员盎然张妍霍俊伟秦栋姜婷
孙小娟,刘庆帅,员盎然,张妍,霍俊伟,秦栋,姜婷
黑穗醋栗果实生长发育过程中抗坏血酸含量及相关酶活性的变化
孙小娟,刘庆帅,员盎然,张妍,霍俊伟,秦栋,姜婷
(东北农业大学园艺园林学院/寒地小浆果开发利用国家地方联合工程研究中心,哈尔滨 150030)
【目的】研究不同品种不同生长时期黑穗醋栗果实内抗坏血酸(AsA)含量及其合成代谢过程中相关酶活性的变化,以明确果实生长发育过程中AsA含量与代谢合成相关酶之间的关系,为全面揭示黑穗醋栗果实AsA积累规律提供理论依据。【方法】以3个不同黑穗醋栗品种(‘亚德’‘布劳德’和‘黑丰’)为试材,测定果实在幼果期、膨大期、半转色期、转色期和成熟期时还原型抗坏血酸(AsA)、氧化型抗坏血酸(DHA)、还原型谷胱甘肽(GSH)和氧化型谷胱甘肽(GSSG)含量以及AsA合成与代谢相关酶活性。【结果】不同品种黑穗醋栗果实大小、AsA含量以及AsA相关代谢物水平存在明显的多样性。其中‘亚德’单果重最大,为1.97 g。果实生长发育过程中,总抗坏血酸(T-AsA)和AsA含量在3个品种中变化趋势一致,均在幼果期含量最高,其中‘亚德’幼果期果实中AsA含量最高,为83.17 μmol·g-1FW,随着果实的生长迅速下降,在成熟期降至21.28 μmol·g-1FW;3个品种果实中GSH和T-GSH含量随着果实发育呈升高趋势,但不同品种升高时期或增加幅度不同;GSSG含量在不同品种间存在较大差异,成熟果中‘黑丰’含量最低,为0.008 μmol·g-1FW,仅为‘亚德’的10.2%。AsA-GSH循环再生代谢中,脱氢抗坏血酸还原酶(DHAR)和单脱氢抗坏血酸还原酶(MDHAR)活性在果实膨大期达到最高,成熟期降至最低,其中‘布劳德’果实中DHAR和MDHAR活性略高于‘亚德’和‘黑丰’;谷胱甘肽还原酶(GR)活性在幼果期最高,‘亚德’幼果期果实中GR活性最高(0.06 μmol·min-1·g-1FW),之后随着果实的生长发育不断下降,抗坏血酸过氧化物酶(APX)活性变化与之相似;L-半乳糖途径的关键酶L-半乳糖-1,4-内酯脱氢酶(GalLDH)活性随着果实生长发育的变化趋势与AsA含量变化相一致,且‘亚德’果实中GalLDH活性在幼果期和成熟期均高于其他两个品种。通过相关性分析发现,GalLDH与T-AsA、AsA、DHA、DHAR和MDHAR呈现极显著正相关关系,相关系数可达0.91以上,即果实中GalLDH活性越高,果实中AsA含量也越高;DHAR和MDHAR与T-AsA、AsA间也存在极显著正相关关系,而APX与T-GSH、GSH间相关性较强。【结论】黑穗醋栗幼果期果实中AsA含量最高,且品种间差异显著;GalLDH、MDHAR和DHAR可能是黑穗醋栗果实中AsA合成代谢的关键酶,黑穗醋栗果实中AsA含量积累主要取决于GalLDH活性,说明合成途径起着更关键的作用,而AsA-GSH循环再生途径相关酶对AsA合成也有一定的贡献,黑穗醋栗高AsA含量的积累是由合成途径与循环途径共同作用的结果。
黑穗醋栗;果实;抗坏血酸;合成;L-半乳糖-1,4-内酯脱氢酶
0 引言
【研究意义】黑穗醋栗(L.)属虎耳草科(Saxifragaceae)茶藨子属()落叶丛生灌木[1],果实为黑色浆果,近球形,味酸甜,营养价值丰富[2-3],是极具开发价值的寒地小浆果[4]。黑穗醋栗以富含高水平的抗坏血酸(Ascorbic acid,AsA)而著称,鲜果中AsA含量高达11.36—22.73 μmol∙g-1FW[5],仅次于猕猴桃,是重要的AsA植物源。AsA含量的高低是衡量与评价果实品质高低的重要指标之一,研究黑穗醋栗果实在整个发育过程中AsA积累量与关键酶活性的变化,对阐明黑穗醋栗AsA的合成与积累机制及培育高AsA含量的品种具有重要意义。【前人研究进展】几乎所有的新鲜蔬菜和水果都含有AsA[6-7]。AsA作为抗氧化剂和酶辅助因子在植物的多数生物过程中起着至关重要的作用[8],在维持人体健康,降低慢性疾病风险[9-11],促进胶原蛋白形成和骨骼发育等方面也有重要的医疗价值[12-13],因此,AsA在植物中的合成与代谢备受重视。植物中AsA的合成途径主要有4条,其中刺梨[14]、猕猴桃[15]和苹果[16]等大多数植物的主要合成途径为L-半乳糖途径,L-半乳糖-1,4-内酯脱氢酶(GalLDH)是催化AsA合成的最后一步关键酶[8]。抗坏血酸-谷胱甘肽(AsA–GSH)循坏再生途径对AsA含量也有一定影响[17],植物体内AsA最终含量受合成和循环再生的调控[18]。脐橙[19]果皮中AsA含量积累主要取决于GalLDH活性,而果肉中AsA含量可能取决于AsA的再生循环系统。已有研究表明黑穗醋栗果实中AsA积累主要是通过原位生物合成[20],黑穗醋栗不同品种成熟期果实AsA含量不同,差异明显,并且对其植株进行摘叶摘果处理后发现,果实中AsA含量变化幅度较小[21]。【本研究切入点】目前,对猕猴桃[22]、苹果[23]、甜樱桃[24]和刺梨[25]等果实中AsA含量以及相关的研究已经较为深入。而有关黑穗醋栗AsA含量研究仅限于测定不同品种果实中的总含量[20,26],对黑穗醋栗不同品种各个发育阶段果实中AsA含量水平,以及影响不同品种AsA合成积累的关键酶活性还未有过系统分析,对其AsA合成再生途径及调控AsA代谢机理的研究还不清楚。【拟解决的关键问题】笔者课题组前期研究表明,不同品种黑穗醋栗成熟期果实AsA含量及AsA合成相关酶活性差异较大[26]。为探明差异的原因,进一步研究黑穗醋栗果实内AsA的代谢机理,本试验选取3个不同品种的黑穗醋栗果实,对不同生长发育期果实内的AsA含量及相关酶活性进行测定,通过分析AsA合成及循环再生代谢的相关酶活性,研究黑穗醋栗AsA积累的机理,为培育更优质的品种奠定基础。
