干热胁迫对杨树叶片气体交换和叶绿素荧光参数的影响
2019-01-04邹吉祥白鑫磊高榕择李新林程文琼金纯伊
邹吉祥,白鑫磊,高榕择,李新林,朱 桐,程文琼,金纯伊,金 华
(大连民族大学 环境与资源学院,辽宁 大连 116600)
随着全球气候变暖和水资源的短缺,我国常出现持续高温和严重的干旱现象,部分地区极端气温甚至超过40℃[1]。干旱和高温会通过多种方式来间接或直接影响植物的生理生化过程[2-3],可降低植物光合反应速率以及通过PSII的电子传递过程影响光合反应速率[4]。干旱和高温胁迫往往协同发生[5],干旱和高温复合胁迫对植物的危害比单一胁迫更大。干热胁迫对植物生理特性影响的研究主要集中在苜蓿、草坪草等草本植物[6],在木本植物方面的研究报道较少。目前对杨树生理生化过程方面的研究报道多集中在干旱胁迫[7]、高温胁迫[8]、低温胁迫[9]、盐胁迫[10]等单一胁迫,而干热胁迫影响杨树叶片气体交换和叶绿素荧光参数的复杂机理有待深入研究,因此研究杨树在干热胁迫下的生理响应机制具有重要意义。
1983年年初,课题组在辽宁西部新民县大柳屯乡吴屯村发现了一种被当地老百姓称之为吴屯杨Populus wutunensis的杨树无性系,吴屯杨适应性强,生长速度快,具有耐盐碱、耐干旱能力强的特点[11],在三北地区均有种植。本研究选用吴屯杨和辽西北主栽的小青杨Populus pseudo-simonii Kitag.、小 美 旱 杨 Populus simoniih(Populus pyramidalis+ Salihmatsudana)为试验材料,研究不同干热胁迫处理对杨树叶片气体交换和叶绿素荧光参数以及光合色素的影响,以探讨杨树在干热胁迫下的生理生化机制,为杨树在干热地区推广种植提供理论依据。
1 材料与方法
1.1 材料与处理
供试材料为吴屯杨、小青杨和小美旱杨,于2016年5月在辽宁省阜新市苗圃采集一年生枝条。选择无病虫害、粗细一致的枝条,在茎干中下部截取20 cm作为插条,洗净后移入1/2浓度的Hoagland's(霍格兰氏)营养液中,在25 ℃室温下进行水培,每3 d换1次营养液。30 d后选择根系发达且苗木生长一致的植株放入光照培养箱中模拟干热胁迫处理。将上述所选植株平均分配至4个光照培养箱中,更换为含有1% PEG-6000的1/2霍格兰营养液模拟干旱条件,温度分别设置为25 ℃/25 ℃、30 ℃ /25 ℃、35 ℃ /25 ℃、40 ℃ /25 ℃,以不添加1% PEG-6000的1/2霍格兰营养液(25℃/25℃)作为对照,设处理编号为:编号1,无1%PEG、25 ℃/25 ℃,即空白对照组;编号2,1%PEG、25 ℃ /25 ℃;编号3,1%PEG、30 ℃/25 ℃;编号4,1%PEG、35 ℃/25 ℃;编号5,1%PEG、40 ℃/25 ℃。所有光照培养箱设置为昼/夜12 h/12 h,光照强度设定为 1 000 μmol·m-2s-1,相对湿度为70%。处理24 h后测定中上部叶片各项生理指标,试验设3次重复。
