不同林龄银合欢生长季土壤呼吸影响因素分析
2018-12-13严思维吴建召罗清虎林勇明王道杰
崔 羽,严思维,吴建召,罗清虎,林勇明*,王道杰
(1.福建农林大学林学院,福建 福州350002;2.省教育厅福建省高校森林生态系统过程与经营重点实验室,福建 福州350002;3.中国科学院水利部 成都山地灾害与环境研究所,四川 成都 640001)
土壤呼吸(Soil respiration)指土壤在其新陈代谢中产生大量CO2释放到大气中的过程[1],主要包括3个生物化学过程:植物根系呼吸、土壤微生物异氧呼吸和土壤动物呼吸;1个化学氧化过程:少量土壤有机质氧化产生二氧化碳[2]。在整个陆地生态系统中,森林作为主体,其土壤呼吸占总呼吸量的69%,森林土壤呼吸的变化影响着全球碳平衡[3],因此,为进一步认识森林土壤碳库对全球碳循环的影响,需对森林土壤的CO2释放量与呼吸动态变化进行研究。近年来,森林土壤呼吸成为陆地生态系统碳循环过程和全球气候变化研究的重要和热点内容之一,国内外学者针对森林土壤呼吸开展了大量的研究工作,然而当前对于森林土壤呼吸的研究主要侧重于由植被类型、分布范围等产生的差异性及主要影响因素[4-6],研究发现土壤呼吸受土壤温湿度、土壤有机质和氮含量、pH、凋落物、植被类型、根系生物量等生物因子、非生物因子与人类活动共同作用影响[7],而人工林生态系统中不同林龄土壤呼吸特征与影响因素的研究却相对较少。人工植被营造措施是干热河谷脆弱生态系统生态恢复与水土流失治理的主要方法之一,其对区域性碳循环与平衡的影响机制、碳库的贡献特征等问题尚不明确,因此,开展脆弱生态系统人工林土壤呼吸及其与理化性质关系的研究,对于探讨脆弱生态系统人工造林对碳循环和平衡的影响效应、人工林土壤碳库循环和平衡具有重要的科学意义。
云南东川蒋家沟流域自1988年以来,人工引种银合欢进行造林,银合欢喜光、耐高温干旱,少有病虫害;深根性,能固氮,生长迅速,萌生力和自然更新能力强,对土壤要求不严,对该地区多变的温度和湿度变化有较好的适应能力,是目前金沙江干热河谷地区优良的造林树种[8],现保存有较完整的不同年龄银合欢人工林群落。前人对该区域银合欢人工林的研究主要集中在群落根系固土垂直效应、枝系分布特征以及各级分枝率、群落结构与物种组成、土壤理化性质变化等方面[9-12],但在多变的水热条件下,银合欢生长季内的土壤呼吸与生态因子之间的动态关系的研究尚未见到报道,从而无法准确把握干热河谷脆弱生态系统在生长季的碳循环特征,不利于营林管理措施的总结优化。因此,本研究采用Li-8100土壤呼吸仪(配带土壤呼吸室),对云南蒋家沟流域不同林龄银合欢生长旺季(8月)土壤呼吸动态变化及其影响因子进行观测与分析,旨在揭示不同林龄银合欢林地土壤呼吸与土壤环境之间的关系,从而解释泥石流频发区人工植被定植、存活、生长的策略,并为进一步揭示干热河谷脆弱生态系统人工林土壤CO2释放的过程以及传递机理提供基础数据,也可为科学估算我国西南地区人工林土壤碳库动态和生态系统碳平衡研究提供参考。
