人睾丸特异表达基因PIAS-NY原核表达和多克隆抗体的制备*
2018-11-29王海燕曾凡强王玉兰郑英戚大石
王海燕,曾凡强,王玉兰,郑英,戚大石
(1.徐州医科大学 人体解剖学教研室,江苏 徐州 221004;2.扬州大学 组胚教研室,江苏 扬州 225001;3.徐州医科大学 遗传学教研室,江苏 徐州 221004)
睾丸特异表达基因的转录调控是精子发生过程中最基本的调节因素[1],具有进化上高度保守、生精细胞特异表达或高表达及阶段特异性表达的特征[2-3]。因此筛选并研究这些差异表达的基因有望发现与精子发生相关的新基因,或发现已知基因的新功能,研究结果对于揭示人睾丸发育/精子发生的分子机制、建立男性不育在分子水平的诊断方法、发现精子发生障碍的基因治疗手段和寻找男性避孕的新方法均具有重要意义。活化型信号转导及转录激活蛋白抑制蛋白(protein inhibitor of activated STAT, PIAS)家族属于这类蛋白。目前,真核哺乳动物中发现的PIAS家族成员主要包括PIAS1、PIAS2、PIAS3和PIASy[4-6]。PIAS蛋白家族在人睾丸组织中存在着多种调节机制,同时在睾丸功能中发挥调节作用[7]。
PIAS2包括PIASxα、PIASxβ和PIAS-NY 3个成员[1]。序列分析发现,PIAS-NY是PIASx的一个新的不同剪接体[7]。PIAS-NY mRNA在成人睾丸中高表达,在胚胎睾丸中低表达,而且成人睾丸中的表达水平是胚胎期的14倍;PIAS-NY mRNA特异表达于人睾丸及胰腺中,而在其他器官中未见表达[6]。提示PIAS-NY主要在精子发生过程中发挥重要作用。
PIAS-NY作为一种新发现的睾丸特异表达基因,同其他家族成员一样,具有典型的N端的SAP结构域、PINIT氨基酸基序及RLD结构域。目前,PIAS-NY相关研究非常少,为了阐述其在精子发生过程中具体作用机制,本研究构建PIAS-NY原核表达质粒,通过诱导大肠杆菌BL21(DE3)表达融合蛋白,纯化后获得高纯度的可溶性原核表达蛋白,多次免疫BalB/C小鼠并制备PIAS-NY多克隆抗体。本研究获得PIAS-NY全长开放阅读框,包含PIAS-NY全部结构域,有利于全面开展作用蛋白的深入研究。
1 材料与方法
1.1 材料
LA-Taq DNA聚合酶购自日本TaKaRa公司,小剂量质粒抽提试剂盒和琼脂糖凝胶切胶纯化试剂盒购于北京BioTeke公司,T4连接酶及质粒pET30a购自美国Promega公司,限制性内切酶EcoRI和XhoI均购自宝生物工程(大连)有限公司,Freund完全佐剂和不完全佐剂购自美国Sigma公司,His-Binding-Resin购自上海悦克生物科技有限公司,透析袋购自美国Solarbio公司,改良Eagle培养基(Dulbecco's modification of Eagle's medium Dulbecco, DMEM)购自美国Hyclone公司,新生胎牛血清购自美国Gibco公司,兔抗鼠RUVBL2单克隆抗体购自美国Abcam公司,辣根过氧化物酶标记山羊抗小鼠IgG和辣根过氧化物酶标记山羊抗兔IgG购自北京中杉金桥有限公司,RIPA裂解液购自武汉博士德生物公司,免疫共沉淀试剂盒购自瑞士Roche公司,超敏化学发光(Enhanced Chemiluminescence, ECL)试剂盒购自上海碧云天生物技术有限公司。人精原cDNA文库、GC2细胞和293T细胞均由扬州大学郑英教授惠赠。
