水生蔬菜答农民问(14):子莲为什么会出现死花死蕾和空壳莲籽?
2018-09-22刘义满魏玉翔
刘义满 魏玉翔
经常有从事子莲(Nelumbo nucifera Gaertn.)种植的农民朋友咨询:“子莲为什么会出现死花死蕾和空壳莲籽?”实际上,这个问题在子莲的整个开花结籽季节都存在。要回答这个问题,需要先了解莲花芽分化时期与始花节位的关系,了解影响莲开花结籽的主要因素,才能更好地理解导致子莲死花死蕾和空壳莲籽比率较高的主要原因及其克服措施。因为子莲与花莲花芽分化和开花结籽习性类似,可以相互借鉴参考,为更好地说明问题,本文也利用了部分花莲观测研究资料。
1 莲花芽分化时期与始花节位
子莲是以采收种子为主的一类莲品种。在植物分类学上,子莲、藕莲及花莲均属于一个“种”。子莲品种花朵数一般达4 000~6 000朵/667 m2;藕莲的膨大根状茎(藕)粗壮(藕莲品种直径不小于4 cm)、产量高,但花朵数较少,每667 m2花朵数一般数十朵到数百朵,有的品种甚至不开花;花莲以观赏为主,花朵数多,品种间花色、花器官(花萼、花瓣、雄蕊、雌蕊等)形状和数量、结籽性、株型及膨大根状茎变异极大。多数花莲品种结籽量较少或不结籽,膨大根状茎(藕)也很小。
曾经有人解剖越冬种藕的顶芽结构,发现其已具备苗的基本结构,一般含有4个幼叶及未伸展的节间(即4个节及节间),以及顶芽生长点。顶芽为混合芽,含有营养芽的基本部分,每个芽内幼叶基部叶背处,都有一个花原基。而且,顶芽内各节上花及叶原基的发育程度,从近藕体(种藕)处向前端方向,有相应递减的现象。春季顶芽萌发后,一般从第4~5 节开始形成当年新的花原基[1,2]。
花芽分化是开花结籽的前提,总体而言,子莲和花莲的花芽分化能力强于藕莲。子莲和花莲的花芽分化规律更为相近,实际观察到:①有的花莲品种越冬前即进行花芽分化,并形成单独的花蕾(纯花芽,图1A),翌春单独萌发(图1B),不过,现有子莲品种在武汉地区种植时未曾见到过这种情况,但在少数花莲品种中存在;②有的品种,顶芽萌发伸长1个节间时,可见花芽萌发(图1C、1D);③有的品种,顶芽萌发伸长2个节间时,可见花芽萌发(图1E);④太空36号子莲田间,可以见到一些植株在顶芽萌发伸长2~3个节间时,花芽萌发 (图1F、1G)。也就是说,子莲早期的花芽,可能在春季顶芽萌发前即已经分化;后期的花蕾,则应为当年春季萌发后分化形成的。

图1 花芽分化时期始花节位
2 莲主要开花结籽习性
2.1 诱导莲花芽分化开花需要长日照
根据植物生理学知识分析,诱导莲植物花芽分化与开花的主要因素应为长日照(即诱导开花的日照长度应长于一定时数)[3]。但是,不同生态型[4]莲品种或资源之间,临界日长(即诱导开花所需的最短日照时数)可能会有差异。由于缺乏系统精准的研究,这里暂且根据相关地区日照和气温数据资料(表1)及不同生态型莲品种或资源的现蕾开花时期,推测不同生态型莲代表分布地区诱导芽分化和开花的实际日照长度。
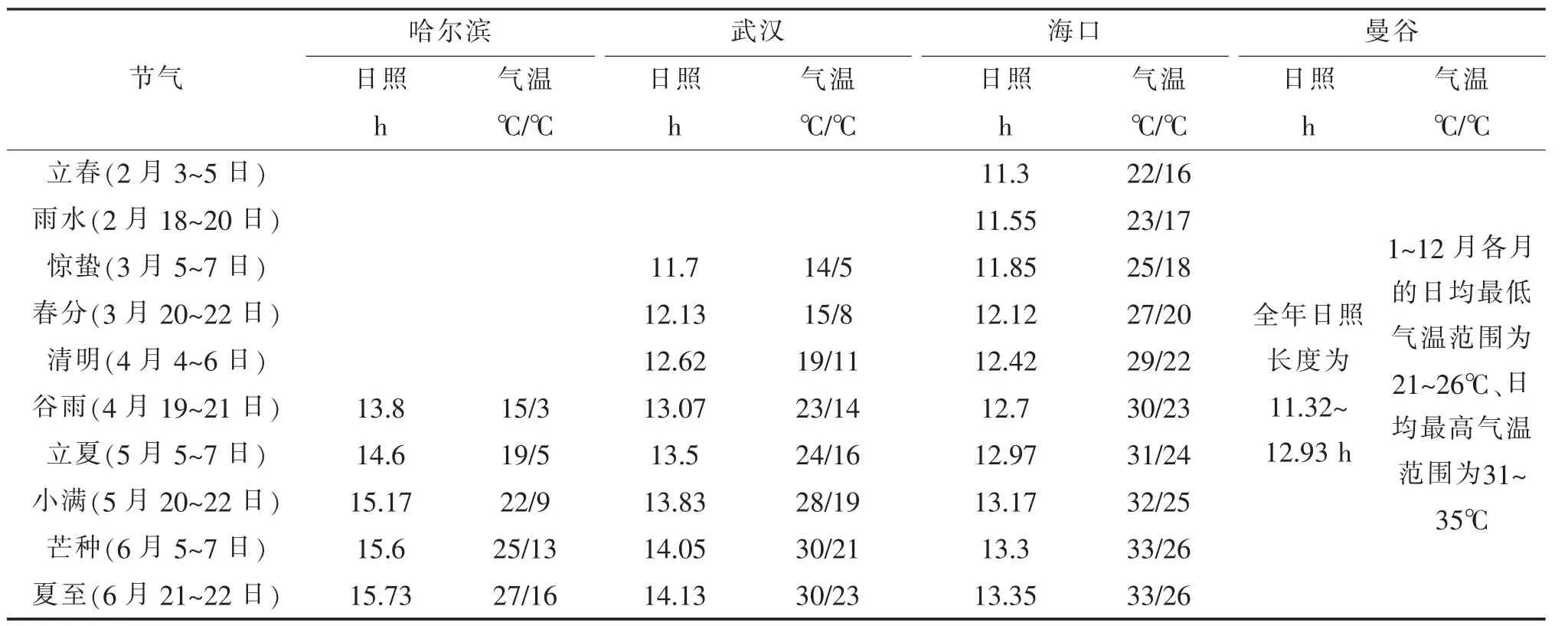