1 研究区基本概况和方法
1.1 基本概况
研究区所在的蒋家沟位于金沙江下游一级支流小江流域内,地理坐标为 103°06′~103°13′E、26°13′~26°17′N。该区新老构造错综复杂,新构造运动尤其活跃,岩性一般较为软弱(以易风化破碎的砂岩和板岩为主),岩层走向十分破碎。地貌类型为侵蚀中山,地势自东到西呈阶梯状下降,海拔范围1042~3269 m,流域内低于1600 m的区域为干热河谷带[13]。干湿季分明,5~10月为湿季,降水量占全年88%。该区多年平均气温大于20℃,极端高温可达40.9℃,最低气温-6.2℃;多年平均降水量约691 mm,平均蒸发量约3638 mm,蒸发量为降水量的5倍以上[14]。长期的水土流失和泥石流作用导致该区域石漠化严重,土壤为含砾石较高的初育土。
1.2 样地设置
试验时间为2015年7月25日至8月26日,选择中国科学院东川泥石流观测研究站西侧坡面的10年生、16年生和27年生银合欢人工林为调查样地。由于取样点坡度较大,林木分布较分散,10年生银合欢林分布面积约为600 m2,仅可设置一个30 m×20 m样地,为实现对照取样,本研究在3个年龄的林地中均选取了30 m×20 m样地;其中10年生与另两个样地的间隔约50 m,16年生和27年生样地间隔约20 m[12],并对其进行样地调查(表1)。在样地调查后,在每个林龄样地,设置3个10 m×10 m样方,用于阻根处理、去凋落物处理和保留凋落物处理,每个处理重复3次。利用挖壕法进行阻根处理,即在样地四周挖垂直深度约50 cm(不见根系的深度)的壕沟,随后将植物根系切断,插入塑料膜作为障碍物,阻止根向样地内生长[15];去凋落物处理即将环内枯落物清走,在土壤环上放置孔径1 mm×1 mm的细网,防止凋落物进入[16];保留凋落物处理即正常处理。处理后分别嵌入PVC土壤环(Φ20 cm×H10 cm),夯入土中 5 cm,并夯实基座外的土壤,试验期间不再移动PVC土壤环。
林下植被样地概况:10年生银合欢林下主要植被以银合欢幼苗为主,并伴有少量的茅草(Imperata cylindrica(L.)Beauv.)和 小 叶 荩 草 (Arthraxon lancifolius(Trin.)Hochst.),林下群落总盖度约为 59%;16年生银合欢主要以银合欢幼苗为主,林下植被主要有多花杭子梢(Campylotropis macrocarpaBge.Rehd.)、小叶荩草、剑麻(Agave sisalanaPerr.ex Engelm.)等,林下植被总盖度约为51%;27年生生银合欢林下主要以银合欢幼苗为主,有少量灌木,分别为苦刺(Solanum deflexicarpumC.Y.Wu et S.C.Huang)和剑麻,草本多为黄背草 (Themeda triandra Forsk.Var.Japonica(Willd.)Makino)和小叶荩草,林下植被总盖度约为55%。