1.2 方法
1.2.1 目的基因的获得 PIAS-NY扩增引物根据Gen Bank基因序列,用Primer 5.0软件设计,PIAS-NY基因正向引物5'-GGGGAATTCGGATGGCGGATTTCG AAGAGTTG-3'(划线部分为EcoRI酶切位点),反向引物5'-GGGCTCGAGTTACCCATCTAATATTAGA CTTTC-3'(划线部分为XhoI酶切位点)。50 μl PCR反应体系:10×LA Tap Buffer 5 μl,dNTP(10 mmol/L)1 μl,Primer F 2 μl,Primer R 2 μl,模板 2 μl,高保真Taq酶0.5 μl,H2O 37.5 μl,反应条件:95℃预热5 min,95℃变性 30 s,58℃退火60 s,72℃延伸90 s,35个循环,72℃延伸10 min,4℃恒温16 h。1.5%琼脂糖凝胶电泳,45 min,割胶回收PIAS-NY片段,大小为1 275 bp。
1.2.2 PIAS-NY-pET30a原核蛋白的诱导表达 将0.5 μl PIAS-NY-pET30a质粒DNA转化到大肠杆菌BL21(DE3)感受态细胞中,37℃ LB/Kan+固体平板上培养过夜。次日,挑选单克隆菌落于10 ml LB液体/Kan+,225 r/min,37℃,过夜。用过夜菌液将250 ml LB液体/Kan+OD调至0.3左右,225 r/min,37℃ 3 h,测得光密度(optical density, OD)值为0.5左右,取出1 ml菌液作为空白对照,剩余菌液中加入终浓度1 mmol/L异丙基-β-D-硫代半乳糖苷(isopropyl-β-D-thiogalactoside, IPTG),两者同时 225 r/min,37℃4 h,分别取出10 μl菌液进行十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis, SDS-PAGE),确定BL21菌株是否表达融合蛋白。阳性菌液3 500 r/min,4℃ 20 min,去除上清液,置入-20℃冰箱冷冻保存。向沉淀中加入 15 ml Binding Buffer A[20 mmol/L Tris-Cl(pH 7.9),0.5 mol/L NaCl,10%甘油]重悬,冰上间断超声裂解菌体(功率25%、超声5 s、间歇5 s,总工作时间600 s),3 500 r/min、4℃、10 min,分别收集上清液和沉淀,向沉淀中加入15 ml Binding Buffer B(破菌缓冲液B:破菌缓冲液A加入8 mol/L尿素),继续超声悬浮液(功率25%、超声5 s、间歇5 s,总工作时间600 s),3 500 r/min、4℃、10 min,分别收集蛋白上清液和沉淀。SDS-PAGE电泳,考马斯亮蓝染色,确定PIAS-NY-pET30a原核表达融合蛋白以哪种形式存在于大肠杆菌中。
