荧光素酶互补法对谷子中SiPKS32/SiCaBP5的互作研究
2018-11-05潘教文李丹吴建国李臻王庆国管延安刘炜
潘教文,李丹,吴建国,李臻,王庆国,管延安,刘炜
(1.山东省农业科学院生物技术研究中心/山东省作物遗传改良与生理生态重点实验室,山东 济南 250100;2.济南大学医学与生命科学学院,山东 济南 250202;3.山东省农业科学院作物研究所,山东 济南 250100)
土壤中高浓度的Na+及Cl-易造成土壤盐渍化进而产生盐胁迫,是限制植物生长发育和农业生产的主要环境因子之一,也是全球面临的重大危机之一[1]。盐胁迫主要表现为离子胁迫、渗透胁迫及过氧化胁迫等造成的次生伤害,可通过破坏细胞的正常离子分布和动态平衡,干扰植物细胞内的各种代谢过程,造成细胞内活性氧的过量积累,引起细胞膜脂过氧化和蛋白质破坏,最终造成植物的伤害甚至死亡[2]。我国盐渍化土地主要分布在东北、华北、西北内陆地区以及长江以北沿海地带[3]。随着我国经济持续繁荣发展、人口数量逐渐增多,对粮食作物的需求量逐渐增大,粮食安全面临更加严峻的挑战。充分解析盐胁迫下植物的信号转导途径及相关机制,是进行植物抗盐分子育种的重要途径[2, 3]。
植物应对盐胁迫形成了一系列的适应机制,其中SOS(salt overly sensitive)信号途径是植物耐盐性的重要信号通路,该途径主要包括SOS1、SOS2、SOS3、SOS4、AKT1、NHX1和HKT1等基因[4, 5]。SOS1对于植物耐盐性非常重要,它是抗盐胁迫的关键基因。其编码细胞膜上的Na+/H+逆向转运体(Na+/H+antiporter),可以将盐胁迫下细胞质内的额外Na+排到胞外。SOS2编码丝氨酸/苏氨酸蛋白激酶,SOS3编码Ca2+结合蛋白[5]。SOS2的N末端有一个催化区域,C末端有一个调控区域,SOS3可与SOS2的调控区发生互作并激活SOS2形成复合体,SOS2-SOS3复合体通过磷酸化作用调控SOS1的表达,从而在维持细胞质钠钾离子的稳态平衡中发挥作用[2,6,7]。研究指出,SOS2和SOS3复合体的形成对于调控SOS1的活性具有重要作用,其在SOS途径中也具有重要地位。在拟南芥中过量表达AtSOS1显著增加转基因植株的耐盐性;将酵母质膜Na+/H+转运蛋白基因SOS2在拟南芥和水稻中过量表达也同样提高了转基因植株的耐盐性;同时在拟南芥中过表达AtSOS1、AtSOS2和AtSOS3,转基因植株的耐盐性比只表达其中一个基因有更加明显的提高,进一步证明SOS1依赖SOS2和SOS3的调节以介导Na+外排及其对植物耐盐性的重要作用[8]。
荧光素酶互补技术(firefly luciferase complementation imaging assay, LCI)是近年来形成的研究体内蛋白互作的技术。该技术是将荧光素酶(luciferase,LUC)分成N端部分(nLUC)和C端部分(cLUC), 它们不能自发重组发挥作用, 但当它们分别融合两个能够互作的蛋白时, 则能引发重组的荧光素酶活性。以含有不同LUC片段的植物表达载体,借助于烟草瞬时表达系统,在研究植物蛋白的相互作用中,可以实现活体细胞内实时、定量分析[9]。pCAMBIA1300-cLUC和pCAMBIA1300-nLUC载体图谱如图1。
