荒漠植物红砂与西北针茅种群的空间格局及其关联性
2019-01-04夏素娟刘旻霞南笑宁李全弟蒋晓轩
夏素娟,刘旻霞*,朱 恭,南笑宁,李全弟,蒋晓轩
(1 西北师范大学 地理与环境科学学院,兰州 730070;2 兰州市南北两山环境绿化工程指挥部,兰州 730046)
种群空间分布格局是指种群个体在一定水平空间的分布状况,是种群生物学特性、种间关系及环境条件综合作用的结果[1-2],通过对植物种群空间格局的研究,既可以揭示种群在群落中的地位及作用,也可以深刻了解植物种群的生态过程及它们与生境的相互关系[3]。在干旱荒漠和气候变迁等外界因素影响下,皋兰县土壤水质流失严重,种群退化,极大地制约了资源的持续利用以及生态环境的可持续发展。因此,本文从干旱荒漠区优势物种的空间分布格局入手,揭示该地区的生态作用机制,对兰州市皋兰县干旱荒漠区制定有效的资源利用效率、改善水土流失,确保生态可持续发展提供理论基础。
点格局分析的理论是由Ripley在1977年最早提出,此后被进一步完善[4]。该方法通过划定一定面积的研究区域,以植物种群个体的空间坐标为基本数据,分析任意尺度下的种群分布格局及种间关系[5],与传统研究的样方法、分布型指数法、距离法等[6]方法相比,点格局分析法克服了种群空间格局单一尺度变化的缺陷[7-9]。近年来国内许多学者应用点格局分析方法,研究了热带山地雨林[10]、东北长白山[11]、北京东灵山[12]等地区的植物种群分布格局及其关联性,从不同尺度揭示了植物种群分布状况。
红砂(Reaumuriasongarica)为柽柳科多年生小灌木,是中国干旱荒漠区分布最广的植物种之一,董秋莲等通过对张掖市龙首山红砂种群结构和分布格局研究[13]认为红砂是良好的固沙植物,其抗逆性强,被认为是一种荒漠复苏植物[14],西北针茅(Stipasareptanavar.krylovii)种群是典型的草原植物,生态幅度较广。皋兰县地形属黄土高原丘陵沟壑区,红砂和西北针茅是该地区的优势物种,在水土保持,水源涵养及保护物种多样性上发挥着重要作用。目前关于红砂和西北针茅单独的种群、群落结构及生理特征研究较多,关于坡向上对红砂和针茅在同一环境下种群分布格局及空间关联性的响应研究较少。本研究应运点格局分析法对不同坡向红砂和西北针茅的空间分布格局及其关联性进行研究,旨在了解红砂和西北针茅种群在不同坡向生境中的空间分布特征,揭示皋兰县干旱荒漠区植物演替过程中,优势种群之间的竞争关系和生存、繁衍策略,更好地了解群落维持和生态系统演替趋势,对红砂和西北针茅种群在干旱荒漠区水土保持及生态环境变化的应对策略提供科学参考依据。
1 研究地区与研究方法
1.1 研究区概况
研究区地处甘肃中部的兰州市皋兰县水阜乡老虎台,坐标为103°49′E,36°16′N,海拔约在1 840 m左右,位于黄河上游,属黄土高原丘陵沟壑区。气候主要为温带半干旱气候,受大陆性气候的影响,该地区降水量少,年均降水量266 mm,蒸发量为1 660 mm,年日照时数约2 768 h,气候干燥,昼夜温差大,年均气温7.2 ℃,无霜期144 d。土质属于黄土高原土壤侵蚀类型,地下水匮乏,干旱严重,蓄水保墒性差,生态环境脆弱,植被稀疏,生态系统退化严重。植被主要为多年生草本,双子叶植物,主要植物种群有西北针茅、红砂、狗尾草(Setariaviridis)、荒漠锦鸡儿(Caraganaroborovskyi)和蓍状亚菊(Ajaniaachilloides)等。
1.2 样地设置与调查
2017年8月中旬在甘肃省皋兰县老虎台进行了野外的群落学调查。在同海拔高度上,选取西北坡、西坡、西南坡及南坡坡向具有代表性的4个样地,并在每个坡向上分别设置10 m×10 m的样方,再将每个样方划分为100个1 m×1 m的小样方。在各个坡向上,记录每个小样方内的植物种类、数量、盖度等,并以样方左下角顶点为基点,在对应的坐标纸上记录样方中红砂和西北针茅个体的地面投影点,确定红纱和西北针茅的相对位置[15]。
