核桃内果皮苯丙氨酸解氨酶基因克隆及表达分析
2018-11-30郭永翠张建良
郭永翠,朱 玲,高 山,张建良,张 锐,2*
(1.塔里木大学植物科学学院, 新疆 阿拉尔 843300; 2.塔里木大学新疆生产建设兵团塔里木盆地生物资源保护利用重点实验室, 新疆 阿拉尔 843300; 3.新疆伊犁州农业科学研究所, 新疆 伊宁 835000; 4.新疆生产建设兵团第一师三团, 新疆 阿拉尔 843300)
【研究意义】核桃(JuglansregiaL.)在中国栽培历史悠久,因其富含蛋白质、脂肪、碳水化合物等人体所必需物质及铁、锌、锰等多种微量元素,对于延缓人体衰老、心血管的保健作用、预防慢性疾病等具有非凡意义,有着极佳的保健效果和药用价值,在国内国际市场均具有巨大的消费潜力[1-4]。【前人研究进展】新疆核桃壳薄、早实丰产、出仁率高且种仁饱满,其中纸皮核桃品质尤为突出,成为近年成为消费热点[5]。‘纸皮’核桃壳厚小于1.0 mm (大多在0.5 ~0.8 mm),出仁率达65.0 %以上,种仁饱满,内果皮发育完整,其中‘温138’核桃是‘纸皮’核桃栽培园中发现的突变株,其平均出仁率高达83.2 %,产量极高,果壳极薄、极易脱皮,但壳面(内果皮发育不完整)开裂导致露仁,是适于核桃深加工的优质种质资源[6-7]。木质素(lignin)是核桃内果皮(硬壳)的主要成分且是内果皮中难分解的大分子有机物质,其沉积出现在次生加厚的细胞壁中以提高细胞壁的强度及硬度,对核桃内果皮发育影响较大[8-9]。苯丙氨酸解氨酶(phenylalanine ammonia-lyase,简称PAL)存在于一些菌类、藻类及所有绿色植物中,是参与木质素生物合成过程中苯丙烷类代谢途径的首个关键酶,在多种植物的木质化组织中含有较高PAL活性,PAL基因对木质素的合成起重要作用[10-11]。目前已从辣椒(CapsicumannuumL.)[12]、当归(Angelicasinensis)[13]、马尾松(Pinusmassoniana)[14]、黄瓜(CucumissativusL.)[15]等多种植物中成功获得了PAL基因并展开相关研究。【本研究切入点】经研究发现,植物中PAL基因对木质素的合成起重要作用,在转基因烟草(NicotianatabacumL.)植株中抑制PAL的表达将导致木质素等相关次生代谢产物含量的下降,但植株生长出现不正常现象[16]。不同PAL基因在拟南芥(Arabidopsisthaliana)植株中各个部位的表达量不同,表明不同的PAL基因具有不同生物活性和表达部位[17];其在不同品种龙眼(EuphorialonganL.)叶片中的表达量存在显著差异[18];且在独一味(Lamiophlomisrotata)的不同组织中表达有差异,表达量由高到低分别是叶片、根部、叶柄、茎[19]。【拟解决的关键问题】目前关于‘温138’核桃露仁现象及其与之相关的核桃内果皮硬化过程中木质素沉积的系统分子机理尚不明确,本研究以‘温138’、‘纸皮’核桃为研究对象,对其内果皮中木质素相关PAL基因进行克隆及生物信息学分析,为运用基因工程改良核桃果实品质提供理论依据。
1 材料与方法
1.1 材料
供试材料露仁种质‘温138’核桃及硬壳发育完整‘纸皮’核桃,均采自新疆维吾尔自治区温宿县核桃木本粮油林场(图1)。在果实膨大期后期6月7日(约花后51 d)至硬核期7月26日(约花后100 d)采样,采样覆盖整个核桃内果皮硬化期。将核桃内果皮剥离存于-80 ℃超低温冰箱中。