表1 不同生态型莲代表分布地区日照和气温数据
①黑龙江哈尔滨 温带地区,全年日照长度为8.50~15.73 h,其中,当地野莲(温带生态型莲)始花期约为7月上旬,从莲春季萌发至进入现蕾开花季节的日照长度变化趋势为13.80~15.73 h。黑龙江野生红莲南移到部分地区的表现如下。a.辽宁沈阳:开花、结藕。7月上旬始花,沈阳地区露地生长的莲,一般4月中旬开始萌发,比哈尔滨地区约早2周,6月1日日照长度为15.02 h,全年最长日照长度为15.23 h(6月 21日)。b.北京:不开花,能结藕。北京地区的最长日照长度为15.02 h。c.山东潍坊:6月上旬播种,实生苗能结藕,但不开花。d.江苏宿迁:不开花。e.湖北武汉:5月结藕,全年不开花。综合分析,推测哈尔滨当地日照长度达15.02~15.23 h即可诱导当地温带生态型莲(黑龙江野生红莲)花芽分化和开花。

图2 热带生态型莲开花结籽情况(新加坡)
②湖北武汉 亚热带地区,全年日照长度为10.17~14.13 h,其中,源自湖北武汉地区的亚热带生态型莲5月下旬开始现蕾,从莲春季萌发至进入现蕾开花季节的日照长度变化趋势为11.7~14.13 h,推测武汉地区日照长度为12~13 h时即可诱导当地亚热带生态型莲花芽分化和开花。
③ 新 加 坡 (地 理 维 度 1°17′22″N,经 度103°51′00″E) 热带地区,全年气温均适合种植莲,且日照长度变化很小。如2006年最短为12.05 h,最长为12.20 h,全年日照长度变化幅度不到9 min(0.15 h)。可较准确地推测,新加坡当地诱导当地热带生态型莲花芽分化和开花 (图2)的日照长度为12.05~12.20 h。
另外,根据相关资料推测,同为热带地区的泰国曼谷和我国海南海口地区,日照长度为11.5~12.0 h时即可诱导当地的热带生态型莲花芽分化和开花。
在北半球,莲春季萌发至进入开花季节期间,由北向南日照长度是逐渐缩短的,因此,温带生态型莲引种到亚热带或热带地区,或者亚热带生态型莲引种到热带地区,可能由于日照时数不够,导致不能诱导花芽分化,进而导致不能开花结籽。前述的黑龙江野生红莲南移引种种植实践证明了这一点。其他实例如:部分源自中国的莲品种引种到泰国曼谷种植,不开花,但人工补充光照后能够开花结籽;有泰国朋友将美洲黄莲(N.lutea,与中国莲类似)引种到泰国曼谷种植,亦多年不见开花。相反,热带生态型莲或亚热带生态型莲,向北引种时,长日照条件容易满足,只要温度条件适宜,则开花性能能够更好地表现。国家种质武汉水生蔬菜资源圃曾经将泰国、新加坡、印度加尔各答等地的莲,向北引种到湖北武汉种植,均能正常开花,且群体花期较长,但结藕较差;武汉地区的荷花品种,北移到东北地区种植,开花相对较早,结藕相对较迟。综合分析,总体上可以说,莲植物北移栽培时开花期相对早,结藕期相对迟或不结藕;南移栽培时开花期相对迟或不开花,结藕期相对早[4]。
2.2 莲具有自我调控花内温度的特性