1.3 研究方法
1.3.1点格局分析本研究采用RipleyK函数分析种群的空间分布格局。该函数以植物个体的空间坐标(位置) 为基础,分析不同尺度下种群的空间分布格局,反映种群生态特征,其公式如下[16]:
(1)
式中:A为研究区(样方)的面积;n为样地内物种的数量;uij表示第i株与第j株个体间的距离;r表示空间尺度,当uij≤r时,Ir(uij)=1,当uij>r时,Ir(uij)= 0;Wij为权重值,用于边缘校正。在实际应用中,通常用改进后的Ripley’sL函数代替:
(2)
当L(r) = 0,种群表现为随机分布;L(r)>0,种群表现为聚集分布;L(r)<0时,种群表现为均匀分布。
为了提高分析种群空间分布格局的精度,通过Monte-Carlo 随机模拟,得到两条包迹线围成的置信区间,计算出不同尺度下的L(r)的值,在包迹线以上,为聚集分布;在包迹线以内,为随机分布;在包迹线以下,为均匀分布。
1.3.2空间关联分析种间的空间关联性是指两个种的点格局分析,也称为多元点格局分析[17]。将RipleyK函数应用于双变量形式进行计算,具体如下:
(3)
式中n1、n2分别代表种群1及种群2的个体数,其他指标与(1)式含义相同,同样,用L12(r)取代K12(r),公式为:
(4)
当L12(r)>0时,表示两种群在r尺度上正相关;当L12(r)=0时,表示两种群无相关;当L12(r)<0时,表示两种群负相关。采用Monte-Carlo 随机模拟方法拟合包迹线,计算出不同尺度下的L12(r)的值,以确定不同物种间是否具有空间关联性,L12(r)的值在包迹线以上表明为正相关,在包迹线之间为无相关,在包迹线以下呈负相关。
1.3.3数据处理本研究选取的空间尺度为5 m。通过Monte Carlo法随机模拟99次,得到两条包迹线围成的99%置信区间。调查数据采用Get Data Graph Digitizer 2.25软件进行坐标提取,通过Excel 2010、Programita 2014等软件进行分析处理。利用Sigmaplot12.0进行绘图。
2 结果与分析
2.1 不同坡向红砂与西北针茅种群的生物学特征
红砂及西北针茅种群在4个坡向的分布具有显著差异,红砂种群在西南坡分布最多,西南坡属半阳坡,阳光充足,土壤较为干旱,其生态环境适宜红砂的生长,重要值达0.828,具有一定的优势度,西北坡分布相对较少(表1)。随着坡向梯度的变化,其盖度、株数及重要值呈现出自西北坡向西南坡逐渐增大,再到南坡缓慢减小的趋势。而西北针茅在西北坡分布最多,西北坡属半阴坡,阳光适宜,土壤较为湿润,适宜西北针茅的生长,所对应的重要值达0.861,南坡分布则相对较少,随着坡向由西北坡向西南坡的转变,其盖度、株高及重要值呈现依次递减的趋势。
2.2 红砂和西北针茅种群的点格局分析
红砂与西北针茅两种群个体分布点图(图1)显示,红砂种群的分布密度:西南坡>南坡>西坡>西北坡,西北针茅种群的分布密度:西北坡>西坡>西南坡>南坡。在不同的空间尺度上,种群的分布点图表现出较大的差异性,说明两个种群的分布格局与空间尺度有密切关系。
红砂种群在不同坡向上的分布状况,在西北坡向上,红砂种群在0~2.1 m小尺度范围内表现为聚集分布,2.1~5 m尺度上为随机分布;西坡坡向上,红砂种群在0~5 m研究尺度内均呈现出随机分布;西南坡向上,红砂种群在0~3.7 m尺度上为聚集分布,3.7~5 m尺度上为随机分布;南坡坡向上,红砂种群在整个研究尺度上呈现出聚集分布(图2)。坡向由西北坡到南坡转变的过程中,西北针茅在研究尺度内的分布格局,在西北坡与西坡坡向上,西北针茅种群在整个尺度内均表现出聚集分布;西南坡坡向上,西北针茅种群在0~1 m及4.2~5 m尺度上表现为随机分布,1~4.2 m尺度上表现为聚集分布;南坡坡向上,西北针茅种群在0~5 m尺度范围均呈现为随机分布(图3)。结果说明不同坡向具有不同的生境环境,种群在不同的生境环境中所表现的生长特征具有显著的差异性。