1 材料与方法
试验于2016—2017年在黑龙江省哈尔滨市东北农业大学黑穗醋栗种质资源圃进行。
1.1 试验材料
供试材料为3个不同黑穗醋栗主栽品种‘亚德列娜亚(以下均简称亚德)’(Adelinia)、‘布劳德’(Brodtrop)和‘黑丰’(Heifeng),10年生,株行距为1.2 m×1.5 m,常规管理。
根据果实的大小和颜色(图1),在幼果期、膨大期、半转色期、转色期和成熟期采样,采样时间为2016年5—7月和2017年5—7月,选取2016和2017年的数据均值分析。测试样品每3棵一次重复,3次重复,每次从不同单株的不同部位均匀采集≥30个果实,新鲜果实采后迅速放入冰盒,带回试验室计数、游标卡尺测量及称重后用刀切碎混合,立即用液氮速冻,分装保存于-80℃冰箱,用于测定AsA合成代谢相关酶活性。

A:亚德Adelinia;B:布劳德Brodtrop;C:黑丰Heifeng。下同 The same as below
1.2 果实单果重及横纵径测定
单果鲜重:用电子天平称取,结果精确到化0.01 g。
果实横纵径:采用游标卡尺测量,结果精确到0.01 mm。以上单果鲜重和果实横纵径全部观测完成后,根据每次观测记录的平均值,绘制果实生长曲线。
1.3 测定项目及方法
还原型抗坏血酸(AsA)、氧化型抗坏血酸(DHA)、还原型谷胱甘肽(GSH)和氧化型谷胱甘肽(GSSG)含量的测定试剂盒及脱氢抗坏血酸还原酶(DHAR)、单脱氢抗坏血酸还原酶(MDHAR)、谷胱甘肽还原酶(GR)、抗坏血酸过氧化物酶(APX)和L-半乳糖-1,4-内酯脱氢酶(GalLDH)活性的试剂盒购于苏州科铭生物技术有限公司,按照说明书进行提取、测定。其中总抗坏血酸(T-AsA)含量为AsA与DHA之和,总谷胱甘肽(T-GSH)含量为GSH与GSSG之和。以上测定均重复3次。
1.4 数据处理
用Excel 2007进行数据处理、作图,以SPSS 17.0软件进行相关分析。
2 结果
2.1 黑穗醋栗果实生长发育动态
随着果实的发育,3个品种果实单果重和横、纵径均呈明显的上升趋势,其中膨大期上升幅度最大(图2)。3个品种成熟果中,以‘亚德’的果实最大,单果重、横径、纵径均最大,分别达到1.97 g、14.28 mm、15.17 mm,‘布劳德’次之,‘黑丰’果实最小,分别为0.98 g、12.03 mm、12.72 mm,‘黑丰’单果重仅为‘亚德’的50.25%。
2.2 黑穗醋栗果实生长发育过程中AsA、DHA及T-AsA含量的变化
3个品种果实中T-AsA、AsA和DHA含量在整个果实生长发育中变化相似,均呈下降趋势(图3),其中在果实的幼果期至膨大期,果实AsA含量下降较快,‘亚德’‘布劳德’和‘黑丰’分别下降57.11%、26.95%和33.31%,而3个品种果实成熟期的AsA含量分别为幼果期的25.59%、37.67%和33.62%。‘亚德’成熟期果实中AsA含量最高,为21.28 μmol∙g-1FW,分别是‘布劳德’和‘黑丰’的1.39倍和2.18倍,这进一步证实了黑穗醋栗品种果实AsA含量具有多样性。3个品种的黑穗醋栗果实生长过程中抗坏血酸主要以还原态的AsA的形式存在,‘亚德’成熟果中DHA含量只达到AsA的21.38%。

同一品种内中不同字母表示差异显著(<0.05)。下同
The different letters in the same cultivar showed significant difference (<0.05). The same as below
图2 黑穗醋栗果实单果重及横纵径变化
Fig. 2 Changes of single fruit weight, horizontal diameter and vertical diameter of black currant
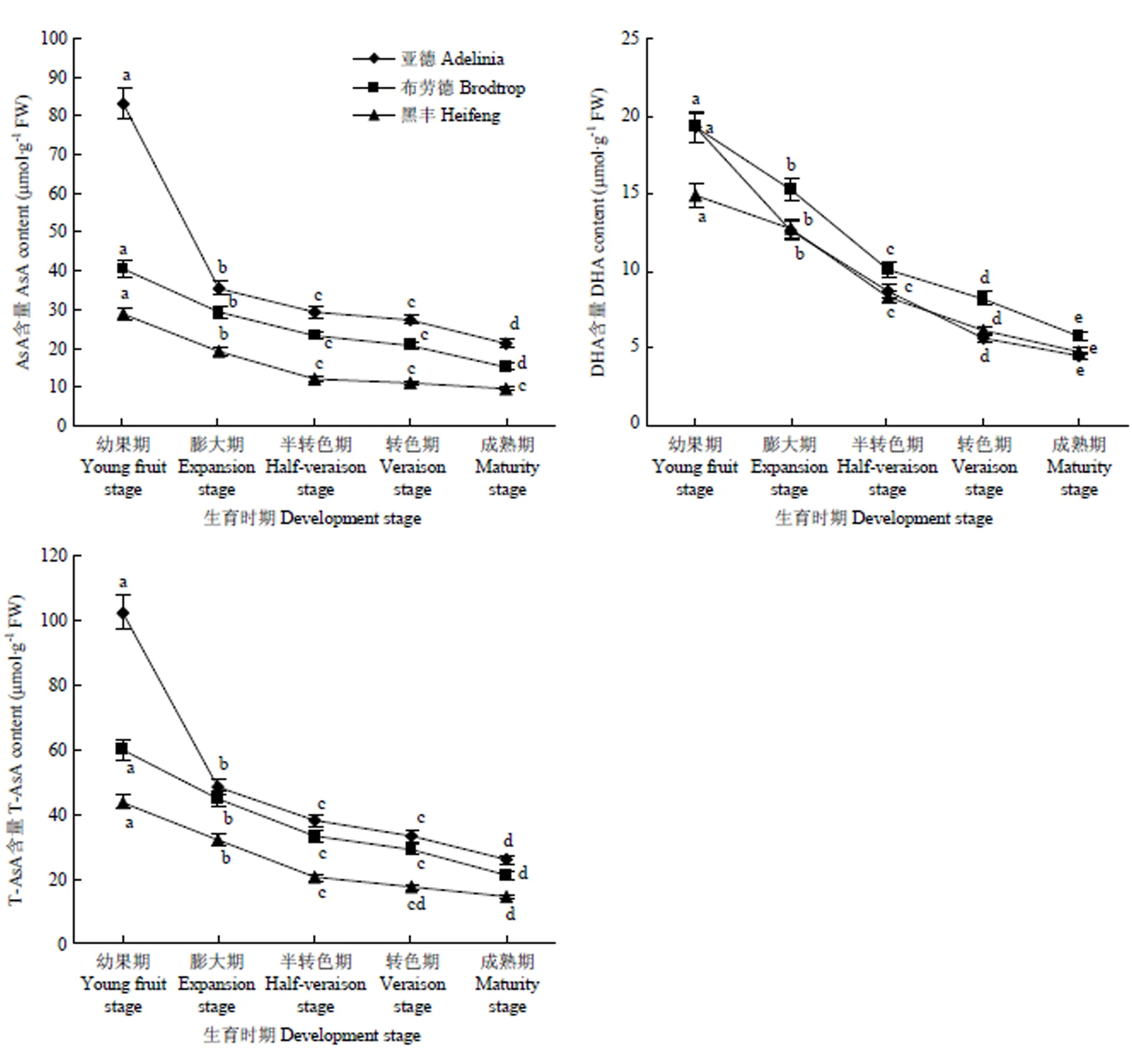
图3 黑穗醋栗果实生长发育过程中T-AsA、AsA和DHA含量的变化
2.3 黑穗醋栗果实生长发育过程中GSH、GSSG及T-GSH含量的变化