1.2.3 透析袋预处理 将透析管裁剪成30 cm小段,将2%(m/V)碳酸氢钠和1 mmol/L 乙二胺四乙酸(ethylene diamine tetraacetic acid, EDTA)(pH=8.0)溶液煮沸,透析袋浸泡其中煮沸10 min;用蒸馏水彻底漂洗;1 mmol/L EDTA(pH=8.0)溶液煮沸,透析袋浸泡煮沸10 min;室温冷却,蒸馏水彻底漂洗,4℃蒸馏水完全浸泡保存。整个过程全部带手套操作,使用前必须仔细检查,不漏方可使用。
1.2.4 融合蛋白纯化及复性 用His-Binding-Resin对收集融合蛋白进行纯化,以下整个过程均在4℃完成。安装好His-Ni柱,先用10 ml Binding Buffer B平衡去除穿透液体。将3 ml融合蛋白上清液加入纯化柱中,收集穿透夜,以观察吸附效果。加入蛋白样品,滴完后用2.5 ml Binding Buffer B洗柱1遍,尽量去除非特异性吸附的杂蛋白。依次用2 ml Eluting buffer洗脱(破菌缓冲液B中加入不同终浓度咪唑,浓度依次为5、25、50、100及200 mmol/L),控制滴速2~3 ml/h,分段收集洗脱液。用5 ml Binding Buffer B过柱,将上述收集的咪唑洗脱液进行SDS-PAGE电泳鉴定。发现100和200 mmol/L洗脱液中含有较高浓度目的蛋白,将100和200 mmol/L洗脱液混合液装入激活的透析袋中,4℃依次经过7、6、5、4、3及2 mol/L尿素缓冲液进行透析复性,各梯度浓度分别复性24 h,最后收集蛋白液体。用Bradford法测定蛋白浓度。以200 μl/管进行蛋白分装,置入-80℃冰箱冷冻保存。
1.2.5 免疫小鼠制备抗体 用苦味酸对20只20 g BalB/C小鼠进行随机标记,从1连续到20,随机挑选出3只作为阴性对照。取2管蛋白液体融化,分别加入等体积的Freund完全佐剂充分混合乳化后,按照100 μg/只蛋白剂量进行BalB/C小鼠腹腔注射,对照组注射等体积1×PBS缓冲液和Freund完全佐剂。2周后,取2管蛋白液体加等体积Freund不完全佐剂充分混合,按100 μg/只小鼠BalB/C小鼠腹腔注射。每隔2周加强1次,共4次。2个月后,将免疫好的小鼠眼球取血,收集血清。分装后作好标记,置入-80℃冰箱冷冻保存。
1.2.6 酶联免疫吸附测定(enzyme-linked immunosorbent assay, ELISA)抗血清滴度 将纯化PIAS-NY蛋白用包被液稀释至所需浓度5 μg/ml,96孔板中每孔加入100 μl,将96孔板放入湿盒,4℃过夜;次日,去除包被液,用PBS-T洗涤3次,5 min/次;加入含0.1%脱脂奶粉PBS-T,每孔150 μl,37℃封闭1 h;去除封闭液,PBS-T洗涤3次,5 min/次;按比例稀释PIAS-NY抗血清(1∶100、1∶1 000、1∶104、1∶105及1∶106),每孔加入100 μl,同时以BalB/C小鼠血清为阴性对照,37℃孵育70 min;PBS-T洗涤3次,5 min/次;二抗孵育:山羊抗小鼠IgG(1∶1 000倍稀释于PBS-T),每孔加入100 μl,37℃孵育60 min,PBS-T洗涤3次,5 min/次,加显色液100 μl,室温避光保存10 min,加入2滴终止液,终止反应,酶标仪读数(A490)。