1.3 样品与数据采集
本研究采用LICOR公司研发的LI-8100开路式土壤碳通量测定系统测定土壤呼吸的各项指标。待各处理样方稳定后,每间隔5天测定1次日平均值,测定时间为早上9:00-11:00,共测5次。若遇降雨,在雨停天晴2日后测定,若持续阴天,则在无降雨的时候测定;在测定土壤呼吸的同时,用军工铲挖取不同林龄0~20 cm的土壤样品,按五点取样法均匀混合,装入标记好的自封袋内带回实验室分析,一部分放置于4℃冰箱中保存留用以测定酶活性。于8月17日、23日、24日3天对3个样地分别进行日动态测定。日动态测定前1天对样方进行检查和测定前准备,在不干扰土壤圈的前提下,从根部齐地剪出地上植物。日动态测定时间为7:00~17:00,时间间隔为2 h,每个处理每次仪器重复3次。在测量土壤呼吸的同时,分别记录LI-8100自带温度计和水分传感器温度和水分变化。
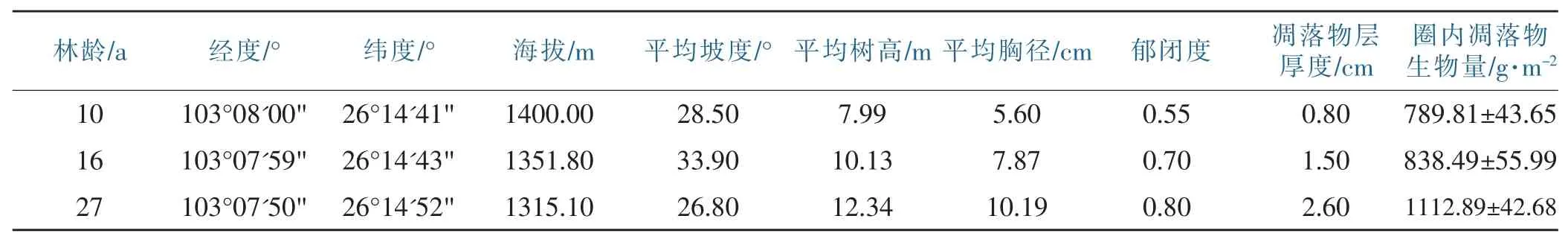
表1 不同林龄银合欢样地概况Table 1 Characteristics of Leucaena leucocephala forests in different ages
1.4 样品测定
蔗糖酶(Invertase)和纤维素酶(Cellulase)活性:采用3,5-二硝基水杨酸比色法测定;过氧化氢酶(Catalase)活性:采用紫外分光光度法测定;土壤有机质(OM):采用重铬酸钾-外加热法测定;土壤全氮(TN):采用凯氏定氮法测定;土壤水解氮(AN):采用碱解-扩散法测定;土壤 pH :pH 计(STARTER3100)测定[15,17-18]。
1.5 数据分析
土壤呼吸速率和温度根据指数模型y=aebx,y表示土壤呼吸速率,x表示土壤温度,a为0°C时土壤呼吸速率,b为温度反映系数。Q10=e10b表示土壤呼吸对温度的响应程度。
运用SPSS19.0对数据进行单因素 (one-way ANOVA)方差分析、双变量相关分析中的Pearson相关分析和LSD检验,利用Excel 2003绘制图形。
2 结果与分析
2.1 不同林龄银合欢土壤呼吸日动态
不同林龄银合欢林土壤呼吸速率的日动态变化规律基本一致,表现为先增大后减小的趋势。一般从7:00开始缓慢上升,随着土壤温度的上升,土壤呼吸达到最大,在13:00–15:00达到峰值,随后土壤呼吸逐渐减小(图1)。10、16、27年生银合欢林土壤呼吸速率日动态变化分别为 2.01~2.30、1.17~1.93、1.21~1.67 μmol·m-2·s-1,日平均土壤呼吸速率为 2.10、1.65、1.46 μmol·m-2·s-1;其呼吸速率高低顺序依次为:10 年生>16年生>27年生。从整个观测周期可知:10年生银合欢林地土壤呼吸速率始终高于16、27年生,且在13:00达到峰值;而16年生土壤呼吸速率随着时间的推移和温度的上升而超过27年生林地呼吸,中老林龄土壤呼吸峰值较幼林龄晚,两者在15:00达到峰值。