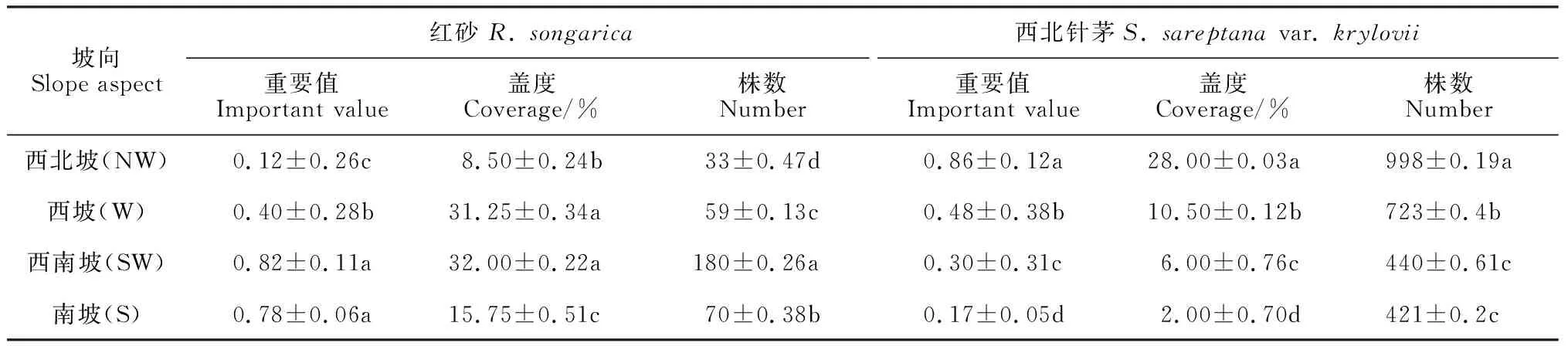
表1 红砂和西北针茅种群在不同坡向的变化特征
注:同列不同的小写字母表示红砂和西北针茅其重要值、盖度、株数在不同坡向上的分布有显著性差异,P<0.05
Note: Different lowercase letters in the same column indicate that there are significant differences in the distribution of important value, coverage and number of plants betweenR.songaricaandS.sareptanavar.kryloviiin different slopes,P<0.05

图1 不同坡向的红砂和西北针茅的空间分布点图Fig.1 Spatial distributions of R. songarica and S. sareptana var. krylovii on different aspects

图2 红砂在不同坡向的空间分布格局Fig.2 Spatial patterns of R. songarica on different aspects

图3 西北针茅在不同坡向的空间分布格局Fig.3 Spatial patterns of S. sareptana var. krylovii on different aspects

图4 红砂与西北针茅在不同坡向的关联性Fig.4 Spatial association between R. songarica and S. sareptana var. krylovii on different aspects
2.3 红砂与西北针茅种群的空间关联性
红砂与西北针茅种群的种间关联性为,西北坡向上,0~2.1 m小尺度研究范围内表现为负关联,较大尺度上表现为无关联;西坡坡向上,两种群在0~1.4 m小尺度内表现为负相关,在1.4~5 m尺度范围上表现为无关联;西南坡和南坡坡向上红砂种群与西北针茅种群在整个研究尺度范围均表现为无关联(图4)。说明在不同坡向上,红砂与西北针茅种群经过长期的种内、种间关系及环境因子的相互作用,表现出种群空间关联性具有显著差异性。
3 讨 论
坡向是山地的主要地形因子之一。在海拔相同的情况下南坡受到较多的太阳辐射,致使坡面有较高的温度与较强的蒸发,分布着具有耐旱结构的植物群落。郑成洋等[18]通过研究福建黄岗山东南坡和西北坡乔木物种多样性及群落特征的垂直变化认为西北坡接受的热量相对西南坡较少,生长着喜阴的植物群落。本研究发现,红砂与西北针茅种群在不同坡向梯度上的空间分布格局具有显著差异性,西北坡西北针茅分布较多,而西南坡红砂分布较多。随着坡向由西北坡、西坡、西南坡到南坡的变化中,西北针茅种群的盖度及重要值呈递减趋势;而红砂种群呈现出先逐渐增加,到南坡缓慢降低的趋势。这主要与红砂和西北针茅的生物学特性有关,红砂种群适宜在干旱,半干旱地区生长,因此在南坡西南坡生长的较好,西北针茅个体喜在较湿、蒸发量较少的地区生长,因此在西北坡分布最多,这体现了植物种群对环境异质性的响应策略。
种群空间点格局特征的形成与变化主要是种群自身的生物学特性与环境因子共同作用的结果,不同的空间格局表现出种群对环境变化的应对机制,利于种群更有效地利用资源[19]。空间分布格局中聚集分布体现了种群内部之间相互有利(正向)的生态关系,均匀分布体现了相互不利(负向)的生态关系,随机分布则意味着种内个体间相互独立,无明显生态关系[20]。本研究发现,西北坡向上红砂个体的分布较少,在0~2.1 m小尺度范围内表现为聚集分布,2.1~5 m尺度上表现为随机分布,其主要原因是由于种群个体幼小,缺乏对空间资源的有利竞争,幼小个体之间需要相互庇护,以此来抵抗外界的干扰。