3个品种GSH和T-GSH含量在不同生长发育时期表现出明显的差异,随着果实发育呈升高趋势,但不同品种升高时期或增加幅度不同(图4)。‘亚德’中GSH和T-GSH含量变化呈现先上升后下降再上升的变化趋势,均在果实幼果期处于最低水平,分别为0.10和0.19 μmol∙g-1FW,随着果实进入膨大期,GSH和T-GSH含量急剧增加,并在果实半转色期达到峰值,分别为幼果期的3.37倍和5.3倍,之后在转色期略有下降,而在果实成熟期回升至与半转色期基本持平。‘布劳德’和‘黑丰’中GSH和T-GSH含量变化模式相似,总体上均呈“M”型变化趋势。果实GSH和T-GSH含量也是在幼果期最低,不同的是在半转色期含量较低,而在转色期快速增加至最大值(图4-A、C)。
3个品种GSSG含量变化趋势有所差异,其中‘亚德’和‘布劳德’呈先上升后下降的趋势,而‘黑丰’GSSG含量一直处于下降。‘亚德’和‘布劳德’成熟果中GSSG含量相差不大,分别为0.078和0.071 μmol∙g-1FW,而‘黑丰’成熟果中含量较低,为0.008 μmol∙g-1FW,仅为‘亚德’的10.2%(图4-B)。
2.4 黑穗醋栗果实生长发育过程中AsA-GSH氧化还原程度
AsA/DHA的大小代表了AsA氧化还原程度的高低,比值越高,则还原态的AsA含量越高,对AsA的积累越有利。3个品种AsA/DHA值随着果实的发育,均呈先下降后上升的趋势(图5),其中‘亚德’果实中AsA/DHA值较高,说明‘亚德’中AsA氧化程度较高。‘亚德’和‘布劳德’果实生长发育前期,AsA/DHA逐渐降低,均在果实膨大期降至最低,分别为2.82、1.93,在果实半转色期和转色期持续增加,在果实转色期达到最大值,分别为果实膨大期的1.71和1.35倍,之后在果实成熟期缓慢降低,但品种内差异不显著(图5)。而‘黑丰’果实中的AsA/DHA比值在果实半转色期降低至最低值(1.45),果实成熟期的AsA/DHA值为果实半转色期的1.39倍。

图4 黑穗醋栗果实生长发育过程中T-GSH、GSH和GSSG含量的变化
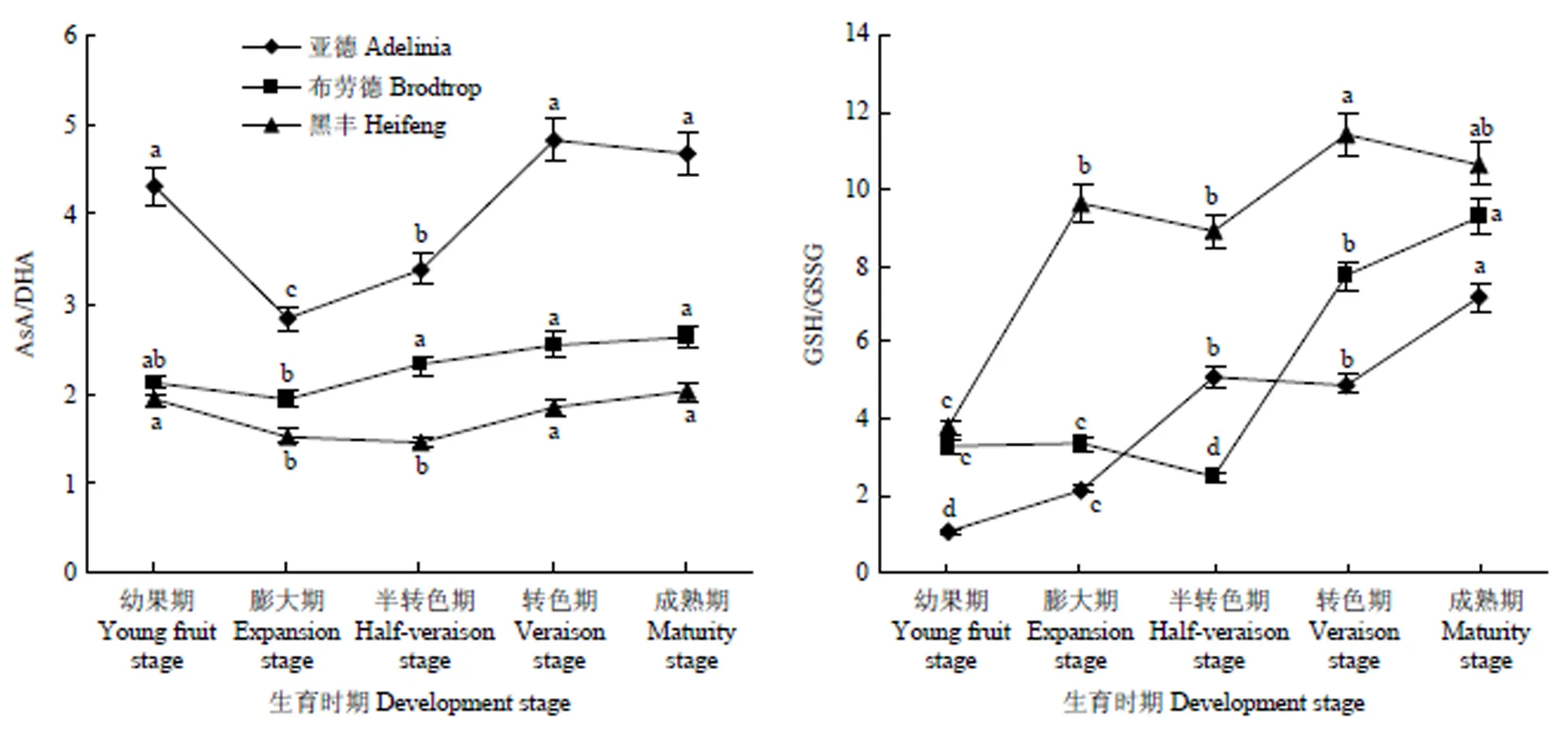
图5 黑穗醋栗果实生长发育过程中AsA/DHA和GSH/GSSH值的变化
GSH/GSSG值用来衡量谷胱甘肽氧化还原状态,通常使用GSH/GSSG值来反应AsA-GSH循环在AsA生成中所起的重要作用。3个品种GSH/GSSG值变化趋势有所不同(图5),随着果实的发育,在‘亚德’和‘黑丰’中总体呈上升趋势,而‘布劳德’在果实半转色期有一个明显的下降趋势。在‘亚德’和‘黑丰’果实的整个发育时期,GSH/GSSG值均在幼果期最低,之后持续提高,说明此时GSH含量呈上升趋势,再生能力增强,合成能力有所减缓,在果实成熟期达到最大,分别为果实幼果期的6.54倍和2.81倍。‘布劳德’中GSH/GSSG值在半转色期最低,之后迅速上升,至成熟期达到9.27。
2.5 黑穗醋栗生长发育过程中AsA-GSH循坏代谢相关酶活性的变化
3个品种APX活性变化趋势基本一致,均在幼果期达到峰值,此时‘亚德’‘布劳德’和‘黑丰’APX活性分别为0.85、0.63和0.56 μmol∙min-1∙g-1FW (图6-A),到果实半转色期后3个品种中APX活性分别降至果实幼果期的23.73%、25.24%和23.43%,APX活性的降低有助于确保AsA含量的积累与稳定。从品种间来看,‘亚德’果实中APX活性较高,其他两个品种相差不大。
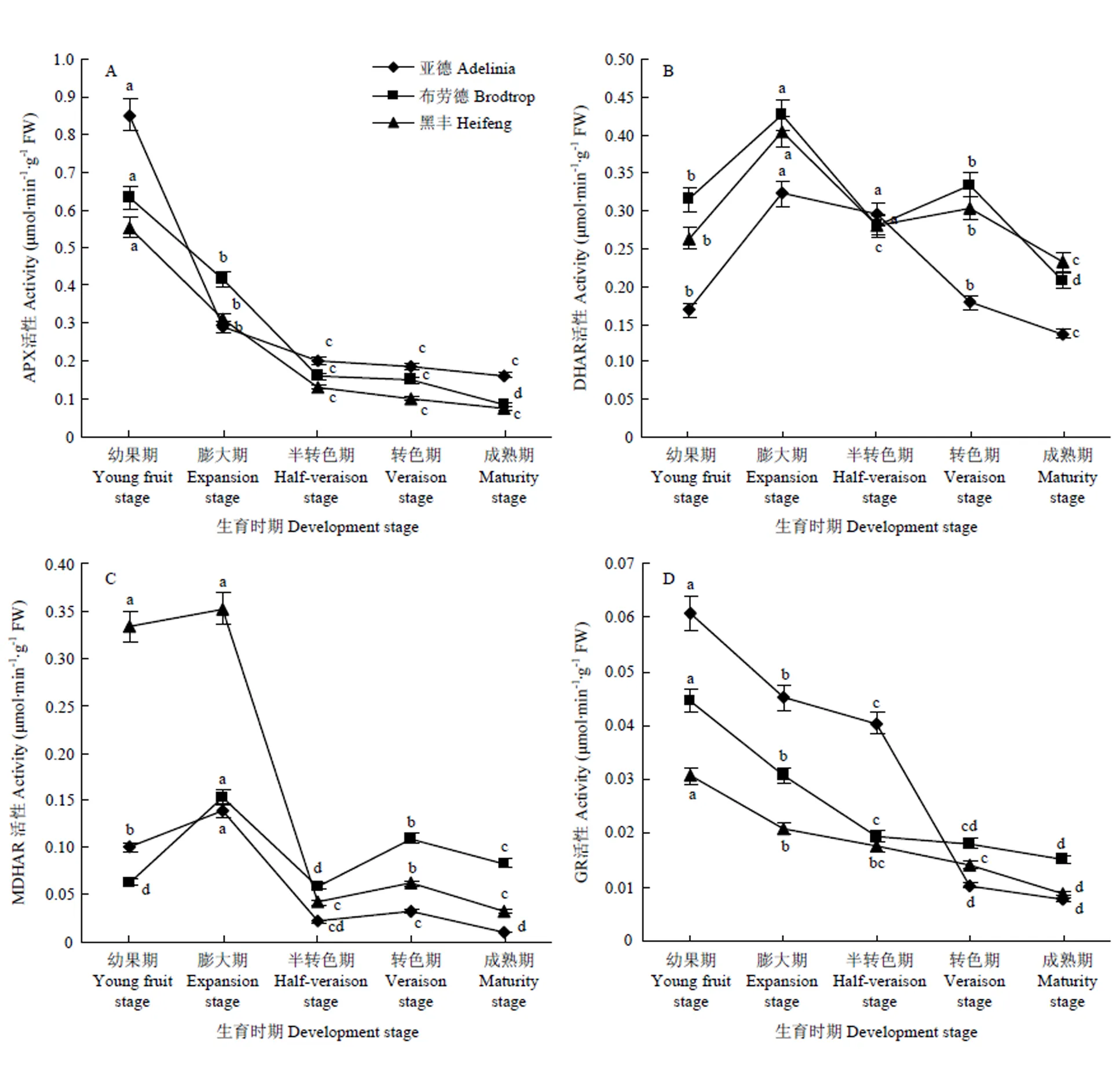
图6 黑穗醋栗果实生长发育过程中AsA-GSH循环酶活性的变化
3个品种果实生长发育过程中DHAR和MDHAR活性总体变化趋势基本一致,均呈“M”型的变化(图6-B、C),即先增加后降低再增加再降低的形式。3个品种果实DHAR活性均在果实膨大期最高,‘亚德’‘布劳德’和‘黑丰’分别为0.32、0.43和0.40 μmol∙min-1∙g-1FW,之后随着果实的生长而下降,‘布劳德’和‘黑丰’在转色期有轻微的回升,随后均随着果实的成熟降至最低,成熟期酶活性分别比膨大期下降了56.25%、51.16%和42.50%。同时,3个品种果实MDHAR活性均在膨大期达到峰值,分别为0.14、0.15和0.35 μmol∙min-1∙g-1FW,在果实成熟期降到最低值,仅分别为果实膨大期的6.93%、54.22%和9.11%。相较于DHAR活性大小,不同品种不同时期果实中MDHAR活性均明显低于DHAR活性。
3个品种果实生长发育过程中GR活性均呈下降趋势(图6-D),都表现为幼果期酶活性最高,之后逐渐下降,并于果实成熟期时降至最低值,此时‘亚德’‘布劳德’和‘黑丰’GR活性分别下降至果实幼果期的12.79%、34.31%和28.92%,‘亚德’GR活性下降幅度最大。同时,也发现果实中GR活性的变化趋势与APX的变化趋势相似。AsA-GSH循坏代谢相关酶活性均在果实幼果期相对较高,说明在黑穗醋栗果实生长发育初期再生循环对AsA含量积累的影响较大。
2.6 黑穗醋栗果实生长发育过程中GalLDH活性变化
3个品种果实生长发育过程中GalLDH活性存在明显的差异(图7),其中,‘亚德’幼果期GalLDH活性最高。GalLDH的活性变化趋势在‘亚德’和‘布劳德’中较为相似,均是在幼果期活性相对较高,分别为4.07和2.69 μmol∙min-1∙g-1FW,这也是果实内AsA含量迅速积累的时期,随着果实的发育,在果实膨大期迅速下降,之后表现为缓慢下降,并在果实成熟期降到最低,分别为幼果期的38.1%和35.83%,且GalLDH活性变化趋势与果实生长发育过程中AsA含量的变化趋势相一致,这说明GalLDH在黑穗醋栗果实AsA合成途径中起着关键作用。