植物总RNA提取采用E.Z.N.A Plant RNA特殊植物RNA提取试剂盒(Omega公司);cDNA第一链合成采用EasyScript One-Step gDNA Removal and cDNA Synthesis SuperMix(100rxns)试剂盒(全式金公司);胶回收采用Easy Pure®Quick Gel Extraction试剂盒(全式金公司);qRT-PCR检测采用SYBR®Premix ExTaqTMGC(Perfect Real Time染料法)试剂盒(TaKaRa公司)。
1.2 核桃内果皮总RNA提取
利用植物总RNA提取试剂盒提取核桃内果皮中总RNA。用1 %琼脂糖凝胶电泳方法检测及鉴定PCR产物的浓度、纯度和亮度。采用cNDA合成试剂盒反转录合成cDNA。
1.3 引物设计
根据核桃转录组数据库和NCBI数据库已发表的核桃PAL基因序列(登录号:JX069977.1)设计引物,‘温138’核桃引物PAL-F1:5′-CCCCTCCAAACAACCCAA-3′及PAL-R1:5′-CCTTCAGACAATCCAGCA-3′,‘纸皮’核桃引物PAL-F2:5′-ACCCAATCCAAGAAAGAA-3′及PAL-R1:5′-CCTTCAGACAATCCAGCA-3′。引物由上海生工生物技术有限责任公司合成。

A:‘温138’核桃硬壳发育情况, B: ‘纸皮’核桃硬壳发育情况A: ‘Wen138’ walnuts shell development, B: ‘Zhipi’ walnuts shell development图1 ‘温138’和‘纸皮’核桃硬壳发育情况Fig.1 ‘Wen138’ walnuts and ‘Zhipi’ walnuts shell development
1.4 核桃内果皮PAL基因的克隆及测序
RT-PCR反应体系(20 μl)及条件为:模板3 μl,上、下游引物(10 μM)各0.5 μl,2×EasyTaq®PCR SuperMix 10 μl,最后加灭菌ddH2O6μl。扩增条件:94 ℃/3 min(预变性);94 ℃/45 s(变性),57 ℃/45 s(退火),72 ℃/2 min(延伸),31个循环;72 ℃/8 min(继续延伸);4 ℃保存。用1 %琼脂糖凝胶电泳检测PCR产物并成像,后将扩增的目的条带进行回收并连接到T载体,筛选出阳性克隆测序。
1.5 生物信息学分析
1.5.1 核苷酸、氨基酸序列分析 将阳性克隆的测序结果及推导氨基酸序列采用NCBI数据库BLAST功能(https://blast.ncbi.nlm.nih.gov/Blast.cgi)进行在线PAL序列匹配度、同源性分析;ORF Finder (https://www.ncbi.nlm.nih.gov/orffinder/)检测cDNA序列的开放阅读框;PAL基因推导氨基酸序列采用Clustal X 1.08软件进行多序列比对;PAL蛋白功能结构域利用NCBI数据库Conserved Domains (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)进行在线分析。
1.5.2 理化性质分析 运用ExPASy数据库ProtParam功能(http://web.expasy.org/protparam/)分析核桃内果皮WJ-PAL蛋白及ZJ-PAL蛋白的相对分子质量、氨基酸数量及理论等电点等相关理化性质。