本研究参照来源于不同物种的SOS2及SOS3同源基因设计特异引物,对谷子中SOS2和SOS3基因SiPKS32(XP_004978441)、SiCaBP5(XP_004969073)进行克隆,并进一步构建SiPKS32、SiCaBP5基因的cLUC、nLUC融合表达载体,以农杆菌介导共侵染烟草幼苗叶片,观察荧光信号情况,以期揭示SiPKS32与SiCaBP5的互作情况,探索谷子中是否存在SOS信号转导途径。对谷子中SOS基因家族作用机制的研究,将为进一步利用基因工程手段培育抗逆作物新品种奠定基础。
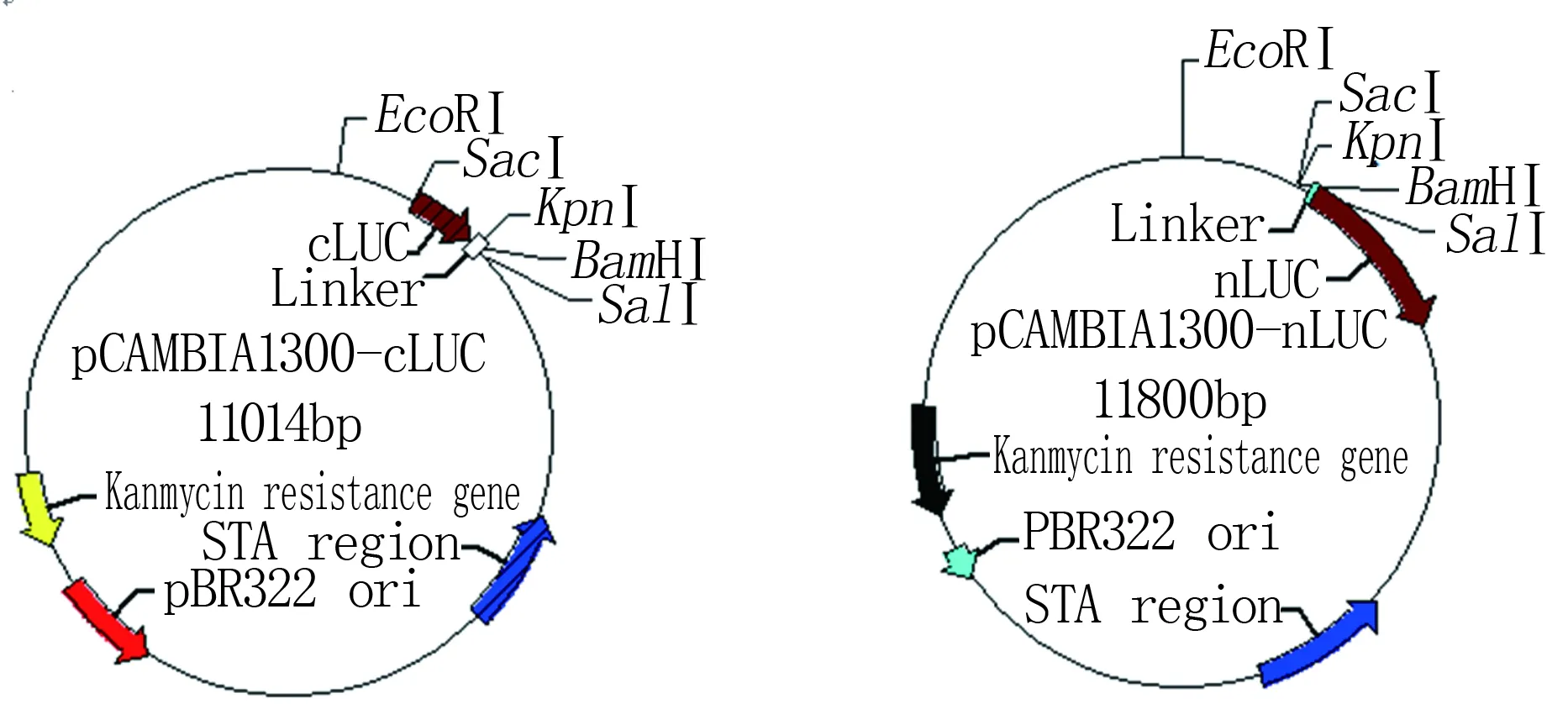
图1 pCAMBIA1300-cLUC、pCAMBIA1300-nLUC载体图谱
1 材料与方法
1.1 试验材料
谷子“豫谷1号”培养至三叶期备用。本氏烟(NicotianabenthamianaL.)播种于含有蛭石的营养土中,置于23℃下、光周期为8 h光照/16 h黑暗的人工气候室中,培养至叶片直径约2~3 cm时备用。
大肠杆菌(Escherichiacoil)TransDH5α、Blunt3克隆载体购于北京全式金生物技术有限公司;pCAMBIA1300-cLUC、pCAMBIA1300-nLUC植物表达载体由山东农业大学葛磊教授提供;农杆菌GV3101由本实验室保存。引物由青岛擎科梓熙生物技术有限公司合成;测序由山东省农业科学院生物技术研究中心完成。
1.2 SiPKS32、SiCaBP5基因的克隆
以谷子幼苗为材料,参照TransZol Plant试剂盒说明书提取总RNA,并参照PrimeScriptⅡ1st Strand cDNA Synthesis Kit试剂盒说明书对所提RNA进行反转录,得到单链cDNA。根据不同物种中SOS2、SOS3基因同源序列信息及KpnⅠ、SalⅠ酶切位点,设计SiCaBP5特异性引物,SiCaBP5-S:AGGTACCATGGGGTGCATACCAACAAAG(下划线为KpnⅠ 酶切位点),SiCaBP5-A:AGTCGACCATTTCAGGTTTGGCAGC(下划线为SalⅠ酶切位点)。SiPKS32特异性引物,SiPKS32-S: AGGTACCATGAGTACAACCAAGGTGAAGA(下划线为KpnⅠ酶切位点),SiPKS32-A:AGTCGACCTAAGCAGCAGGTTGCATCG(下划线为SalⅠ酶切位点)。经94℃下5 min;94℃ 30 s,57℃ 30 s,72℃ 45 s,35个循环;72℃延伸10 min, 获得谷子中的SOS2同源基因序列SiPKS32(XP_004978441),SOS3同源基因序列SiCaBP5(XP_004969073),将基因片段连接到克隆载体后转化大肠杆菌DH5α,送公司测序。
1.3 含有N端及C端LUC片段的SiPKS32、SiCaBP5互补表达载体的构建
将克隆测序正确的SiPKS32、SiCaBP5基因序列与pCAMBIA1300-cLUC、pCAMBIA1300-nLUC载体分别进行KpnⅠ、SalⅠ双酶切,酶切片段经T4连接酶连接,转化大肠杆菌DH5a,进行阳性重组质粒pCAMBIA1300-SiPKS32-cLUC、pCAMBIA1300-SiCaBP5-nLUC的鉴定。将鉴定为阳性的重组质粒进一步转化农杆菌GV3101。