‘黑丰’果实生长发育过程中,GalLDH活性呈 先增加后下降的变化趋势,在果实进入膨大期后,GalLDH活性开始迅速增加,至半转色期达到最大值,而在转色期又迅速下降,至成熟期降至最低,为1.45 μmol∙min-1∙g-1FW,这与另外两个品种果实生长发育过程中AsA含量的变化趋势存在较大差异。从品种间来看,‘亚德’GalLDH活性在幼果期和成熟期均高于其他两个品种。

图7 黑穗醋栗果实生长发育过程中GalLDH活性的变化
2.7 黑穗醋栗果实生长发育过程中AsA合成代谢相关指标间的相关性
通过对果实(以‘亚德’为例,另外两个品种与‘亚德’类似)不同生长发育时期相关酶活性和果实中AsA含量进行相关性分析发现(表1),在‘亚德’果实生长发育过程中,AsA和GSH的氧化还原状态显示出显著的一致性,AsA/DHA和GSH/GSSG的相关系数为0.918。T-AsA、AsA和DHA含量与GalLDH、MDHAR和DHAR活性分别呈极显著正相关,相关系数达0.91以上,与GSSG含量显著正相关,相关系数达0.71以上。GSSG含量与MDHAR酶活性呈显著正相关,GSH/GSSG比值与MDHAR和GR活性呈显著负相关。APX与GR之间呈现出极显著的正相关性,相关系数为0.929,与AsA/DHA、T-GSH、GSH、GSSG和GSH/GSSG呈极显著负相关,与MDHAR呈负相关。MDHAR和DHAR两者之间极显著正相关。可见,AsA合成途径中的GalLDH,AsA-GSH循环途径中的MDHAR和DHAR对黑穗醋栗果实AsA含量起了关键作用,其中GalLDH是AsA合成关键酶。而APX对果实中的GSH含量有较大的影响。

表1 ‘亚德’果实生长发育过程中AsA合成代谢相关酶的相关系数
*表示显著水平0.05;**表示显著水平0.01 * denote significant at 0.05; ** denote significant at 0.01
3 讨论
3.1 黑穗醋栗AsA的积累变化
AsA是植物生长发育所必需的物质,不同物种间其含量差异很大,即使同一物种不同品种间AsA含量也存在差异,这在猕猴桃[18]和油梨[27]上已证明。本研究中发现‘亚德’成熟果实中AsA含量最高,分别是‘布劳德’和‘黑丰’的1.39倍和2.18倍。黑穗醋栗果实中AsA的含量随着果实发育呈相反的变化,在幼果期AsA含量最高,随着果实的膨大有一个明显的下降趋势,这可能是由于果实细胞处于快速生长期,AsA参与细胞分裂引起的,也可能是这个时期的细胞膨大对AsA含量起到稀释作用[25],这种变化与树莓[28]、桃[29]、西印度樱桃[30]等果实中AsA积累一致,但与刺梨[14]果实AsA的积累不同,其在发育后期才开始大量积累AsA,这也反映了不同物种间的差异性。
3.2 黑穗醋栗AsA合成代谢相关酶活性的变化特征
GalLDH是AsA合成关键酶,在L-半乳糖途径中直接氧化AsA合成前体L-半乳糖-1,4-内酯,且对底物具有较强的特异性[18]。对甜樱桃[24]、苹果[31]和马铃薯[32]等研究中发现GalLDH在AsA合成途径中起到主导作用,同时对烟草过表达GalLDH发现AsA含量显著提高[33]。本研究结果显示,‘亚德’和‘布劳德’在整个果实发育时期,GalLDH活性变化趋势与AsA含量变化基本一致,而且‘亚德’果实GalLDH活性明显高于其他两个品种,这也进一步证明GalLDH活性的高低可能是影响黑穗醋栗品种间果实AsA含量差异的重要原因,同时也解释了‘亚德’AsA含量高的原因,进而推测GalLDH可能在AsA合成过程中起着关键作用。而‘黑丰’果实中GalLDH活性变化与AsA含量变化存在差异,可能与AsA-GSH循环再生途径相关酶的代谢调控相关,也可能与品种有关。
植物中AsA含量主要受合成途径的影响,但再生途径对AsA含量也有重要影响,其中AsA-GSH循环系统是植物中AsA再生的主要途径[34]。本研究中发现,黑穗醋栗果实发育前期APX活性明显高于其他酶活性,随后在果实膨大期开始迅速降低,这与AsA/DHA值之间呈现出显著的负相关,推测原因可能是果实发育前期细胞分裂活动比较旺盛,细胞内产生大量的H2O2,此时APX以AsA为电子供体清除细胞内的H2O2,这样可有效防止活性氧对细胞的毒害,保护细胞免受氧化损伤[35]。随后随着果实的发育缓慢下降,在半转色期后其酶活性基本维持在较低的水平,这表明在这一时间段内果实内积累的AsA极少被氧化,从而确保果实体内AsA的积累与稳定。而作为电子供体的AsA通过AsA-GSH循环,经MDHAR和DHAR作用又可以重新生成AsA。在本试验中,DHAR和MDHAR活性与AsA含量变化趋势基本一致,在果实发育过程中总体均呈下降的趋势,说明DHAR和MDHAR活性是影响黑穗醋栗果实AsA含量的决定因素之一。此外物种的不同,AsA的再生过程中起关键作用的酶则不同,蓝莓[36]、苹果[37]、番茄[38]等研究表明果实中MDHAR在对AsA循环上起更主要的作用,但在刺梨果实中却发现DHAR的表达和活性变化均要高于MDHAR[14]。而在本研究中发现,AsA含量较低的‘黑丰’其MDHAR活性在整个果实发育期内均高于‘亚德’,而AsA含量高的‘亚德’其DHAR活性在整个果实发育阶段均低于‘布劳德’和‘黑丰’,至于黑穗醋栗果实内DHAR和MDHAR哪个酶活性更能起决定,仍需进一步探究。随着黑穗醋栗果实的成熟,果实GR活性下降,而AsA含量也下降,表明GR活性有利于果实中AsA的积累。