1.5.3 蛋白质二级结构及三维结构预测 运用EBI数据库InterProScan功能(http://www.ebi.ac.uk/interpro/search/sequence-search)在线分析及预测核桃内果皮PAL蛋白保守区域、采用PredictProtein在线平台(https://www.predictprotein.org/)分析PAL蛋白二级结构,选用在线同源服务软件SWISS-MODEL (http://swissmo del.expasy.org/)预测PAL蛋白质三维结构。
1.5.4 系统进化树 采用NCBI数据库BLAST在线比对,选取与克隆基因相似性较高的植物,采用MAGA5.1软件(Neighbor-Joining法)构建PAL基因与其它相似性高植物的系统进化树。
1.6 PAL基因的表达分析
以18S为内参,采用实时荧光定量PCR (qRT-PCR)检测PAL基因的相对表达量。根据已克隆的核桃内果皮木质素PAL基因序列,按照标准荧光定量PCR引物原则,分别设计‘温138’、‘纸皮’核桃PAL基因荧光定量PCR引物,露仁种质‘温138’核桃PCR引物PAL-WF:5′-AGGTGAAACGCATGGTGG-3′,PAL-WR:5′-CCTGTGATATGGTCAGGGTCTT-3′,硬壳完整种质‘纸皮’核桃PCR引物PAL-ZF:5′-CCGTTGTTGACGCTGTTTAT-3′,PAL-ZR:5′-GGGAG ACCCTGACCATTTC-3′,内参基因引物:NBQ-F:5′-AGTCGTAACAAGGTTTCCGTAGGT-3′;NBQ-R: 5′-GCTGGGCAGGTATCGACAAT-3′。以‘温138’及‘纸皮’核桃8个时期的cDNA为模板,采用两步法进行Real-Time PCR扩增,反应体系:cDNA模板1 μl,正、反引物(10 μM)各0.2 μl,SYBR®Premix ExTaqTMGC5 μl,最后加灭菌ddH2O至10 μl。扩增条件:95 ℃/30 s,95 ℃/5 s,40个循环;熔解曲线生成步骤:58 ℃/30 s,72 ℃/30 s,95 ℃/15 s,55 ℃/15 s;55 ℃/20 min;95 ℃/15 s。利用2-△△Ct方法[20]分别计算PAL基因在硬壳完整和露仁核桃内果皮发育过程中的表达量。
2 结果与分析
2.1 总RNA的提取及检测
核桃内果皮中总RNA凝胶成像呈现较清晰完整的28S rRNA和18S rRNA条带,5S rRNA条带较模糊,表明所得的总RNA有较好的完整性,28S rRNA的亮度是18S rRNA的两倍,测得核酸浓度A260/A280为1. 90~2. 20,说明提取的总RNA有较高的纯度(图2)。
2.2 核桃内果皮PAL基因的克隆
以‘温138’和‘纸皮’核桃内果皮总RNA反转录后的cDNA为模板,通过所设计的引物(PAL-F1、PAL-R1和PAL-F2、PAL-R1)进行RT-PCR反应(图3),得到露仁及硬壳完整核桃PAL基因序列,分别命名为WJ-PAL和ZJ-PAL(图4)。WJ-PAL基因片段长度为1000 bp (含有600 bp的ORF),可编码200个氨基酸,经分析发现其分子量为21.11 kD,理论等电点为8.44,WJ-PAL属于碱性蛋白;ZL-PAL基因片段长度为1000 bp (含有690 bp的ORF),可编码230个氨基酸,经分析发现其分子量为24.10 kD,理论等电点为7.44,ZJ-PAL属于弱碱性蛋白。