莲具有自我调控花内温度的特性,环境气温在10~45℃范围变化时,能将花朵内温度调控到30~36℃,有助于受精[3]。但是,已有报道中的试验材料比较单一,至于自我调控花内温度的能力在莲不同品种类型之间是否一致,则需要进一步研究。如果不同莲品种之间自我调控花内温度的能力有区别,则开花结籽率相对较高的子莲品种,其自我调控花内温度的能力应该相对较强。
2.3 莲兼具自花授粉和异花授粉能力,但异花授粉结实率一般较高
论及子莲授粉受精习性,有必要弄清楚“自花授粉”“异花授粉”和“自然授粉”的含义和区别。
①自花授粉(Self-pollination,自交) 生产上,子莲采用无性繁殖,自花授粉不仅仅指雌蕊接受同一花朵的花粉,而且同一品种内不同花朵、不同植株之间的相互授粉,也是自花授粉。
②异花授粉(Cross-pollination,异交) 于子莲而言,不同品种之间的授粉称为异花授粉。
③自然授粉(Oopen-pollination) 也叫自由授粉或开放授粉,指主要依靠昆虫、鸟、风或其他自然机制进行的传粉。
对于子莲而言,如果规模化种植单一品种,则整个种植区内植株之间的自然授粉,本质上还是自花授粉;但如果是相邻种植了不同品种,则自然授粉时,可能有自花授粉和异花授粉同时发生。
从莲的开花习性看,莲花开放首日,雌蕊柱头已经具备黏液,具备接受花粉的能力。不过,首日开放的花中,雄蕊花药尚未成熟开裂,无花粉散出,因而其雌蕊柱头接受不到自身花朵产生的花粉,但可以接受外来花粉。有人曾经用6个花莲品种试验,结果表明,①自交结实率平均为8.70%,自然授粉结实率平均为27.27%,二者差异明显;②品种间杂交结实率差异明显,低者为0,高者达38.77%;③母本品种相同时,父本品种不同,结实率差异大;父本品种相同时,母本品种不同,结实率差异亦大;④有的杂交组合,正反交结实率差异大[6]。有人调查,子莲品种鄂城红莲自交结实率达到47.68%,而以其为母本的9个杂交组合平均结实率达67.72%[7]。莲以异花授粉为主,自花授粉为辅,自然授粉条件下异花授粉机会比自花授粉多[8]。
中国和泰国学者进行的联合研究发现,野生红花莲Rachine和Nakhon Sawan(在泰国,皆被农民用来采集莲籽),二者具有自交不亲和性,其自花授粉结籽率分别为2.3%和2.8%,自交不亲和指数[Index of self-incompatibility,自交不亲和指数=(异花授粉单花结籽粒数-自花授粉单花结籽粒数)/异花授粉单花结籽粒数×100%]则分别高达95.5%和96.0%。同时,Rachine作母本与Nakhon Sawan作父本进行的杂交结实率为69.2%;Nakhon Sawan为母本与Rachine为父本进行的杂交结实率为50.8%;自然授粉时,Rachine和Nakhon Sawan的结实率分别为43.4%和61.6%。这一现象似乎表明,部分莲品种单独种植时,种子产量较低或不能产生种子。选择配置适宜的品种相邻种植,让不同品种之间相互自然授粉,可以提高子莲结实率[9]。
2.4 植株营养与生理状况