3.3 黑穗醋栗AsA含量与合成代谢相关酶的关系
在越橘[39]、草莓[40]、甜樱桃[24]、猕猴桃[41]等研究中发现,DHAR和MDHAR在AsA-GSH循环中起到关键性作用。本研究结果表明,‘亚德’果实中DHAR和MDHAR活性与AsA含量呈显著正相关,说明黑穗醋栗果实中AsA的再生主要依赖DHAR和MDHAR维持,同时发现DHAR和MDHAR呈显著正相关,说明AsA循环再生酶可能随着一种酶活性的变化而相应变化。近年来通过MDHAR和DHAR的表达来提高AsA含量已有不少成功的例子。如在烟草植株[42]中超表达拟南芥叶绿体的MDHAR后,烟草叶片中AsA含量上升2.2倍。而将小麦胞质DHAR分别在烟草和玉米中过量表达后,结果发现烟草和玉米中DHAR活性均增加了32倍,其AsA含量也增加了4倍,不仅对AsA含量的影响显著,而且GSH含量与GSH/GSSG的比率也有所增加,说明由DHAR过量表达引起的AsA含量提高的同时,GSH的合成也伴随着增加,这在马铃薯[43]中同样得到了证实。作为L-半乳糖途径中的关键酶GalLDH,它的有无决定了植物能否自身合成AsA[34]。猕猴桃中AsA含量的积累与GalLDH活性呈显著正相关[41],同样水稻中GalLDH活性与AsA含量也存在正相关关系[44]。本试验3个品种中,GalLDH与AsA、T-GSH、MDHAR和DHAR均有较高的相关性,说明GalLDH是黑穗醋栗果实AsA合成代谢的关键酶。这些结果说明,AsA合成能力在维持AsA含量中起着主导作用,但再生能力同时也具有一定作用,且可能在特定条件和发育阶段对AsA水平起到主要的调控作用。
4 结论
AsA含量在黑穗醋栗不同品种间存在明显差异,但均随着果实的生长发育呈下降趋势。GalLDH、MDHAR和DHAR是黑穗醋栗果实中AsA合成代谢的关键酶,而APX对果实中的GSH含量有较大的影响。黑穗醋栗果实中AsA含量积累主要取决于GalLDH活性,占主导地位,AsA-GSH循环再生途径对AsA合成也有一定的贡献,黑穗醋栗高AsA含量的积累是由合成途径与循环途径共同作用的结果。黑穗醋栗果实中AsA含量是评价和衡量黑穗醋栗果实品质的重要指标,相关研究结果也为黑穗醋栗资源的评价与利用、果实品质改良和新品种选育提供了指导。
[1] 秦栋, 张椿浩, 刘庆帅, 薛晓晓, 霍俊伟, 员盎然. ABA和GA3对黑穗醋栗二次萌发芽超微结构的影响. 果树学报, 2017, 34(10): 1301-1308.
QIN D, ZHANG C H, LIU Q S, XUE X X, HUO J W, YUN A R. Effect of GA3and ABA on cell ultra-structure of the secondary- bursting buds in blackcurrant ()., 2017, 34(10): 1301-1308. (in Chinese)
[2] 李贺, 李歆昕, 陆璐, 柯筱纯, 阮成江. 5种黑穗醋栗果实中糖酸组成与含量分析. 食品工业科技, 2016, 37(5): 137-147.
LI H, LI X X, LU L, KE X C, RUAN C J. Analysis of contents and constituents of sugar and organic acid in 5 black currant cultivars., 2016, 37(5): 137-147. (in Chinese)
[3] 张亚楼. 黑加仑营养成分及保健功能研究进展.环境卫生学杂志, 2004, 31(2): 108-111.
ZHANG Y L. Research progress of nutrients and health function of black currant., 2004, 31(2): 108-111. (in Chinese)
[4] 宋杨, 张春雨, 张志东, 温景辉, 李亚东, 吴林, 刘海广. 黑穗醋栗品种亲缘关系的ISSR分析. 园艺学报, 2011, 38(9): 1747-1752.
SONG Y, ZHANG C Y, ZHANG Z D, WEN J H, LI Y D, WU L, LIU H G. Genetic relationship of blackcurrant cultivars revealed by ISSR markers., 2011, 38(9): 1747-1752. (in Chinese)