图2 电泳检测核桃内果皮总RNAFig.2 Electrophoresis of total RNA from walnut endocarp
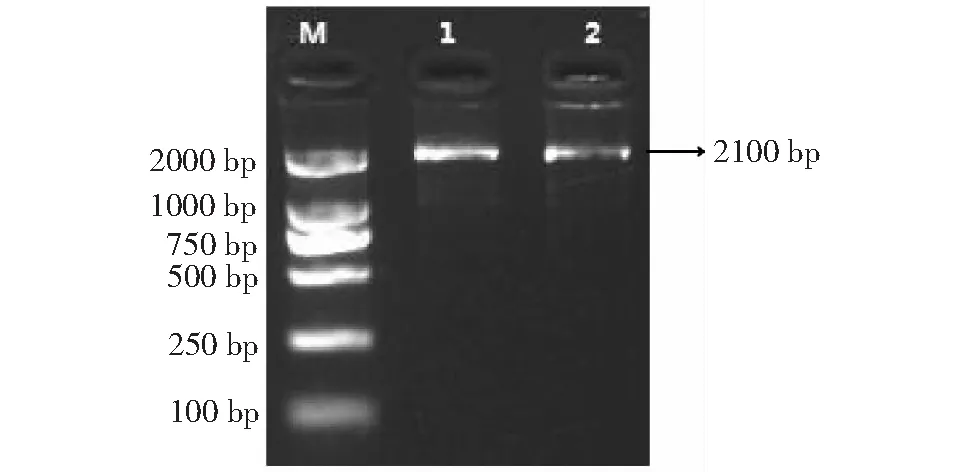
M: DL2000 DNA Marker; 1: ‘温138’核桃; 2: ‘纸皮’核桃M: DL2000 DNA Marker; 1: ‘Wen138’ walnut; 2: ‘Zhipi’ walnut图3 核桃内果皮PAL基因的保守区PCR产物Fig.3 PCR product of PAL gene conservative region in walnut endocarp
2.3 生物信息学分析
2.3.1 序列分析 将得到的测序结果与NCBI数据库己知植物PAL基因进行序列同源性比对的结果(图5)表明,WJ-PAL与ZJ-PAL核苷酸序列及氨基酸推导序列均与其它植物PAL基因序列有较高的匹配度。
将本研究获得的2个基因进行相似性分析,结果显示WJ-PAL和ZJ-PAL核苷酸序列与黑核桃、桃、日本杏等物种的PAL基因序列匹配度最高,其同源性均可达到77 %~96 %,其中均与黑核桃(AJ278454.1)的匹配度最高,WJ-PAL、ZJ-PAL与黑核桃同源性分别达到89 %、96 %。其相似性经NCBI数据库BLASTp在线分析可知,WJ-PAL和ZL-PAL
与其它植物推导氨基酸序列相似性均在79 %以上。PAL推导氨基酸序列发现WJ-PAL与ZL-PAL均存在Phenylalanine ammonia-lyase and histidine ammonia-lyase (PAL-HAL)、PPK09367、huth及HutH 4个非特异性位点,并且含有phe_am_lyase、芳香族裂解酶的Lyase_aromatic及PAL (PLN02457) 3个功能结构域,WJ-PAL与ZL-PAL为多结构域蛋白,属于裂解酶I (Lyase_I_Like)超家族(图5)。
2.3.2 系统进化树分析 将本研究获得的两个基因核苷酸序列进行多重序列比对(图6),露仁种质‘温138’克隆所得WJ-PAL基因和硬壳完整品种‘纸皮’克隆所得ZJ-PAL基因序列一致性显示只有54.76 %。
经过在线比对,发现核桃WJ-PAL及ZJ-PAL基因与其它植物的PAL基因具有较高的相似性,选取与WJ-PAL及ZJ-PAL相似性较高的十九种植物构建核桃PAL基因与其它植物PAL基因的系统进化树(图7)。结果显示,核桃与乔木类植物亲缘关系较近,‘纸皮’核桃与黑核桃聚为一支,川杨、颤杨、胡杨、毛果杨等杨树较好的聚为一支,盐肤木与芒果这两个漆树科植物聚为同一支,芸香科柑橘属的柠檬与甜橙聚为同一支,葡萄科的葡萄与桑科的桑树较好的聚为一支,并且自展值均达到100,其它植物也各自与相近的植物聚为一支。硬壳完整核桃‘纸皮’(ZJ-PAL)与黑核桃(Juglansnigra)聚在一起,却与露仁核桃种质‘温138’ (WJ-PAL)未能聚为同一支,这说明就PAL基因而言,硬壳完整核桃‘纸皮’与黑核桃聚为一支说明其亲缘关系较近,而露仁种质‘温138’核桃却未与黑核桃及其它同源植物聚在一起说明有较远的亲缘关系。