1.2 气体交换参数的测定
采用Li-6400 便携式光合测定仪(Li COR,USA)和配套使用的红蓝光源叶室测定杨树叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等值。测量时设定光强为 1 000 μmol·m-2s-1,温度为 25 ℃,CO2浓度为400 μmol·m-2s-1,其他测定条件与周围环境保持一致,测定时每个叶片测定5次数据,取平均值。
1.3 叶绿素荧光参数的测定
采用Li-6400 便携式光合测定仪(Li COR,USA)和配套使用的荧光测量叶室测定各处理组杨树叶片的叶绿素荧光参数,待测杨树叶片经暗适应30 min后,测定荧光参数Fo、Fm和Fv/Fm,置于光照的条件下测定荧光参数Fs、Fo′、Fm′和Fv′/Fm′。根据相应的公式计算出其他的荧光参数。计算方法如下。
光化学猝灭系数:qP = (Fm′-Fs)/(Fm′-Fo′) 。
非光化学猝灭系数:qN=1-(Fm′-Fs)/(Fm′-Fo′)。
实际光化学效率:ΦPSⅡ= (Fm′-Fs)/Fm′。
电子传递速率:ETR = ΦPSⅡh0.5h0.84 。
1.4 光合色素的测定
选取处理后的杨树叶片,洗净后剪成0.5 cm宽的叶段,称取0.2 g新鲜叶片,参照李合生(2000)的方法测量杨树叶片中光合色素含量, 使用UV-2100型紫外分光光度计测定并计算叶绿素a、叶绿素b、总叶绿素及类胡萝卜素含量。
1.5 统计分析
实验数据利用SPSS 22.0软件统计分析,采用邓肯氏新复极差法进行数据差异显著性检测。
2 结果与分析
2.1 干热胁迫对不同种杨树叶片气体交换参数的影响
从图1可以看出,3个杨树品种的净光合速率(Pn)和气孔导度(Gs)随着干热胁迫的程度增加均呈先上升后下降的趋势,在处理编号2时达到最大值。当净光合速率和气孔导度达到最大值后,3种杨树的净光合速率随着胁迫强度的加大而下降,即使在重度胁迫处理下,小青杨和吴屯杨叶片的净光合速率仍然比对照组高,小美旱杨略低于对照组。3种杨树在轻度干热胁迫时蒸腾速率(Tr)呈缓慢上升的趋势,3种杨树的蒸腾速率峰值对不同程度干热胁迫的反应有较大区别,干热胁迫使吴屯杨和小美旱杨叶片的蒸腾速率最大值比对照分别增加了3.6倍和4.5倍,而小青杨叶片的蒸腾速率始终随着胁迫处理的增强而增加,在重度干热胁迫时达到最大值,比对照增加5.8倍。胞间CO2浓度(Ci)整体呈缓慢下降趋势,小青杨在重度胁迫低于吴屯杨和小美旱杨。
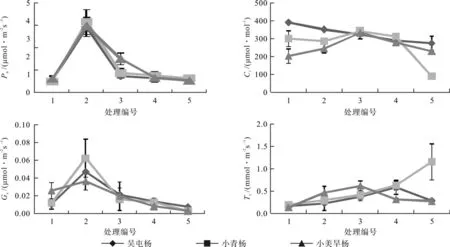
图1 干热胁迫对杨树叶片气体交换参数的影响Fig.1 Effects of drought and heat stress on leaf gas exchange parameters
2.2 干热胁迫对不同种杨树叶片荧光参数的影响