赵峰侠等[21]研究表明,种群在小尺度内的聚集分布利于其形成群体效应,利于自身适应环境并能够增强种间竞争力,促进种群的正常生长,随着尺度的增加,红砂种群聚集程度减弱并趋于随机分布,是由于西北坡向上土壤较为湿润,蒸发量较少,不适宜红砂的生长,使得红砂分布密度过小,因此在较大尺度上红砂表现为随机分布;西坡坡向上,红砂在任何研究尺度内都表现为随机分布,主要是西坡处于西北坡与南坡的过渡段,红砂种群分布密度相对增加,且物种多样性丰富,种间排斥作用加剧,因此红砂表现为明显的随机分布;西南坡向上红砂种群在0~3.7 m的尺度范围上表现为聚集分布,3.7~5 m尺度上呈现出随机分布,是由于西南坡阳光较充足,满足红砂的生长需求,因此,红砂种群密度较大,在小尺度内表现为聚集分布,随着研究尺度的增加,红砂种群聚集程度减弱,呈现出随机分布,刘旻霞等[22]通过研究甘南亚高寒草甸坡向梯度上矮嵩草与珠芽蓼种群点格局及其关联性认为种群除了受其他物种排斥外,也会因种内竞争发生自疏现象,使得种间排斥作用加剧,因此在较大尺度上趋于随机分布;南坡坡向上,红砂种群在整个研究尺度上都表现为聚集分布,可能是因为南坡较为干旱,光照较强,其环境有利于种群的生长、扩散,因此导致红砂的聚集强度增加。西北针茅种群在各个坡向上的空间分布格局具有差异性,杨君珑等[19]研究认为小规模的聚集由植物本身的特征所决定,而大规模聚集是植物从原分布中心扩散,由环境因子影响决定,在西北坡与西坡坡向上,西北针茅种群在整个研究尺度范围内均表现为聚集分布,主要是与种子在母株周围就近传播及自身的生物学特征有关,西北坡和西坡土壤较湿润,温度相对较低,适宜西北针茅的生长,使得西北针茅呈现为聚集分布;西南坡向上,西北针茅种群在0~1 m小尺度及4.2~5 m较大尺度上呈现出随机分布,主要是因为随着坡向梯度的变化,西南坡向上水分蒸发量较大,土壤较干旱,所以在0~1小尺度上,西北针茅种内互相竞争,争夺水分及土壤养分,使得空间分布呈现为随机分布,在4.2~5 m较大尺度上表现为随机分布,这是对空间尺度的一种响应,张璞进等[23]研究发现物种的空间格局具有尺度效应,物种间在更大的范围内缺少竞争,物种趋于随机分布,在1~4.2 m尺度上呈现出聚集分布,可能是西北针茅适应干旱环境的一种策略,是由于受到物种的生物特性及地形、土壤、水分等综合因素的影响,西北针茅分布密度仍然较大,因此,西北针茅在西南坡向上仍然以聚集分布为主;南坡坡向上西北针茅在任何尺度内呈现出随机分布,主要是由于南坡水分短缺,种内及种间竞争激烈致使老年和幼小个体死亡,种群表现出随机分布。
种间关联性是指不同物种在空间分布上的相互联结性,是群落形成、维持和演化的基础[24]。种间关联理论认为,正负关联特征可能是物种间相互作用的结果,也可能是物种生境趋同或趋异的表现[25]。本研究中,两种群在小尺度范围上表现为负关联,随着尺度的增大,空间分布格局趋于不关联,这与前人的结论一致[26-27],红砂与西北针茅种群在西北坡,西坡坡向上小尺度范围内表现为负关联,是因为在一定资源限制下,种群之间对土壤养分及水分竞争激烈,种间存在排斥,使得种群间呈现出负关联,随着研究尺度的增大,两种群空间格局表现出无关联,是由于红砂在西北坡和西坡坡向上的分布密度较小,对西北针茅的生长不会产生实质性的影响;在西南坡与南坡坡向上,红砂和西北针茅种群在整个研究尺度内均表现为无关联,可能是因为两种群均为西南坡与南坡的优势物种,二者之间表现出正向的相互关系,竞争成为次要的种间关系,资源策略由争夺转变为共享,种间关联性表现为不相关[28]。
综上所述,甘肃省兰州市皋兰县老虎台干旱荒漠区红砂和西北针茅种群的空间格局及关联性对坡向梯度具有明显的响应机制,这种变化不仅与其自身生物学特性有关,而且还受种群间相互关系及环境异质性的影响。本研究有助于我们认识干旱荒漠区植物种群的种内、种间复杂的竞争过程、不同微生境植物的生存策略及群落的演替趋势,更好地了解种群维持及动态变化机制,对皋兰县植被的恢复、水土保持、物种的更新与可持续经营具有重要意义。