A: WJ-PAL基因片段测序; B: ZJ-PAL基因片段测序A: Sequencing result of WJ-PAL gene fragment; B: Sequencing result of ZJ-PAL gene fragment图4 核桃PAL基因片段测序Fig.4 Sequencing result of PAL gene fragment in walnut

A: WJ-PAL基因的结构域分析; B: ZJ-PAL基因的结构域分析A: Domain analysis of WJ-PAL gene; B: Domain analysis of ZJ-PAL gene图5 PAL基因的结构域分析Fig.5 Domain analysis of PAL gene

图6 WJ-PAL及ZJ-PAL基因多重序列比对Fig.6 Multiple sequence alignment of WJ-PAL and ZJ-PAL gene
2.4 蛋白质二级结构分析
提交WJ-PAL、ZJ-PAL蛋白的氨基酸序列进行在线预测分析(图8)。分析得到,WJ-PAL蛋白含有58.29 %的无规则卷曲,是WJ-PAL蛋白二级结构中最大量的结构元件,其次含有35.68 %的α-螺旋和6.02 %的延伸链等结构元件,N-末端存在较多的无规卷曲结构,而C-末端则存在较多的延伸链结构。ZJ-PAL蛋白主要含有48.03 %的α-螺旋,是ZJ-PAL蛋白二级结构中最为重要的结构元件,其次含有46.03 %的无规则卷曲结构和5.68 %的延伸链。
在线预测WJ-PAL和ZJ-PAL蛋白的保守区域,结果表明这两个蛋白属于L-天冬氨酸家族(L-Aspartase-like),WJ-PAL和ZJ-PAL蛋白保守区域包含芳香族氨基酸裂解酶(IPR001106)活性位点。其中ZJ-PAL蛋白具有PAL-HAL信号(含有PAL-HAL活性位点),具备苯丙氨酸解氨酶(PAL)家族蛋白的特征。
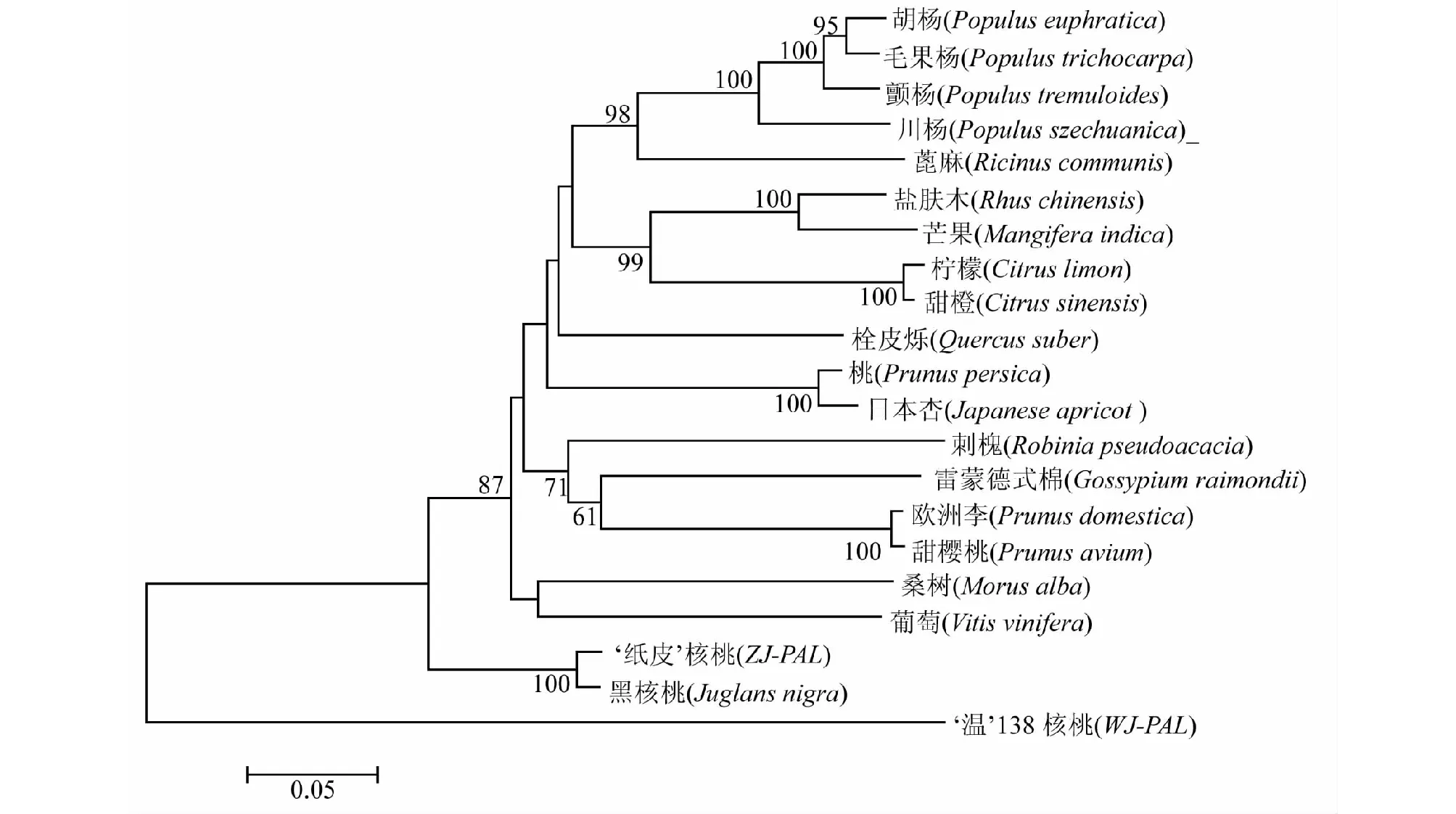
图7 核桃与18种植物PAL基因系统进化树分析Fig.7 Phylogenetic tree of PAL gene in walnut and 18 other plants

A: WJ-PAL蛋白二级结构; B: ZJ-PAL蛋白二级结构; 红色:α-螺旋; 蓝色:延伸链A: Secondary structure of WJ-PAL protein; B: Secondary structure of ZJ-PAL protein; Red: α-helix; Blue: Extended strand图8 PAL蛋白二级结构Fig.8 Secondary structure of 7 PAL protein
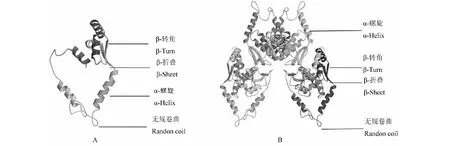
A: WJ-PAL蛋白三级结构; B: ZJ-PAL蛋白三级结构A:The tertiary structure of WJ-PAL protein; B: The tertiary structure of ZJ-PAL protein图9 PAL蛋白三级结构Fig.9 The tertiary structure of PAL protein