1.4 荧光素酶互补试验
选取叶色浓绿、生长旺盛的本氏烟叶片用作烟草瞬时表达系统的材料。将含有不同重组质粒的农杆菌GV3101培养至稳定期,5 000 r/min离心10~15 min,先用3 mL左右的悬浮液悬浮,测OD600值,然后用悬浮液把OD600统一调至1,静置3 h。将烟草叶片分为四个区域,分别注射不同的菌悬液组合,试验组:pCAMBIA1300-SiPKS32-cLUC和pCAMBIA1300-SiCaBP5-nLUC;对照组1:pCAMBIA1300-SiPKS32-cLUC和pCAMBIA1300-nLUC;对照组2:pCAMBIA1300-SiCaBP5-nLUC和pCAMBIA1300-cLUC;阴性对照组:pCAMBIA1300-nLUC和pCAMBIA1300-cLUC。
注射后的烟草叶片在光照培养箱中继续培养2 d,之后在相同的注射部位注射荧光素酶底物D-荧光素钾盐溶液,注射面积需大于菌液注射面积,于黑暗中静置5 min,将叶片取下,背面向上,置于活体成像系统中,利用CCD相机检测荧光信号,并拍照记录。
2 结果与分析
2.1 SiPKS32、SiCaBP5基因序列分析
利用拟南芥SOS2(NM_122932)和SOS3(NM_001203448)蛋白序列在NCBI中进行BLAST检索分析,分别找到谷子中同源基因SiPKS32(XP_004978441)和SiCaBP5(XP_004969073),通过设计特异引物序列进行PCR克隆后测序分析。序列比对分析发现,SiPKS32与AtSOS2的相似性为56%,在N端含有保守的激酶区(图2),而且含有保守的SOS3结合区序列FISL motif。SiCaBP5与AtSOS3的相似性为51.88%,含有3个保守的EF hand Ca2+结合位点(图3)。
2.2 SiPKS32-cLUC、SiCaBP5-nLUC互补表达载体的构建
将测序正确的SiPSK32和SiCaBP5序列经KpnⅠ、SalⅠ双酶切,分别与cLUC、nLUC载体片段连接,获得重组质粒pCAMBIA1300-SiPKS32-cLUC、pCAMBIA1300-SiCaBP5-nLUC,利用双酶切对质粒进行鉴定, 鉴定结果如图4,酶切后的片段大小与SiPKS32和SiCaBP5基因大小一致,条带清晰,重组质粒构建成功。
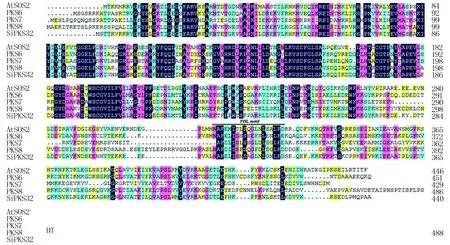
AtSOS2(NM_122932);PKS6(AF339147);PKS7(AF339148);PKS8(AF339149);SiPKS32(XP_004978441)。图中划线处为FISL motif区。

AtSOS3(NM_001203448);SCaBP5(NP_567533.1);SCaBP6(NP_194387.1);SiCaBP5(XP_004969073)。图中划线处为EF hand Ca2+结合位点。

M:DNA Maker,由上到下依次为5000、3000、2000、1000、750、500、200 bp和100 bp;1、2为pCAMBIA1300-SiPKS32-cLUC的酶切片段;3、4为pCAMBIA1300-SiCaBP5-nLUC的酶切片段。