从图2可以看出,随着干热胁迫程度的增加,3种杨树的PSII最大光化学量子产量(Fv/Fm)和PSⅡ有效光化学量子产量(Fv′/Fm′)均呈下降趋势,处理4的PSII最大光化学量子产量呈快速下降趋势,处理编号5达到最小值,吴屯杨、小青杨和小美旱杨比对照分别降低了98%、99%和97%。小青杨的光化学淬灭系数(qP)随着干热胁迫的增加而升高,处理编号5达到最大值,而吴屯杨和小美旱杨的光化学淬灭系数处理编号2显著提高,随后干热胁迫导致光化学淬灭系数的下降,表明重度胁迫降低了PSⅡ的传递活性。非光化学淬灭系数(qN)随着胁迫程度的增加变化较平稳,与光化学淬灭系数的变化趋势相反。小青杨的实际光化学效率(ΦPSⅡ)和光合电子传递速率(ETR)均随着干热胁迫的增加而升高,吴屯杨和小美旱杨的实际光化学效率和光合电子传递速率则先升高后下降,在重度干热胁迫下处理编号5小青杨的实际光化学效率是对照的1.39倍,吴屯杨与对照基本一致,而小美旱杨的实际光化学效率相比对照下降了61.33%,光合电子传递速率与其变化趋势较相似。
2.3 干热胁迫对不同种杨树叶片光合色素含量的影响
随着干热胁迫强度的增加,3个杨树品种叶绿素a的含量呈下降趋势,在重度干热胁迫下处理编号5均达到最小值,其中小美旱杨的叶绿素a含量下降幅度最大。同对照相比,重度干热胁迫的处理编号5使小青杨、吴屯杨和小美旱杨的叶绿素a含量分别降低了8.32%、10.66%和44.89%。3个杨树品种叶绿素b的值也随着干热胁迫强度的增加显著下降,且在重度干热胁迫的处理编号5时降到最小值,与对照相比小青杨、吴屯杨和小美旱杨的叶绿素b的含量分别降低了23.03%、36.19%和50.32%,小美旱杨的叶绿素b含量下降显著。随着干热胁迫强度的不断增加,小青杨、吴屯杨和小美旱杨3个杨树品种总叶绿素含量呈降低趋势,在重度干热胁迫下处理编号5降至最低,这与叶绿素a和叶绿素b含量变化趋势一致。不同干热胁迫强度对3个杨树品种的类胡罗卜素含量有较大影响,都随着干热胁迫程度的增强而上升,小青杨、吴屯杨和小美旱杨的类胡罗卜素含量均在重度干热胁迫的处理编号5达到最大值,其中小青杨和小美旱杨上升明显,相比对照上升了34.61%和34.95%,吴屯杨相比对照则上升了13.33%。

图2 干热胁迫对杨树叶绿素荧光参数的影响Fig.2 Effects of drought and heat stress on chlorophyll fluorescence parameters
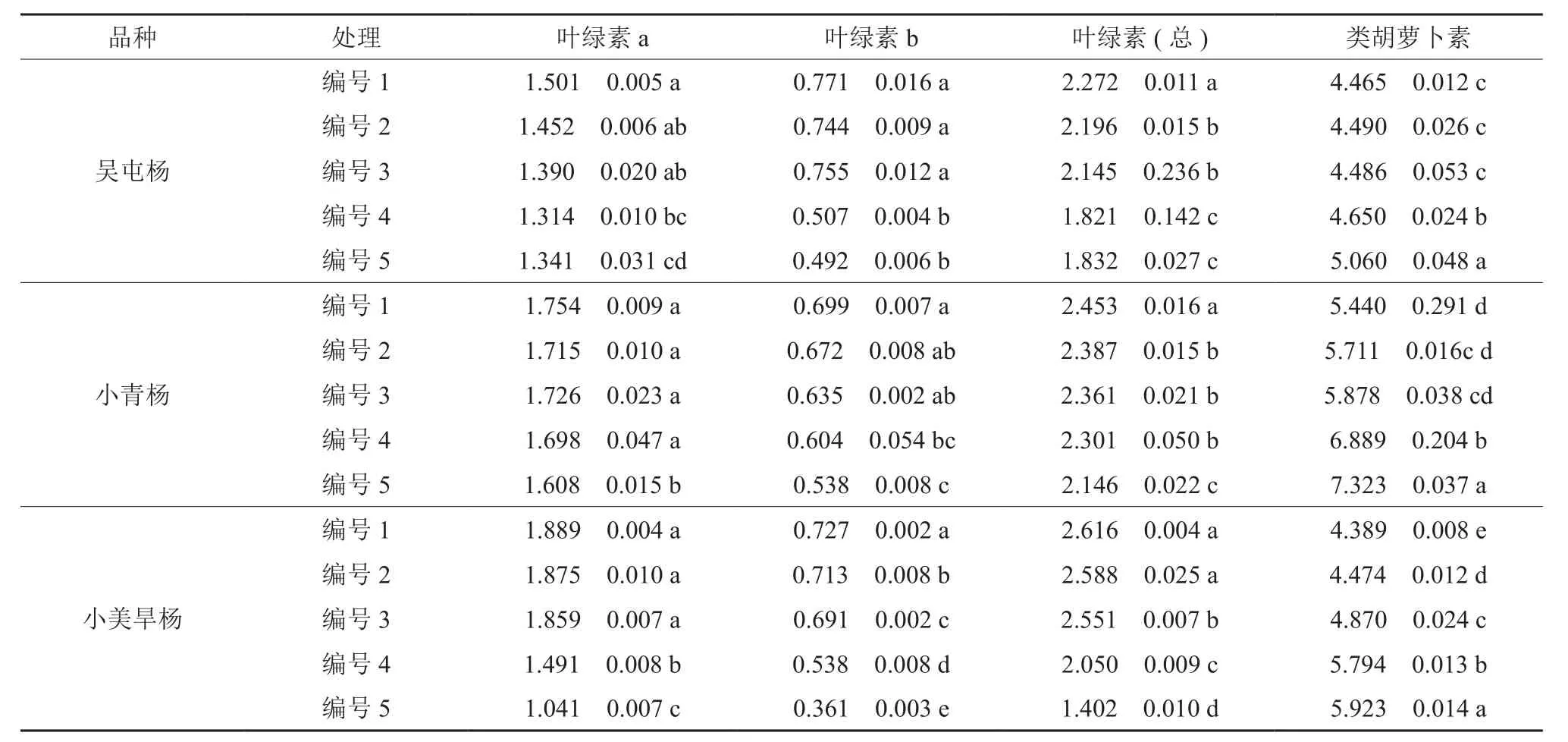
表1 干热胁迫对杨树叶片光合色素含量的影响Table 1 Photosynthetic pigment contents in the leaves of three Populus cultivars under drought and heat stress (mg·g-1)
3 讨论与结论