A:WJ-PAL基因相对表达量; B: ZJ-PAL基因相对表达量; W: ‘温138’核桃; Z:‘纸皮’核桃; W(Z)67: 花后51 d (6月7日); W(Z)613: 花后57 d (6月13日); W(Z)621: 花后65 d (6月21日); W(Z)628: 花后72 d (6月28日); W(Z)74: 花后78 d (7月4日); W(Z)712: 花后86 d (7月12日); W(Z)720: 花后94 d (7月20日); W(Z)726: 花后100 d (7月26日)A: Relative expression of WJ-PAL gene; B: Relative expression of ZJ-PAL gene; W: ‘Wen 138’ walnut; Z: ‘Zhipi’ walnut; W (Z) 67: 51 days after anthesis (June 7th); W (Z) 613: 57 days after anthesis (June 13th); W (Z) 621: 65 days after anthesis (June 21st); W (Z) 628: 72 days after anthesis (June 28th); W (Z) 74: 78 days after anthesis (July 4th); W (Z) 712: 86 days after anthesis (July 12th); W (Z) 720: 94 days after anthesis (July 20th); W (Z) 726: 100 days after anthesis (July 26th)图10 核桃内果皮硬化过程PAL基因相对表达水平Fig.10 Relative expression level of PAL gene during hardening of walnut endocarp
2.5 蛋白质三级结构预测
分别建立WJ-PAL与ZJ-PAL蛋白三维空间结构模型(图9)。WJ-PAL与ZJ-PAL这两种蛋白质三级结构由α-螺旋、β-转角、β-折叠和无规则卷曲构成,其中均α-螺旋为主要结构元件,其它元件较少。蛋白三级结构分析可知ZJ-PAL较WJ-PAL结构复杂的多,其α-螺旋、β-转角、β-折叠和无规则卷曲结构等元件含量多。
2.6 WJ-PAL与ZJ-PAL基因相对定量分析
实时荧光定量PCR技术分析WJ-PAL与ZJ-PAL在核桃内果皮硬化过程中的表达水平(图10)。WJ-PAL基因相对表达量在‘温138’核桃的8个不同发育时期中有非常明显的差异。相对表达量最高的花后100 d (117.78)是相对表达量最低的花后51 d (1.00)的117倍。这表明,‘温138’核桃内果皮硬化过程的前期PAL基因表达量相对较小,至核桃内果皮硬化过程的后期其PAL基因大量表达。在核桃内果皮硬化过程的8个不同发育时期ZJ-PAL基因的相对表达量相较WJ-PAL差异较小,在这8个时期中,相对表达量最高的花后65 d (1.33)是相对表达量最低的花后72 d (0.25)的5倍。PAL基因在露仁核桃‘温138’内果皮中前期木质素相对表达量均较低,于花后86 d开始增长,在花后100 d达到最高值,是前期的近100倍,硬壳完整核桃‘纸皮’在硬核期整个时期木质素相对表达量变化较平缓,于花后72 d最低,最高出现在花后65 d,整个种皮发育过程中,PAL基因在硬壳完整核桃内果皮中的表达量低于同期露仁核桃内果皮中的表达量。
3 讨 论