[5] 霍俊伟, 李著花, 秦栋. 黑穗醋栗营养成分和保健功能及产业发展前景. 东北农业大学学报, 2011, 42(2): 139-144.
HUO J W, LI Z H, QIN D. Review of nutritional ingredients and health protectal function of black currant fruit and its prospect in industrial development., 2011, 42(2): 139-144. (in Chinese)
[6] 安华明, 陈力耕, 樊卫国, 胡西琴. 高等植物中维生素C的功能、合成及代谢研究进展. 植物学通报, 2004, 21(5): 608-617.
AN H M, CHEN L G, FAN W G, HU X Q. Advances in research on function, biosynthesis and metabolism of ascorbic acid in higher plants., 2004, 21(5): 608-617. (in Chinese)
[7] CRUZRUS E, AMAYA I, SÁNCHEZ-SEVILLA J F, BOTELLA M A, VALPUESTA V. Regulation of L-ascorbic acid content in strawberry fruits., 2011, 62(12): 4191-4201.
[8] SMIRNOFF N, WHEELER G L. Ascorbic acid in plants: biosynthesis and function., 2000, 35(35): 291-314.
[9] DELRIO R, MOYA E A, ITURRIAGA R. Carotid body and cardiorespiratory alterations in intermittent hypoxia: the oxidative link., 2010, 36(1): 143-150.
[10] MISSO N L A, BROOKS-WILDHABER J, RAY S, VALLY H, THOMPSON P J. Plasma concentrations of dietary and nondietary antioxidants are low in severe asthma., 2005, 26(2): 257-264.
[11] FRITZ H, FLOWER G, WEEKS L, COOLEY K, CALLACHAN M, MCGOWAN J, SKIDMORE B, KIRCHNER L, SEELY D. Intravenous vitamin C and cancer: A systematic review., 2014, 13(4): 280-300.
[12] SARKAR N, SRIVASTAVA P K, DUBEY V K. Understanding the language of vitamin C., 2009, 5(1): 53-55.
[13] OLMOS E, KIDDLE G, PELLNY TK, KUMAR S, FOYER C H. Modulation of plant morphology, root architecture, and cell structure by low vitamin C in., 2006, 57(8): 1645-1655.
[14] HUANG M, XU Q, DENG X X. L-ascorbic acid metabolism during fruit development in an ascorbate-rich fruit crop chestnut rose (Tratt)., 2014, 171(14): 1205-1216.
[15] BULLEY S M, RASSAM M, HOSER D, OTTO W, SCHUNEMANN N, WRIGHT M, MACRAE E, GLEAVE A, LAING W. Gene expression studies in kiwifruit and gene over-expression inindicates that GDP-L-galactose guanyltransferase is a major control point of vitamin C biosynthesis., 2009, 60(3): 765-778.
[16] LI M J, MA F W, ZHANG M, PU F. Distribution and metabolism of ascorbic acid in apple fruits (Borkh cv.Gala.)., 2008, 174(6): 606-612.