1.2.7 免疫共沉淀 293T细胞铺6孔板,4℃预冷PBS轻轻洗涤细胞表面3次,每孔加入Roche免疫共沉淀试剂盒中的lysis buffer1 80 μl,冰上5~10 min,用刮棒将每孔内细胞裂解物刮干净,吸出于一EP管中,冰上30 min(每5 min上下颠倒混匀1次)。4℃13 000 r/min 15 min。将上清吸至另一EP管中,置于冰上。取出60 μl上清液液,剩余蛋白液体平均分为2份,每份分别加入鼠IgG和PIAS-NY抗血清各8 μl(或者兔IgG和RUVBL2单克隆抗体各8 μl)。4℃冰箱,滚筒上2 h。取80 μl protein G小珠子于一EP管中,10 000 r/min 20 s弃上清液,加500 μl lysis buffer1,混匀,10 000 r/min,弃上清液。2 h后将2份蛋白上清分别加入含小珠子的EP管中,混匀。4℃滚筒上过夜。次日,上下翻转,4℃ 10 000 r/min 20 s,弃上清。每管中加入500 μl lysis buffer 1洗涤珠子,上下翻转,4℃10 000 r/min 10 s,弃上清液。1 ml的wash buffer 2洗涤珠子,上下翻转,4℃10 000 r/min 10 s,弃上清液。1 ml的wash buffer 3洗涤珠子,上下翻转,4℃ 10 000 r/min 10 s弃上清液。加35 μl的1×loading buffer,混匀,煮沸5 min,上下翻转,4℃ 10 000 r/min 5 min收集上清液,即为兴趣蛋白-目的蛋白混合物。SDS-PAGE电泳后分别用兔源RUVBL2抗体免疫印迹检测。
2 结果
2.1 PIAS-NY-pET30a质粒的构建与鉴定结果
1.5%琼脂糖凝胶电泳得到5 388和1 275 bp片段(见图1)。测序结果中插入片段序列与PIAS-NY基因开放阅读框序列完全一致,因此原核表达载体PIASNY-pET30a已构建成功。
2.2 PIAS-NY原核表达情况
与诱导前菌液和对照组比较,诱导后大肠杆菌中均存在一分子量约为43 kD且表达量相当高的蛋白条带。与融合蛋白His-PIAS-NY 45 kD大小十分接近,而且仅存在2个IPTG诱导实验组中。因此提示PIAS-NY原核表达成功。见图2。
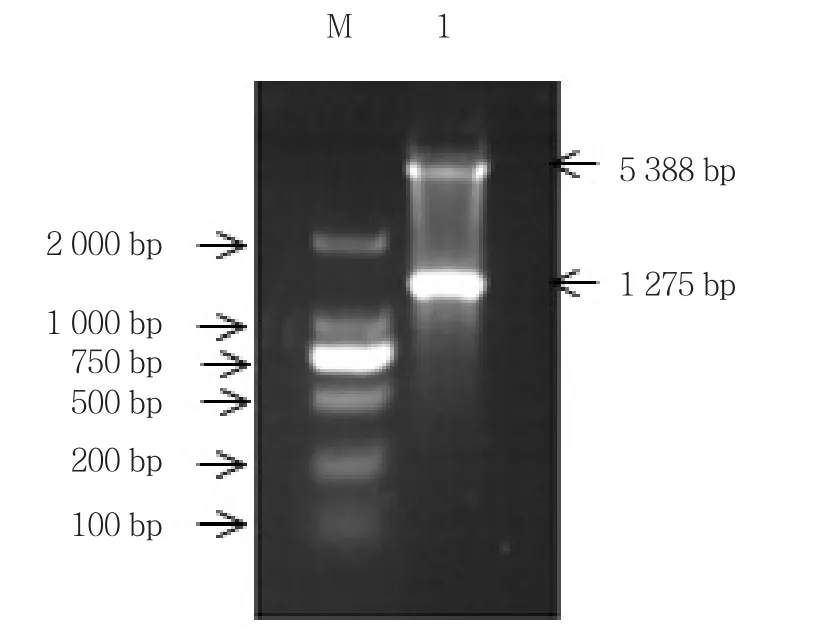
图1 PIAS-NY-pET30a重组质粒电泳图
2.3 His-PIAS-NY-His蛋白的纯化及复性
His-PIAS-NY-His融合蛋白主要存在于沉淀中,即以包涵体形式存在于大肠杆菌BL21中。100 mmol/L咪唑洗脱液和200 mmol/L咪唑洗液脱液中含高纯度蛋白。见图3。
2.4 Western blot检测结果
与对照组相比所得抗体效价均达到1∶100 000。PIAS-NY抗血清能够特异性识别GC2细胞和293T细胞中的PIAS-NY蛋白。见图4。
2.5 免疫共沉淀检测情况
293T细胞总蛋白中PIAS-NY抗血清沉淀出的复合物中存在RUVBL2蛋白,而且与293T细胞中的RUVBL2蛋白大小一致,然而BalB/C小鼠血清沉淀复合物中并未发现RUVBL2蛋白。结果显示制备的多克隆抗体在分子水平对蛋白具有特异识别作用。见图5。
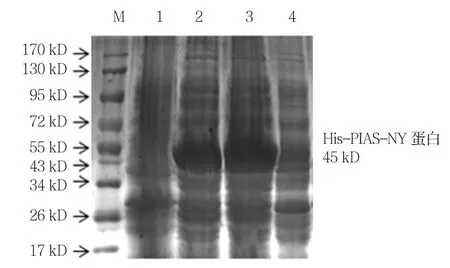