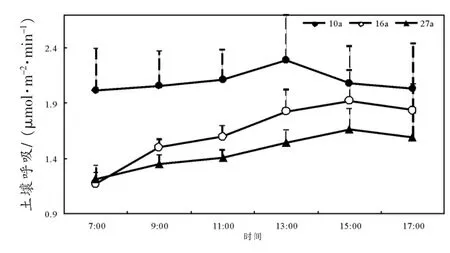
图1 不同林龄银合欢土壤呼吸速率日动态变化Fig.1 Diurnal variations of soil respiration rate in different ages of Leucaena leucocephala
不同林龄银合欢林地生长季土壤呼吸速率、温度、湿度变化曲线随着日期的推移呈现出规则的单峰变化曲线(图 2 a、b、c),即在 5~15 日之间呈现上升趋势,20日均呈现显著下降趋势,25日又有回升的趋势。3种林龄的土壤呼吸速率变化基本一致,并与对应5 cm深温度和湿度的变化曲线基本吻合;土壤呼吸大小具有一定的差异:在整个观测期内,10年生,16年生,27年生银合欢林土壤呼吸平均排放速率分别为1.96、1.74、1.60 μmol·m-2·s-1,土壤呼吸排放速率呈随着林龄的增大而减小的趋势。10年生,16年生,27年生银合欢林土壤温度的月温差分别为:7.05、6.95、7.23°C(图2b);湿度变化范围随着林龄增大而增大,分别为:26.18~30.78、24.15~30.40、25.99~34.50% (图 2c)。
图2 不同林龄银合欢土壤呼吸、温度、湿度生长季变化Fig.2 The variations of soil respiration rate(a),soil temperature(b)and soil moisture(c)in different ages of Leucaena leucocephala(Lam.)de Wit in Augest
2.2 不同林龄银合欢土壤呼吸与土壤5cm深温度和水分的关系
研究区域各个林龄的5 cm深土壤呼吸速率对土壤温度的变化十分敏感,二者呈显著正相关指数函数关系(p<0.05)(表 2),R2介于 0.84~0.99 之间,在 3 种林龄中,16年生银合欢土壤呼吸与温度的拟合程度最好。10、16、27年生银合欢土壤呼吸的Q10值分别为:1.48,2.14,2.25,表明在观测期内,土壤呼吸对温度的依赖程度随着林龄的增大而增大。
影响土壤呼吸的另一重要指标是土壤湿度;研究区所属的干热河谷降雨主要集中在夏秋季,而降水是土壤水分最主要的来源。在整个观测期内,不同林龄银合欢林地土壤CO2排放速率与土壤5 cm处湿度均存在显著的线性相关(p<0.05)。但土壤呼吸与水分的相关性小于温度。
2.3 根系和枯落物对银合欢土壤呼吸的影响