子莲进行正常的花芽分化与开花结籽,是以植株正常的生理功能发挥为基础的,如出现土壤养分供给能力下降、根系养分吸收能力下降或因病虫为害而死亡、叶片受到伤害或死亡、发育中的莲籽直接受到害虫为害、植株营养缺乏、生理代谢功能被破坏等现象,都可能导致花芽分化和开花结籽进程异常,进而出现死花死蕾和空壳率增加的现象。

图3 死花死蕾和空壳莲籽
3 子莲会出现死花死蕾和空壳莲籽的主要原因及其克服措施
死花死蕾和空壳莲籽(图3)在子莲产区非常常见,直接结果是减产。
3.1 品种习性
本文前述内容已经谈过,诱导子莲花芽分化开花需要长日照,而且不同生态型之间的临界日长可能有所不同。目前生产上主要应用的子莲品种不多,南北引种时,品种间的适应性差异尚未引起重视。但是,随着子莲新品种选育工作的进一步开展,品种将越来越丰富。品种南移栽培时,应该考虑品种对日照长度的适应性,尤其是将偏北地区(如武汉)选育品种向偏南地区(如海南)引种时应予以重视。另外,如果子莲品种之间调控花内温度的能力存在差异,则子莲新品种选育和引进利用时,也需要考虑该因素。
建议措施:在规模化种植子莲新品种前,应先行小规模引进多个品种试种不少于2年,之后再根据不同品种表现,筛选确定规模化种植的品种。特别是维度跨度较大地区之间引种时,应做到先试种筛选,再扩大规模。
3.2 早期花芽分化质量
子莲“早期花芽”指春季萌发前种藕芽内已经分化形成的花芽。“早期花芽”分化与发育质量,应该与越冬前种藕形成时的养分供应、营养物质贮藏积累有关,也与春季气温影响等关系较大。
建议措施:越冬前,在荷叶尚绿时,加强田间管理,适当追肥,促进种藕发育、花芽分化及养分积累。春季,在种藕采挖、转运、定植及生长前期,主要防止低温冷害,请参阅《水生蔬菜答农民问(5):莲藕定植后,为什么迟迟不“发棵”?》一文中的相关内容[10]。
3.3 除草剂为害
常见现象是邻近水稻田的子莲田,在水稻田使用除草剂时,除草剂随风飘、随水流而进入子莲田,导致为害。也有子莲田种植者在田埂或田间使用除草剂不当,导致除草剂为害。除草剂为害子莲时,妨碍了植株自身的生理代谢,重者整片植株死亡,轻者叶片发黄、花药白化、花粉失活、授粉受精不良、心皮泡状凸起、空壳秕籽增加、结实率降低或不结籽、根状茎节间不伸长等[11,12]。如草甘膦(农达)主要抑制植物体内的烯醇丙酮基莽草素磷酸合成酶,从而抑制莽草素向苯丙氨酸、酪氨酸及色氨酸的转化,使蛋白质合成受到干扰,为害严重时常导致植物死亡。
建议措施:生长季节防止除草剂进入子莲田,发生为害时及时换水,并追肥增强植株长势。有关内容请阅读《几种除草剂为害莲的症状》[11]和《水生蔬菜答农民问(6):莲藕叶片为什么发黄了?》[12]等文章。另外,部分用于病虫害防治的农药也可能对子莲植株产生药害,需要防止。
3.4 不合理整地施工导致土壤耕作层的破坏