图2 诱导前后菌液SDS-PAGE电泳图
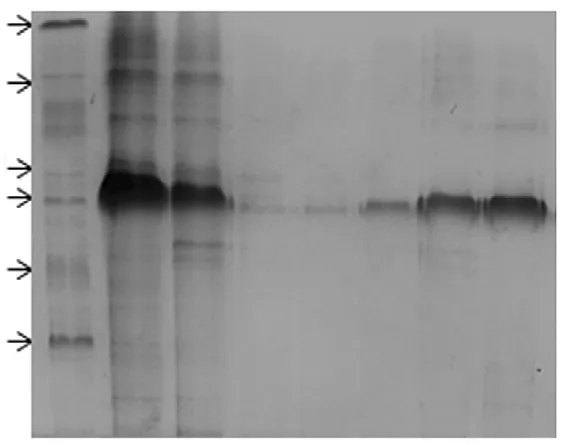
图3 His-PIAS-NY纯化SDS-PAGE电泳图
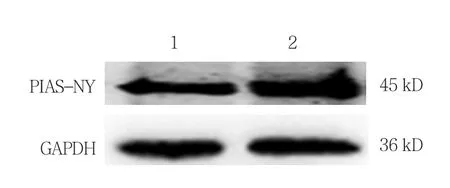
图4 PIAS-NY多克隆抗体特异性
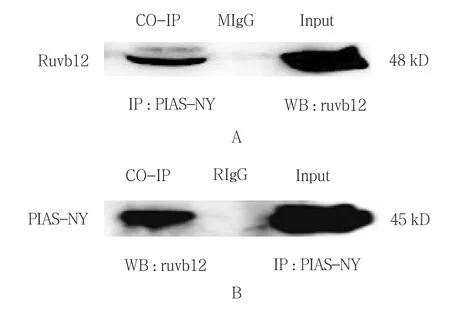
图5 PIAS-NY多克隆抗体特异性
3 讨论
抗体的制备技术经历很长时间的发展。包括多克隆抗体阶段、单克隆抗体阶段和基因工程抗体阶段[8]。目前实验过程中使用的抗体多为市场化的单克隆抗体,但对一些效果不太理想或者市场上没有的蛋白质抗体,实验室制备多克隆抗体满足一些实验需求。
PIAS蛋白家族最初是作为激活STAT转录活性的抑制蛋白被发现的[9]。最近研究发现,PIAS蛋白可以与60多种蛋白作用参与多种细胞活动[10],涉及到胚胎发育、造血肝干细胞自我更新[11]、先天性和后天适应性免疫反应[12]、空间学习和长期记忆等多个方面[13]。PIAS-NY基因是2004年新发现,相关研究报道非常少。之前的研究主要集中在细胞mRNA水平,而蛋白水平未见报道。因此,制备抗体是继续深入开展蛋白水平研究的基础。本实验用到的全部分子生物学技术是研究生阶段时郑英教授课题组实验室的固有技术,现均已成熟掌握。原核蛋白表达、提取、纯化和复性等一系列美国国立卫生研究院的实验原理和方法能深刻理解并能成熟运用,之前所在实验室也已成功制备出人NYD-SP28多克隆抗体[14]和Setd8多克隆抗体[15]。
基于前期的工作,笔者制备PIAS-NY多克隆抗体。诱导的融合蛋白中包括了蛋白PIAS-NY全部氨基酸序列,并且氨基端和羧基端均存在His标签,大大提高融合蛋白和Ni柱的亲和性,咪唑洗脱后获得高浓度可溶性融合蛋白。经过浓度梯度尿素稀释复性后,笔者获得具有一定空间构像的PIAS-NY免疫原。纯化蛋白经过充分乳化后,6次腹腔注射BalB/C小鼠。虽然BalB/C小鼠广泛用于免疫学、生理学的动物实验,但由于小鼠本身存在免疫缺陷,因此在免疫过程中仍应注意小鼠的健康状态。实验组共17只小鼠,最终只有13只小鼠存活,眼球取血。
ELISA检测所有存活小鼠的血清效价均在1∶100 000以上;Western blot也证实抗血清能够特异性识别293T细胞和GC2细胞的PIAS-NY蛋白;免疫共沉淀实验结果也说明抗血清能够识别并结合自然状态下PIAS-NY蛋白,同时也提示真核哺乳细胞中PIAS-NY与RUVBL2存在相互作用。而RUVBL2是进化上高度保守的AAA+家族的成员之一,在睾丸中高表达,具有ATP酶活性,与同源蛋白RUVBL1形成多聚物参与到不同的细胞活动中[16-17],如有丝分裂纺锤体的组装[18-20]、端粒酶的形成[21]、染色体重塑等[22-25],但是具体的分子机制并不清楚。通过质谱对睾丸RNA结合蛋白组学进行了分析,包括17种蛋白质,在圆形精细胞中表达量均比长形精子中高3倍,而RUVBL2是其中之一[26]。这也提示PIAS-NY和RUVBL2在精子发生中共同发挥重要作用。
综上所述,实验过程中已成功制备出PIAS-NY多克隆抗体,为下一步进行蛋白的功能和机制研究打下了一定的基础。