整个生长季观测周期内,不同林龄银合欢林3种处理土壤呼吸速率波动趋势相似。总体表现为:阻根处理测的土壤呼吸速率最小,保留凋落物处理的土壤呼吸速率大于去凋落物处理(图3 a、b、c)。分析根系与枯落物对土壤呼吸的影响发现:3种林龄土壤呼吸均随着林龄增大呈现出不断减小的趋势,而根系呼吸和枯落物呼吸对总呼吸的贡献则随着年龄增大而增大,其中根系对土壤呼吸的贡献按林龄增大顺序分别为:16.8%、43.5%、47.7%,枯落物对土壤呼吸的贡献按林龄增大顺序分别为31.7%、34.2%、42.6%(图3d)。
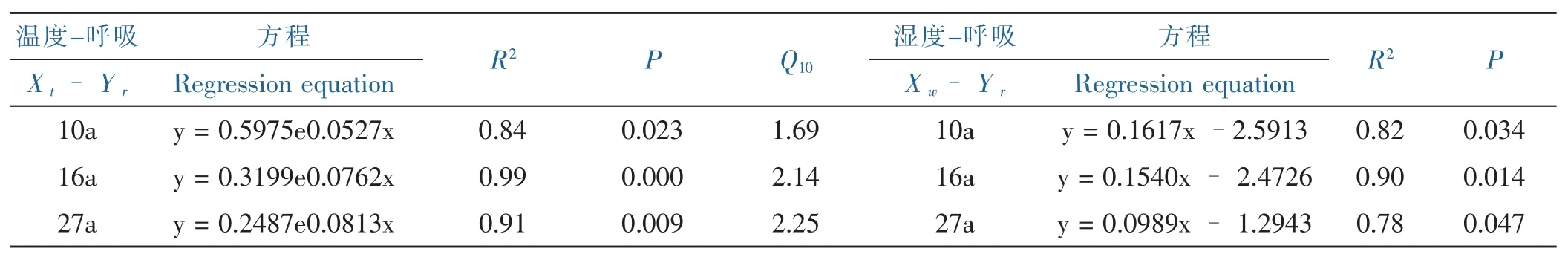
表2 不同林龄银合欢土壤呼吸与土壤温度、土壤呼吸与湿度的关系Table 2 Correlation of soil respiration to soil temperature and soil moisture in different ages of Leucaena leucocephala(Lam.)de Wit
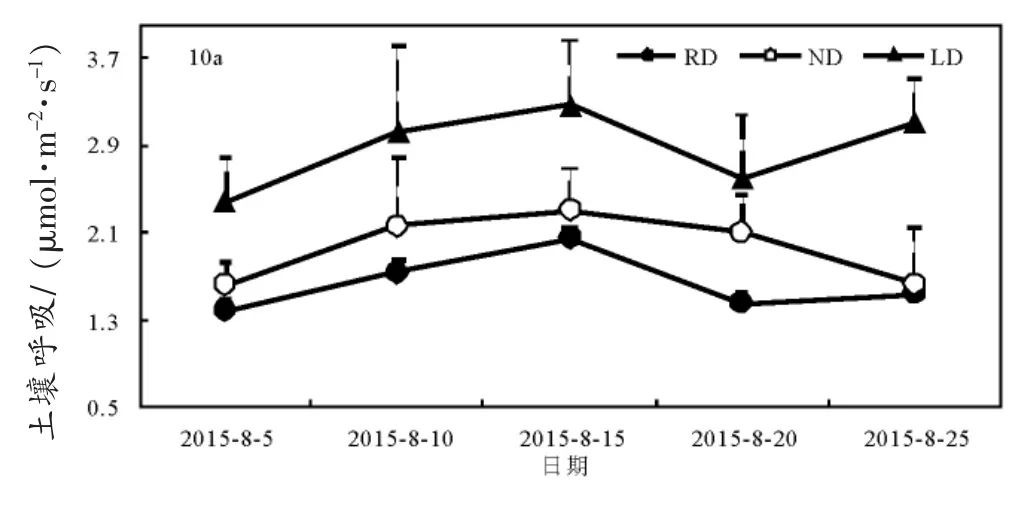
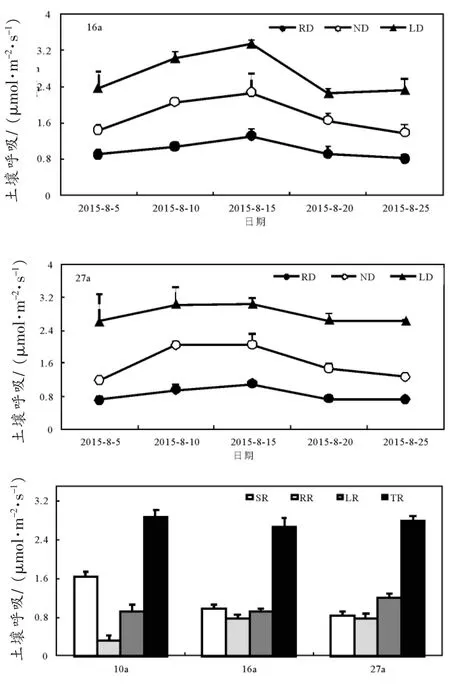
图3 去根和加凋落物处理对不同林龄土壤呼吸的影响Fig.3 Effects of root exclusion and litter treatment on soil respiration indifferentagesof Leucaenaleucocephala(Lam.)de Wit
2.4 土壤理化性质对银合欢土壤呼吸的影响
分析不同林龄银合欢林地的土壤理化性质可发现(表3):不同林龄银合欢林孔隙度表现为随着林龄变大而减少的趋势,且10年生与16年生、27年生间有显著差异性(p<0.05),表明10年生银合欢林地的土壤通气状况、土壤松紧度优于16年生和27年生银合欢林地。不同林龄银合欢林地土壤pH值均呈弱碱性,10年生pH最小但三者间无显著差异。土壤有机质、全氮、水解氮、过氧化氢酶与蔗糖酶含量随着林龄增长均呈下降趋势,除水解氮含量外,10年生均显著大于16年、27年生。
通过Pearson相关性分析对土壤呼吸、土壤理化性质动态变化规律进行分析可得(表4),土壤呼吸与过氧化氢酶、蔗糖酶、土壤有机质、全氮和孔隙度间均存在显著正相关关系(p<0.05),而水解氮则与土壤呼吸没有显著的相关性(p>0.05)。
3 结论与讨论
3.1 水热条件对土壤呼吸的影响
前人研究表明,土壤呼吸的变化是温度和水分共同作用的结果[7]。温度升高可导致植被生物量增加,土壤碳含量升高,进而影响土壤呼吸,二者存在显著相关性,主要关系模型有线性模型、指数模型、二次方程、乘幂模型等[19]。本研究中,土壤呼吸与土壤温度的月变化曲线基本一致,采用指数型模型对其进行模拟,结果显示同一林龄的土壤呼吸速率与5 cm深土壤的温度存在显著正相关(p<0.05)关系,符合前人的研究结果[20-21]。不同林龄银合欢林地在观测期内的温差较小(分别为 7.05、6.95、7.23),而 10 年生,16 年生,27年生银合欢的温度对土壤呼吸敏感性Q10值分别为1.69、2.14、2.25,这表明老林龄对温度变化的响应较为敏感,造成老龄林银合欢Q10高于中幼龄林的原因:可能是老龄林枯落叶堆积厚,温度波动较小,使得土壤呼吸更多地受到土壤温度的限制,即中老龄林银合欢土壤呼吸对温度反应敏感。