部分地区在子莲栽培用土地整理过程中,采取了不合理的土地平整方法。主要表现为将原有地块上的耕作层土壤推移他处,直接对耕作层以下的土层进行耕翻平整,其后对耕作层土壤未予回填。这种情况有的是局部发生(同一田块“挖高填低”),有的是连片发生(实施土地整理项目,大面积连片平整)。土壤耕作层的破坏,主要导致土壤结构不良,通透性降低,养分供给能力差,不利于根系正常生长或功能发挥,进而导致植株生长不良或死亡。通常见到的现象是,根系表现正常,但不能正常吸收养分,植株出现“饥饿”症状,叶片发黄、枯萎直至死亡。这种情况出现时,花芽分化和花蕾发育自然也不能正常进行。关于这方面的问题,在《水生蔬菜答农民问(6):莲藕叶片为什么发黄了?》[12]一文中进行了较为详细介绍,建议参阅。
3.5 病虫为害
病虫为害导致子莲死花死蕾和空壳莲籽的现象比较常见。常见病虫害包括为害浮叶的莲潜叶摇蚊,为害立叶的叶斑病、莲叶脐黑腐病、莲缢管蚜、斜纹夜蛾、中喙丽金龟,为害根状茎和根系的莲藕腐败病、食根金花虫,南方地区还有为害叶的福寿螺、茶黄蓟马。还有直接为害莲籽的蛀螟和直接危害叶片的梨剑纹夜蛾等(图4)。病虫为害严重时,花芽分化和开花结籽也自然受到抑制。有关病虫为害及其防治,请阅读《水生蔬菜答农民问(4):莲藕有哪些主要病虫害?如何防治?》[13]。
3.6 异花授粉受精率
众多研究证明,“莲兼具自花授粉和异花授粉能力,但异花授粉结实率一般较高。”[8]目前,子莲种植中,同一区块内通常采用单一品种,并强调保持单一品种的品种纯度,客观上是以自花授粉为主。如何提高异花授粉受精率、减少死花死蕾和空壳莲籽,是一个值得思考的问题。
建议措施:①不同子莲品种混杂种植。不同子莲品种混杂种植,之前是不提倡的,但理论上应该是可行的。如果选择品质优、产量高、长势相似、花期一致、相互授粉后受精结实率高的品种,混杂种植,无疑提高了异花授粉率。事实上,现有通心白莲产区、枯莲籽(壳莲)脱壳加工产区在收购原料时,来自不同品种的原料实际上也是混收的,并没有严格的品种区分。如果品种选择配置合理,对产品商品性没有影响。

图4 蛀螟和梨剑纹夜蛾为害子莲

图5 子莲产区放养蜜蜂
②保护和放养传粉昆虫。目前,提倡子莲田放养蜜蜂(图5)。子莲田放养蜜蜂传粉,一般可以提高结籽率、降低死花率及增加产量,效果明显[14~16]。通常,可以提高产量10%以上,高者增产近30%。③人工辅助授粉。人工收集花粉,进行人工辅助授粉,也可以大大提高子莲授粉受精率,减少死花死蕾和空壳莲籽,提高结籽率和产量。
3.7 开花结籽季节的连阴雨天
子莲开花结籽期若出现持续2天及以上的连阴雨天气,则随着降雨天数的增加,对子莲授粉受精的妨碍越大,致死花死蕾率增加,开花花结籽降
低[17]。
建议措施:持续的连阴雨天对子莲授粉受精的妨碍,需要进一步研究有效的应对措施。
3.8 施肥技术
施肥技术不当,直接影响植株养分供应。目前,产区种植户在子莲栽培时,有的不施基肥,而以1~2次追肥为主;有的只是进行一次性施肥;有的采取“基肥+1~2次追肥”的方式施肥。这些施肥方式,在土壤养分本底值较高的田块(如多年的水产养殖塘)是可行的,但若采用水稻田等浅水田栽培子莲,则有进一步改进的必要。与藕莲种植不同,子莲最大的特点有2条:其一,主要产品器官为种子,系有性器官;其二,子莲开花结籽是一个持续的过程,莲籽成熟采收也是一个持续的过程,开花结籽与莲籽成熟过程在时期上存在交叠。