2.3 荧光素酶互补法对SiPKS32/SiCaBP5互作检测
将上述鉴定为阳性的SiPKS32-cLUC、SiCaBP5-nLUC重组质粒进一步重转化农杆菌GV3101中。应用LCI方法,对SiPKS32和SiCaBP5在体内的互作情况进行检测。结果(图5)显示,含有SiPKS32-cLUC、SiCaBP5-nLUC重组质粒的菌液共转化烟草叶片后,可检测到荧光信号,而阴性及对照组荧光信号较弱或无信号,说明这两个蛋白在植物体内可以发生互作。
3 讨论与结论
土壤盐渍化已经成为全球面临的重大危机之一,它可以导致土地荒漠化、耕地退化等问题,给农作物生产带来了严重的负面影响。近年来,研究人员通过克隆基因和遗传转化等方法找到了参与植物抗盐胁迫的一些重要途径及相关基因,并构建了一些调控机制模型,例如活性氧清除、离子区域化、渗透调节等。其中盐胁迫信号的感知及传递是植物响应盐害及抗盐的重要过程。SOS信号通路即为该过程中的关键组分。拟南芥中SOS信号通路的研究相对深入,近年来,根据公共数据库(如NCBI、TAIR、NASC)中大量的拟南芥表达谱芯片数据,通过对盐胁迫下SOS途径响应基因表达和蛋白质互作信息的整合, 已构建了SOS途径特异表达调控网络[4]。在水稻[10]、油菜[11]等研究中,也有关于SOS信号通路的报道。谷子近年来作为新兴禾本科植物的模式作物,其突出的抗旱耐瘠特性,已越来越受研究者的关注。谷子如何参与逆境响应,抗逆过程中是否也存在SOS信号途径,是否也存在通路组分SOS2、SOS3,它们之间是否有互作等问题也亟待解决。
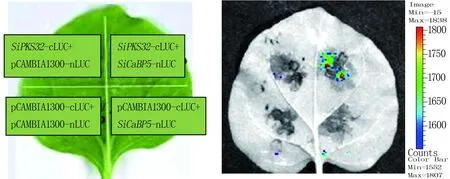
左侧为不同区域菌液组合示意图,右侧为荧光图。
参照拟南芥中的研究进展,在谷子基因组中检索到部分SOS2、SOS3类基因的同源序列。经同源克隆进一步对PCR产物进行测序及序列分析,结果显示谷子中获得的SiPKS32、SiCaBP5基因分别属于SOS2、SOS3类基因,含有SOS2和SOS3保守的结构域。初步显示谷子中存在SOS信号通路。已知SOS2和SOS3复合体的形成,可进一步激活SOS1的活性,从而使SOS通路参与植物抗逆过程[12]。为了进一步检验谷子中SiPKS32、SiCaBP5是否也存在互作进而形成复合物,以便深入解析基因功能,本研究应用LCI法对其进行了检测。LCI法不受植物自发荧光的干扰,且操作简单,不需要裂解细胞提取蛋白质,可以很好地反映植物生理条件下蛋白的互作状态。当含有SiPKS32基因的cLUC与含有SiCaBP5基因的nLUC由于基因的互作而结合后,就会表达出完整的荧光素酶,作用于荧光素酶底物从而检测到荧光信号。本研究结果显示含有SiPKS32与SiCaBP5的菌液组合共侵染的烟草叶片可检测到荧光信号,而仅含有空载体的组合或一个带有基因的载体与一个空载体的组合均不能使侵染后的叶片产生荧光信号,说明蛋白间存在互作。SOS信号途径中,除了SOS2、SOS3,还包含SOS1、SOS4、AKT1、NHX1和HKT1等组分[5, 13]。目前也已在谷子中分离并鉴定了SOS1类基因,对其余各组分的鉴定及功能分析工作也正在开展,将为全面验证及解析谷子中SOS通路及其作用机制、培育谷子抗逆新品种提供依据。