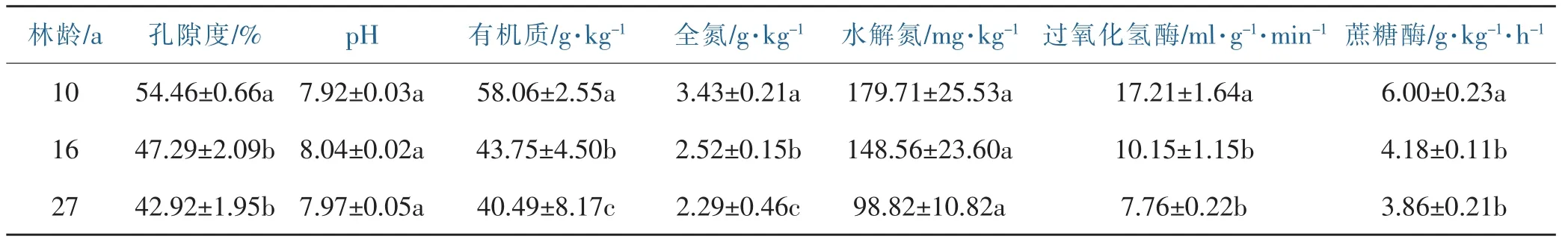
表3 不同林龄银合欢土壤理化性质Table 3 soil physical and chemical properties in Leucaena leucocephala(Lam.)de Wit forests of different age

表4 银合欢林地土壤呼吸与土壤理化性质的关系Table 4 Correlation of soil respiration and soil physical and chemical properties in different ages of Leucaena leucocephala(Lam.)de Wit
影响森林生态系统土壤表面CO2通量的另一个重要环境因子是土壤水分,它通常影响植物的根系分布、呼吸作用以及微生物活性[22]。前人研究发现,当土壤水分在土壤持水量范围内,尤其在土壤水分是影响土壤呼吸主要胁迫因子的区域,如干旱或半干旱地区,二者呈现显著正相关关系[23]。本研究发现不同林龄银合欢林地土壤呼吸与其对应5 cm深的湿度呈现显著正相关,土壤呼吸与土壤湿度的月变化也曲线基本一致,说明干热河谷地区林地土壤湿度也是土壤呼吸的主要影响因子。土壤含水量太低导致根系或微生物活动减弱,CO2将会减少;土壤水分过高使土壤孔隙减小,会影响O2进入与CO2排放[20]。本研究发现不同林龄土壤湿度表现出老龄林>幼中龄林的趋势,与土壤呼吸速率大小存在差异(幼中龄林>老龄林)(图1),这可能是因为老龄林土壤湿度更接近田间持水量,反而阻碍了土壤通透性,导致CO2和O2传输困难,土壤呼吸值偏小[24],但相关内容仍需进一步研究。
3.2 根系和枯落物对土壤呼吸的贡献
前人研究认为,凋落物层可为微生物、真菌的活动提供有机质,主导着土壤主要生物化学过程,凋落物的减少可引起土壤呼吸减弱[7,25]。从本研究整个观测周期来看:与正常处理比较,阻根处理降低了土壤呼吸速率,而保留凋落物处理则提高土壤呼吸速率。去根处理使土壤呼吸速率降低,根系呼吸按林龄从小到大依次贡献:16.8%、43.5%、47.7%(图3),与北温带阔叶林和樟树生长季根系呼吸贡献率大小基本一致[26-27]。植物在生长旺季时,较强的光合作用可为根系提供较多的物质和能量,刺激根部生长,从而增大了根系CO2的排放;然而,阻根处理不仅切断了植物生理活动的呼吸,还切断根际微生物养分的来源(根系能分泌易被微生物利用的有机质和改变pH、土壤结构、水分等土壤物理和化学环境),导致根际微生物数量及其酶活性的下降,使土壤呼吸速率大幅下降[28],因此,去根处理对根系较为发达的中老龄林影响更大。