PAL能够催化高等植物中苯丙氨酸解氨生成氨气和反式肉桂酸,是木质素单体合成过程中所需的第一步反应酶[21],其受环境影响较小可用作物种鉴定[22]。目前关于露仁核桃在PAL基因克隆及表达分析方面研究还未见文献报道,PAL对核桃露仁现象的作用机理还不得而知。本试验克隆得到露仁核桃种质‘温138’及硬壳发育完整核桃种质‘纸皮’内果皮PAL基因,分别命名为WJ-PAL与ZJ-PAL,利用生物信息学预测获得PAL基因的理化性质参数、二级结构、三级结构等,分析PAL基因的进化关系及在核桃内果皮硬化过程中不同时期中的相对表达特性。
WJ-PAL与ZJ-PAL基因均为苯丙氨酸解氨酶基因的家族成员,根据试验结果得知,WJ-PAL与ZJ-PAL推导氨基酸序列存在相同的非特异性位点(PPK09367, HutH, hutH及PAL-HAL)及功能结构域(PLN02457, phe_am_lyase及Lyase_aromatic),这一结果白桦(Betulaplatyphylla)、美洲南瓜(Cucurbitapepo)相关研究结果吻合[11, 23]。同源性分析显示核桃露仁种质WJ-PAL基因与其它植物没有聚为一支,而硬壳发育完整ZJ-PAL基因与同科的植物较好地聚在一起,其与乔木类植物亲缘关系较近,其中与黑核桃亲缘性最高。WJ-PAL与ZJ-PAL核苷酸序列及其编码的氨基酸序列与黑核桃PAL基因及其氨基酸序列的相似性分别为89 %和99 %。发现不同植物间PAL基因也有较高的一致性,推测PAL在核桃发育过程中对内果皮(硬壳)完整程度有较大的影响。
本试验实时荧光定量PCR结果显示,露仁‘温138’核桃WJ-PAL在内果皮硬化的8个不同发育时期基因的表达量存在明显差异,其中前期的相对表达量均较低,在核桃硬核期的花后100 d达到最大值,是前期的近100倍,对照‘纸皮’核桃ZJ-PAL在内果皮硬化的8个时期基因表达量差异较小,在花后51~100 d间相对表达量变化较小,在花后71 d出现最低值,这与薛应钰的研究有一定的出入。美洲南瓜(Cucurbitapepo)研究发现木质素含量随种子的成熟显著降低,其种仁裸露裸仁现象是由于种皮发育过程中厚壁组织中缺乏木质素的合成和积累;时空表达研究发现,裸仁美洲南瓜在授粉后的8 ~18 d种皮中随木质素的合成和积累PAL酶活性逐渐升高,随后PAL酶活性明显降低[24]。在自交授粉20 d后,PAL基因在硬壳完整美洲南瓜种皮中表达量上升,在裸仁美洲南瓜种皮中表达量则下降;其中在同一时期有壳美洲南瓜的表达量远高于裸仁美洲南瓜中的表达量[11]。本试验相对定量分析结果发现,PAL基因在硬壳完整核桃内果皮中的表达量低于同期露仁核桃内果皮中的表达量,本结果与美洲南瓜研究有较大的出入。说明在核桃生长发育过程中,PAL基因在露仁核桃中的表达量与酶活性具有一定同步性,‘温138’核桃内果皮出现露仁现象可能由于前期木质素代谢中间产物形成太少,进一步说明了露仁核桃内果皮形成的原因可能是由于前期缺乏木质素代谢产物的积累。本试验结果可以看出在硬核期木质素的改变对核桃内果皮发育存在一定影响,核桃出现露仁现象与PAL活性的关系还有待进一步深入研究。
4 结 论
通过基因克隆获得露仁核桃及硬壳完整核桃内果皮PAL基因,WJ-PAL与ZJ-PAL推导氨基酸序列存在相同的非特异性位点、功能结构域,均属于Lyase_I_Like超家族,黑核桃序列相似性极高。露仁核桃‘温138’前期PAL基因表达远低于后期表达,核桃露仁现象可能由于内果皮硬化过程前期木质素积累不足造成。