植物的光合作用是一个极其复杂的CO2气体扩散过程和发生在光合反应位点的生理生化反应过程[12],同时也是植物对环境温度变化响应最为敏感的生理过程之一[13-14]。本研究结果显示,3个杨树品种净光合反应速率的最大值均出现在处理编号2,随着干热强度的加大而明显降低,但即使是在重度胁迫环境下,小青杨和吴屯杨叶片的净光合速率仍然比对照组高,小美旱杨略低于对照组,说明一定程度的干热胁迫提高了杨树叶片的净光合反应速率,但随着胁迫的加剧,破坏了光合器官的结构和功能,导致净光合反应速率急剧下降。干热胁迫使吴屯杨和小美旱杨叶片的蒸腾速率 先升高后降低,其中小青杨叶片的蒸腾速率始终随着胁迫程度的增强而增加,并且在重度干热胁迫时才达到其最大值。说明吴屯杨、小美旱杨和小青杨有着适应干旱高温生境的光合特征,当胁迫加剧,吴屯杨和小美旱杨的叶片出现衰老,生理作用减缓,所以蒸腾速率降低。胞间CO2浓度和气孔导度均在处理编号3之后迅速下降,说明在中度干热胁迫时3种杨树的叶片光合作用下降主要是气孔限制导致的[15],在重度干热胁迫下迅速下降是由非气孔限制因素导致的,可能是光合反应位点、电子传递链和 PSⅡ开放等产生损伤以及RuBP梭化酶活性下降导致了杨树光合作用下降。光合作用反应中心PSⅡ在逆境胁迫条件下会出现可逆性的失活或不可逆的伤害,导致光合原初反应过程受阻,从而影响植物光合作用[16]。吴屯杨和小美旱杨的qP、ΦPSⅡ、ETR变化趋势都是先升高后降低,小青杨则均随胁迫强度的增加而升高并均在处理编号5时达到最大值,这表明重度干热胁迫条件下使PSII最大光化学量子产量和PSⅡ有效光化学量子产量降低,PSⅡ开放的反应中心数目减少,QA→QB的电子传递受到抑制,从而影响光合反应过程。
叶片光合色素含量是反映植物光合能力的一个重要指标 , 环境因子的改变会引起叶绿体色素含量的变化,进而引起光合性能的改变[17]。有研究表明,叶绿素含量与光合速率关系密切,而叶绿素在叶内处于不断合成与分解的动态平衡,其含量受多种因素影响。多种逆境胁迫伤害植物的机理之一就是破坏植物体内自由基攻击膜系统,造成膜脂过氧化从而引发植物生理代谢失调进而植物受到伤害[18]。类胡萝卜素分子能够接受过剩的激发态叶绿素分子的能量,从而避免形成单线态氧,起到一种光保护作用。本研究显示,随着干热胁迫的增强,3个杨树品种的类胡萝卜素均呈现上升的趋势,这表明可能是干热胁迫促进了类胡萝卜素的合成,是一种抵抗逆境的表现能力。叶绿素a、叶绿素b、总叶绿素的含量随着胁迫程度的增强都呈现下降的趋势,可能是干热胁迫导致叶绿素的合成速率降低,干热胁迫可以使体内活性氧的积累增加,加速了叶绿素的降解和类胡萝卜素的增加,也是造成叶绿素含量降低的原因之一。另外我们还发现,在重度干热胁迫下处理编号5小青杨和吴屯杨的叶绿素a、叶绿素b、总叶绿素和类胡萝卜素的含量变化幅度比小美旱杨小,小美旱杨的叶绿素a、叶绿素b和总叶绿素均明显低于对照。由此可以推断 3 个品种的耐干热能力为小青杨和吴屯杨较强,小美旱杨最弱。