在整个土壤呼吸观察周期,保留凋落物处理显著增加土壤的CO2排放,这与Sulzman研究得出的凋落物能增加土壤呼吸的结论一致[29]。Riach等[30]得出全球森林生态系统中枯枝落叶对土壤呼吸的平均贡献率为33%,本研究发现枯落物处理对幼、中、老林龄的土壤呼吸贡献率分别为31.7%、34.2%、42.6%,老龄林的贡献率与Riach的研究结果存在较大差异,这可能由以下两个原因造成:1)银合欢老龄林根系过多,呼吸作用强于幼、中龄林;2)枯落物的增加引起CO2浓度与温度的升高,而厚重的枯落物又使老龄林土壤降温慢,吸引更多种类和数量的土壤微生物和真菌,增强了土壤呼吸作用。
3.3 土壤呼吸与土壤养分和酶的关系
林地的生长发育需要松紧适宜的土壤环境,而土壤环境易受外界因素影响[31]。本研究表明,银合欢林地土壤呼吸与土壤孔隙度存在显著正相关 (p<0.05);这是由于土壤是多孔系统,土壤微生物等呼吸所释放的CO2多聚集在孔隙当中,进而扩散到大气中,从而表观出土壤呼速率受CO2释放通道顺畅程度的影响[32]。土壤孔隙度呈现出随着林龄增大而减小的趋势,这可能是由于银合欢密集的细根导致土壤紧实度增加,使孔隙度、水稳性团聚体减少,引起土壤透气性下降,从而阻碍了气体的顺畅排放,最终导致呼吸排量的降低[33]。
土壤理化特性可以调控土壤呼吸。在土壤温度和湿度相对稳定时,土壤有机质是微生物的物质基础,对土壤的碳通量起决定性作用。土壤有机质含量影响土壤通风性、湿度和阳离子保持能力,为土壤微生物进行分解过程排放二氧化碳提供物质基础,一般而言,土壤有机质含量与土壤呼吸呈正相关[23,34-36],这与本研究得到的结果一致。土壤中氮是植物氮素营养的主要来源,有机态的氮经矿化作用转化为铵,被植物利用并释放二氧化碳,在温度水分稳定的情况下,氮含量的高低将影响土壤呼吸速率和土壤呼吸动态变化[37]。土壤中氮含量与微生物活性间也存在显著的相互作用关系,并最终影响土壤CO2的排放,因此土壤呼吸速率与全氮、水解氮呈显著正相关[38],本研究证实银合欢林地土壤全氮与土壤呼吸呈现显著正相关(p<0.05),但水解氮对土壤呼吸却无显著相关;这说明在干热河谷区水解氮并非影响植物根系和土壤微生物代谢活动的主要成分。另外,本研究还发现,不同土壤有机质、全氮、水解氮含量以及微生物过氧化氢酶、蔗糖酶活性皆随着林龄增大而减小,且幼龄林与老龄林间存在显著差异(除水解氮外)。银合欢为了获取更多的养分和水分以及应对松散的土壤状况,会延伸更多的细根来适应松散、贫瘠、干旱的土壤环境[12],这种现象在中老林龄银合欢林地中较为普遍,这可能是导致老龄林土壤中有机质、全氮下降的原因。
本研究仅对短周期内不同林龄银合欢林地生长季土壤呼吸与土壤理化性质之间的关系进行研究,下一步将继续对其长期跟踪调查,以期为解释泥石流频发区人工植被定植、存活、生长提供一定的依据与参考。