[17] 郑俊鶱, 孙艳, 韩寿坤, 张浩, 王益权.土壤紧实胁迫对黄瓜抗坏血酸-谷胱甘肽循环的影响. 中国农业科学, 2013, 46(2): 433-440.
ZHENG J S, SUN Y, HAN S K, ZHANG H, WANG Y Q. Effect of soil compaction stress on ascorbate-gluthione., 2013, 46(2): 433-440. (in Chinese)
[18] 原玉林, 同晓蕾, 侯长明, 马锋旺, 李明军. 不同基因型猕猴桃果实中抗坏血酸合成与代谢的差异. 植物生理学报, 2016, 52(12): 1877-1883.
YUAN Y L, TONG X L, HOU C M, MA F W, LI M J. Difference of ascorbic acid synthesis and metabolism in different genotypes of kiwifruit., 2016, 52(12): 1877-1883. (in Chinese)
[19] 黄艳花, 曾明, 王玲利, 苏芳芳, 李兴发. 晚熟脐橙果实发育过程中抗坏血酸含量及相关酶活性的变化. 植物生理学报, 2014, 50(6): 875-879.
HUANG Y H, ZENG M, WANG L L, SU F F, LI X F. Changes in ascorbic acid contents and related enzyme activities during fruit development of late-maturing navel orange., 2014, 50(6): 875-879. (in Chinese)
[20] HANCOCK R D, WALKER P G, PONT S D, MARQUIS N, VIVERA S, GORDON S L, BRENNAN R M, VIOLA R. L-ascorbic acid accumulation in fruit ofoccurs by in situ biosynthesis via the L-galactose pathway., 2007, 34(12): 1080-1091.
[21] QIN D, ZHAO L J, GAO Y, LI F X, LI S L, HUO J W, LOU S, LIU P. Effects of thinning on ascorbate-glutathione cycle metabolism in black currants (L.)., 2017, 28(5): 903-908.
[22] 侯长明, 李明军, 马锋旺, 梁东, 杜国荣. 猕猴桃果实发育过程中AsA代谢产物积累及相关酶活性的变化. 园艺学报, 2009, 36(9): 1269-1276.
HOU C M, LI M J, MA F W, LIANG D, DU G R. Changes of product accumulation and related enzyme activities in AsA metabolism during kiwi fruit growth and development., 2009, 36(9): 1269-1276. (in Chinese)
[23] 李明军. 苹果和猕猴桃抗坏血酸形成与积累的生理和分子机理研究[D]. 杨凌: 西北农林科技大学, 2009.
LI M J. Physiological and molecular mechanisms of ascorbic acid formation and accumulation in apple and kiwifruit [D]. Yangling: Northwest A&F University, 2009. (in Chinese)
[24] 夏惠, 林玲, 高帆, 倪知游, 高丽扬, 吕秀兰, 梁东. 甜樱桃‘佐藤锦’果实生长发育过程AsA含量及其相关酶活性的变化. 西北植物学报, 2016, 36(10): 2008-2014.
XIA H, LIN L, GAO F, NI Z Y, GAO L Y, LV X L, LIANG D. Changes of AsA content and related enzyme activities in sweet cherry ‘Satonishiki’ during fruit development., 2016, 36(10): 2008-2014. (in Chinese)
[25] 黄明. 刺梨高含量抗坏血酸积累的分子机理研究[D]. 武汉: 华中农业大学, 2013.
HUANG M. Molecular mechanism for the accumulation of high content of L-ascorbic acid in chestnut rose (Tratt) [D]. Wuhan: Huazhong Agricultural University, 2013. (in Chinese)
[26] 李芳晓, 秦栋, 李曙雷, 战歌, 霍俊伟, 王欢欢, 赵毅, 韩伟, 步鹏志. 黑穗醋栗AsA含量及其代谢酶活性差异分析. 南方农业学报, 2014, 45(7): 1237-1241.
LI F X, QIN D, LI S L, ZHAN G, HUO J W, WANG H H, ZHAO Y, HAN W, BU P Z. Ascorbic acid content and difference of related enzyme activities in fruit of blackcurrant., 2014, 45(7): 1237-1241.(in Chinese)
[27] 魏永赞, 王一承, 刘丽琴, 舒波, 谢江辉, 李伟才, 石胜友. 12份引进油梨品种果肉有机酸组分和抗坏血酸含量分析.果树学报, 2018, 35(2): 177-184.
WEI Y Z, WANG Y C, LIU L Q, SHU B, XIE J H, LI W C, SHI S Y. Analysis on the composition and content of organic acids and vitamin C in avocado flesh of twelve introduced cultivars., 2018, 35(2): 177-184. (in Chinese)
[28] 王学勇, 张均营. 树莓和黑莓的研究进展. 安徽农业科学, 2010, 38(10): 5070-5073.
WANG X Y, ZHANG J Y. Research progress on raspberry and blackberry., 2010, 38(10): 5070-5073. (in Chinese)
[29] IMAI T, BAN Y, TERAKAMI S, YAMAMOTO T, MORIGUCHI T. L-Ascorbate biosynthesis in peach: cloning of six L-galactose pathway-related genes and their expression during peach fruit development., 2009, 136(2): 139-149.
[30] BADEJO A A, FUJIKAWA Y. Gene expression of ascorbic acid biosynthesis related enzymes of the Smirnoff-Wheeler pathway in acerola ()., 2009, 166(6): 652-660.
[31] 李明军, 高静, 马锋旺, 梁东, 侯长明. 苹果果实GalDH和GalLDH基因的表达与AsA的关系. 中国农业科学, 2010, 43(2): 351-357.
LI M J, GAO J, MA F W, LIANG D, HOU C M. Relationship between expressions of GalDH and GalLDH and ascorbate content in apple fruits., 2010, 43(2): 351-357. (in Chinese)
[32] 秦爱国, 于贤昌. 马铃薯抗坏血酸含量及其代谢相关酶活性关系的研究. 园艺学报, 2009, 36(9): 1370-1374.
QIN A G, YU X C. Study on ascorbic acid content and its relationship with metabolic enzyme activity in potatoes., 2009, 36(9): 1370-1374. (in Chinese)
[33] LIU W, AN H M, YANG M. Overexpression ofL-galactono-1,4-lactone dehydrogenase in tobacco plant enhances ascorbate accumulation and abiotic stress tolerance., 2013, 35(5): 1617-1624.
[34] 俞乐, 刘拥海, 袁伟超, 周丽萍, 彭长连. 植物抗坏血酸积累及其分子机制的研究进展. 植物学报, 2016, 51(3): 396-410.
YU L, LIU Y H, YUAN W C, ZHOU L P, PENG C L. Recent advances in the study of accumulation of ascorbic acid and its molecular mechanism in plants., 2016, 51(3): 396-410. (in Chinese)
[35] MULLER-MOULE P, CONKLIN P L, NIYOGI K K. Ascorbate deficiency can limit violaxanthin de-epoxidase activity in vivo., 2002, 128(3): 970-977.