建议措施:将施肥方式改为“基肥+1~2次重追肥+多次轻追肥”,同时注意补充硼肥等微肥。譬如,以667 m2面积为单位,中等肥力田块,“基肥”可以施 N∶P2O5∶K2O 为 15∶15∶15 的复合 肥 50 kg、硼 砂1.0 kg(或硼酸 0.5 kg)及七水硫酸锌 1.0~1.5 kg(或一水硫酸锌 0.5~1.0 kg);“1~2 次重追肥”宜于定植后30~35天和第60~65天分别施第一次、第二次追肥,每次施 N∶P2O5∶K2O 为 15∶15∶15 的复合肥和尿素各10 kg;“多次轻追肥”即“少量多次追肥”,指进入采收期后,每15天追肥1次,每次施尿素和硫酸钾各3~5 kg。追肥时,应避免肥料溅落或滞留于叶片上。另外,进入开花期后,宜用0.1%~0.2%硼砂、硼酸或聚硼酸钠水溶液进行叶面喷施,喷用量60 kg,每10~15天一次[18]。目前,各级行政和技术部门均在倡导有机肥替代化学肥料,减少化学肥料施用,表2中根据几种市售商品有机肥的检测报告(均符合NY 525-2012《有机肥料》规定)[19],列出了子莲的推荐施用量和方法,供参考。
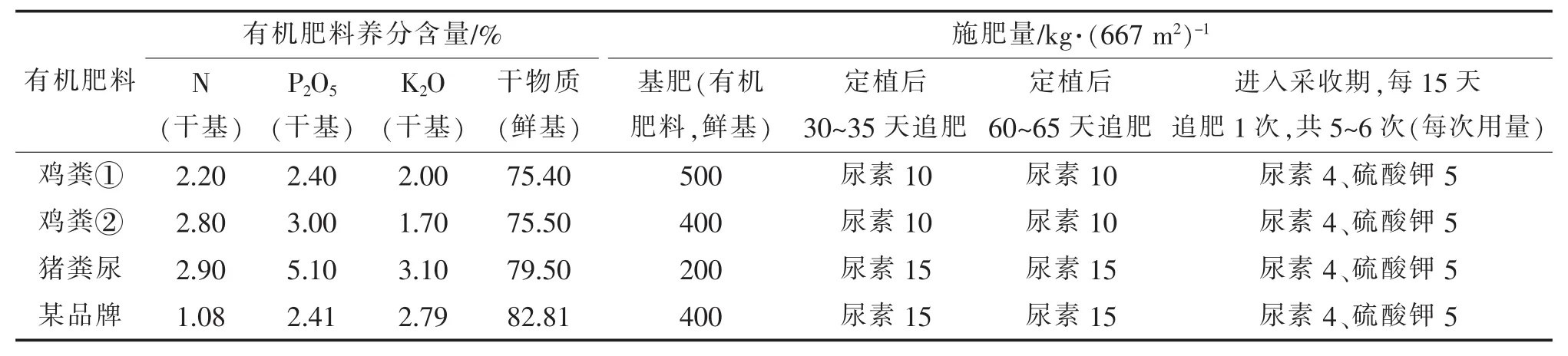
表2 标准化有机肥料的籽莲田推荐施用量及配合施肥方案
3.9 植株调整
目前,在湖北、湖南等子莲主产区较为普遍采用的栽培模式是“一次定植,连续3~4年栽培”。这种模式下,虽然节省了每年耕翻整地和大田定植的人工,但也存在一些问题。主要问题是从定植翌年开始,田间植株密度往往过大。一般第1年大田定植用种量为120~150支/667 m2,但当年栽培季节结束时,田间藕支数就达到800~1 500支/667 m2,这些藕支也就成立翌年的种藕。显然,植株密度是偏大的,容易出现养分供给过于分散,株间过于密闭,需要疏苗[17]。
建议措施:从定植后的第2年开始:①湖北地区宜在6月中下旬以前,高强度采收藕带销售,一方面疏除了部分植株,另一方面采收的副产品藕带也是增收的重要渠道;或②待春季田间植株萌发一段时期后(武汉地区为4月中旬),用机耕船将全田旋耕一遍,以重新萌发出的植株进行栽培管理;或③6月中下旬前,间隔割除部分植株,可以每5 m宽的田间,割除3 m,保留2 m留田栽培。此外,在年度内栽培管理过程中,可以结合田间管理和莲蓬采摘,及时摘除老叶、病叶、过密叶、瘦弱叶,增加田间通风透光性。
3.10 其他
湖北洪湖市部分种植户经验,进入10月后,气温渐低,为了延缓秋季低温对子莲开花结籽的不良影响,延长开花结籽期,采取逐步加深种植塘水深的做法,也有一定效果。加深水深的做法一般在鱼塘改成的子莲田进行,水深可加深至1.2 m左右。因为水体保温性好,秋季水温下降速度比气温迟,本质上起到了与设施覆盖保温类似的效果,至11月上中旬仍有部分莲蓬可以采收鲜食。这也可以视为子莲延长栽培季节的一种有效措施。