[36] LIU F H, WANG L, GU L, ZHAO W, SU H Y, CHENG X H. Higher transcription levels in ascorbic acid biosynthetic and recycling genes were associated with higher ascorbic acid accumulation in blueberry., 2015, 188: 399-405.
[37] LI M, CHEN X, WANG P, MA F. Ascorbic acid accumulation and expression of genes involved in its biosynthesis and recycling in developing apple fruit., 2011, 136(4): 231-238.
[38] STEVENS R, PAGE D, GOUBLE B, GARCHERY C, ZAMIR D, CAUSSE M. Tomato fruit ascorbic acid content is linked with monodehydroascorbate reductase activity and tolerance to chilling stress., 2010, 31(8): 1086-1096.
[39] GIACOMA C, KATJIA K, MARKO S, ANJA H, HELY H, ANNA S, LLARIA M, LAURA J. Ascorbic acid metabolism during bilerry (L.) fruit development., 2012, 169(11): 1059-1065.
[40] 张丙秀, 李柱刚, 高媛, 刘丹, 高庆玉. DHAR与草莓AsA积累关系及DHAR RNAi遗传转化研究. 南方农业学报, 2012, 43(11): 1626-1632.
ZHANG B X, LI Z G, GAO Y, LIU D, GAO Q Y. Relationship between DHAR and AsA accumulation in strawberry and its DHAR RNAi genetic transformation., 2012, 43(11): 1626-1632. (in Chinese)
[41] 吴寒. 毛花猕猴桃果实抗坏血酸合成酶相关基因的克隆及定量表达分析[D]. 南昌: 江西农业大学, 2015.
WU H. Cloning and quantitative analysis of genes related ascorbate biosynthesis in[D]. Nanchang: Jiangxi Agricultural University, 2015. (in Chinese)
[42] YIN L, WANG S W, ELTAYEB A E, UDDIN M I, YAMAMOTO Y, TSUJI W, TAKEUCHI Y, TANAKA K. Overexpression of dehydroascorbate reductase, but not monodehydroascorbate reductase, confers tolerance to aluminum stress in transgenic tobacco., 2010, 231(3): 609-621.
[43] QIN A, SHI Q, YU X. Ascorbic acid contents in transgenic potato plants overexpressing two dehydroascorbate reductase genes., 2011, 38(3): 1557-1566.
[44] LIU Y H, YU L, WANG R Z. Level of ascorbic acid in transgenic rice for L-galactono-1,4-lactone dehydrogenase overexpressing or suppressed is associated with plant growth and seed set., 2011, 33(4): 1353-1363.
The Changes in the Contents of Ascorbic Acid and the Activities of Related Enzymes in Black Currant Fruits during the Process of Its Growth and Development
SUN XiaoJuan, LIU QingShuai, YUN AngRan, ZHANG Yan, HUO JunWei, QIN Dong, JIANG Ting
(School of Horticulture and Landscape Architecture, Northeast Agricultural University/Small Fruit Development and Utilization of National Local Joint Engineering Research Center in Cold Region, Harbin 150030)
【Objective】The changes in the ascorbic acid (AsA) contents and the enzymatic activities during the anabolic process of different cultivars and growth stages of black currant fruits were studied to determine the relationship between AsA contents and anabolic enzymes during the growth and development of fruits, so as to provide a theoretical basis for comprehensively revealing the accumulation rule of AsA in black currant fruits. 【Method】Three different cultivars of black currant fruits (Adelinia, Brodtrop and Heifeng) were studied and determined the contents of reduced AsA, oxidized ascorbic acid (DHA), reduced glutathione (GSH) and oxidized glutathione (GSSG) and anabolic enzymatic activities of young, expansion, half-veraison, veraison and maturity stages. 【Result】There were significant diversities in fruit sizes, AsA contents and AsA metabolites of different cultivars of black currant fruits. The Adelinia had the largest weight of single fruit (1.97 g). During the growth and development process of fruits, the changes in the total ascorbic acid (T-AsA) and AsA contents of fruits were consistent among the three cultivars, and the young fruits had the highest contents. The AsA content of young Adelinia fruit was the highest (83.17 μmol∙g-1FW) and then sharply decreased rapidly to the maturity stage with the growth of the fruit, which decreased to 21.28 μmol∙g-1FW at maturity stage. The contents of GSH and T-GSH in the three cultivars increased with the development of fruits, but the different cultivars increased in different stages and degrees. The content of GSSG was quite different among different cultivars. For the mature fruits, the GSSG content of Heifeng was the lowest, which was 0.008 μmol∙g-1FW and only accounted for 10.2% of Adelinia. In AsA-GSH recycling regeneration metabolism, the activities of dehydroascorbate reductase (DHAR) and monodehydroascorbate reductase (MDHAR) showed the highest level at expansion period, and finally decreased to the lowest level at maturity stage. The DHAR and MDHAR activities of Brodtrop fruits showed slightly higher than those of Adelinia and Heifeng fruits. The activity of glutathione reductase (GR) was the highest level at young stage. The GR activity of Adelinia young fruits was the highest (0.06 μmol∙min-1∙g-1FW), and then decreased with the growth of fruits. The changes in the activities of ascorbate peroxidase (APX) were similar to the changes in the activities of GR. The changes in the activities of L-galactose-1,4-lactone dehydrogenase (GalLDH), a key enzyme of L-galactose pathway, were consistent with the changes in AsA contents. The GalLDH activity of Adelinia young and mature fruits showed higher than that of Heifeng and Brodtrop young and mature fruits, respectively. According to the correlation analysis, the GalLDH activity showed a highly significant positive correlationship with T-AsA, AsA, DHA, DHAR and MDHAR. The correlation coefficient was above 0.91. The higher GalLDH activity was found in the fruits, the higher AsA contents of fruits also was found. There was a highly significant positive correlationship between DHAR and MDHAR, T-AsA and AsA. The APX had a high correlation with T-GSH and GSH. 【Conclusion】 The AsA content of black currant young fruits was the highest and there were significant differences among the three cultivars. The GalLDH, MDHAR and DHAR might be the key enzymes for AsA anabolism in black currant fruits. The accumulation of AsA content of black currant fruits resulted from the activity of GalLDH, which indicated that the anabolic pathway played a more important role and were found to be a dominant position. The related enzymes of AsA-GSH recycling regeneration pathway also contributed to the AsA anabolism. The accumulation of high AsA content in black currant fruits resulted from the combined effects of anabolic and recycling pathways.
L.; fruit; ascorbic acid; anabolism; L-galactose-1, 4-lactone dehydrogenase
10.3864/j.issn.0578-1752.2019.01.010
2018-05-29;
2018-09-19
国家自然科学基金青年科学基金(31201584)、黑龙江省自然科学基金面上项目(C2017015)、黑龙江省博士后启动基金(LBH-Q17029)、东北农业大学“青年才俊”(16QC07)
孙小娟,E-mail:1837770189@qq.com。通信作者霍俊伟,E-mail:junweihuo@aliyun.com。通信作者秦栋,E-mail:dongq9876@126.com
(责任编辑 赵伶俐)
