谷子FT/TFL1家族的全基因组鉴定与分析
2018-09-12赵庆英杨致荣
赵庆英,杨致荣
(山西农业大学文理学院,山西 太谷 030801)
谷子(Setaria italica)是传统的优势作物,具有耐瘠薄、耐干旱、耐储藏、营养丰富、粮草兼收等特点[1]。针对日益严重的水资源短缺问题,谷子作为重要的战略储备作物,对保障干旱、贫瘠地区的粮食安全起着非常重要的作用[2]。随着人们消费水平的提高以及对食品营养与安全的重视,人们对小米的需求也逐年增加,但是小米的产量低,还不能满足现代化产业的需求。因此,提高谷子产量是谷子研究的主要方向之一。
开花是植物从营养生长到生殖生长转变的开始,是植物生长发育的重要阶段,植物在适宜的条件和时间开花,能够确保其生物量和繁殖率[3]。植物开花受到几种开花途径的共同调节,如光周期途径、春化途径、内源激素和自主途径等[4],FT和TFL1基因是这些开花途径下游的关键基因,编码磷脂酰乙醇胺结合蛋白(PEBP)[5],在植物从营养生长向生殖生长转变的过程中起着重要的作用[6]。在拟南芥中,FT/TFL1家族有6个成员,分别为FLOWERING LOCUS T (FT),TERMINAL FLOWER 1(TFL1),TWIN SISTER OF FT (TSF),BROTHER OF FT AND TFL1 (BFT),ARABIDOPSIS THALIANA CENTRORADIALIS HOMOLOG(ATC)和 MOTHER OF FT AND TFL1(MFT),可以分为 3 个亚类,分别为FT亚类(包括FT和TSF),MFT亚类和TFL1亚类(包括TFL1,BFT和ACT基因)[6-7]。
FT是一个开花整合基因,也是诱导开花的关键基因,它能促进植物从营养生长到生殖生长的转变[7-8]。在长日照条件下,FT基因能在长日照植物(拟南芥等)的叶片维管组织伴胞中被CONSTANS(CO)基因诱导激活表达[9],FT蛋白与一种新型的内质网膜蛋白 FT-INTERACTINGPROTEIN1(FTIP1)结合后被转运到茎顶端分生组织,与碱性亮氨酸拉链结构域(basic Leucine Zipper,bZIP)转录因子 FD结合,直接激活开花基因SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1(SOC1)和 APETALA 1(AP1)基因,控制植物的成花转变[10-11]。
在拟南芥中,FT基因过量表达导致开花时间提前[12];在水稻中,Hd3a是FT的同源基因,在短日照条件下促进开花[11];将日中性植物番茄中的FT同源基因SlSP3D转到拟南芥中,能使拟南芥开花时间提前,说明SlSP3D是一个开花促进基因[11]。转录因子FLC和光敏色素B1可以抑制FT基因的表达[13]。
在拟南芥中,FT的同源基因TFL1,参与维持茎尖分生组织的状态、延迟其向花器官的转变[14],并与花序的形成有关[15]。TFL1蛋白抑制FT下游基因LEAFY(LFY)和 AP1的表达[16],维持花序分生组织状态,抑制花芽分化[17]。tfl1拟南芥突变体叶片减少、开花提前,并且茎尖分生组织转化为顶端花;TFL1过表达,拟南芥的花期延长,次生花序数目增加[17]。
ATC和BFT过表达导致拟南芥晚花,与TFL1基因过表达具有相似的表型,MFT过表达导致开花时间略有缩短,但这些基因的功能缺失突变并没有显示出任何明显的表型[7]。MFT在种子萌发过程中也具有一定的作用[18]。TSF与FT在氨基酸水平高度相似,TSF促进开花,但只有在短日照条件下才有明显的早花表型[5]。
植物成花转变是一个复杂的过程,依赖于基因互作遗传网络的精细调控,FT/TFL1在该过程中起着重要的作用。人们对拟南芥和水稻等模式植物的FT/TFL1研究比较深入,但是在谷子中相关的研究较少。
本试验通过对谷子FT/TFL1基因家族进行生物信息学分析,初步探究FT/TFL1基因家族与成花转变的关系,旨在为深入研究谷子抽穗开花机制提供理论依据。
1 材料和方法
1.1 筛选拟南芥、水稻和谷子FT/TFL1家族成员
在拟南芥中,有6个FT/TFL1家族成员已被报道,利用 NCBI(https://www.ncbi.nlm.nih.gov/)和Phytozome(https://phytozome.jgi.doe.gov/)网站在线搜索拟南芥FT/TFL1家族成员蛋白序列并下载,通过BLAST比对,得到水稻和谷子FT/TFL1家族成员的蛋白序列和启动子序列。
1.2 构建FT/TFL1家族系统发育树
通过MEGA7[19]软件对谷子、拟南芥以及水稻FT/TFL1家族成员进行蛋白序列比对,利用Neighbor-joining算法分析构建系统发育树,选择Bootstrap方法,其他参数选择默认值。
1.3 FT/TFL1家族保守序列分析
利用MEME v4.11.0(http://meme-suite.org/tools/meme)网站在线分析拟南芥、水稻和谷子的保守序列。保守位点的宽度设置为≥6和≤50,最大保守序列鉴定数目设定为10。
1.4 FT/TFL1家族启动子元件分析
利用 PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线分析启动子元件。
1.5 FT/TFL1家族表达模式分析
利用R[20]软件的heatmap函数绘制基因表达热谱图。
2 结果与分析
2.1 FT/TFL1家族系统发育树分析
拟南芥FT/TFL1基因家族中已知的6个成员FT,TSF,MFT,TFL1,BFT 和 ATC 根据其结构和功能分为3个亚类,分别为FT亚类(包括FT和TSF),MFT亚类和 TFL1亚类(包括 TFL1,BFT和ACT 基因)[5,7,21]。
为了分析谷子FT/TFL1家族成员的系统进化关系,利用Mega 7.0软件构建了拟南芥、水稻和谷子的系统进化树。从图1可以看出,谷子FT/TFL1家族可以分为3个亚类,即FT,MFT和TFL1亚类,FT和MFT亚类对开花起促进作用,TFL1亚类抑制开花,同一亚类的基因具有相同或相似的功能,在植物开花过程中共同作用,调控植物开花。

2.2 FT/TFL1家族保守序列分析
利用MEME在线软件对拟南芥、水稻和谷子的FT,MFT和TFL1亚类的蛋白保守位点进行了分析,结果如图2所示,3个亚家族成员均有一段含有50个氨基酸残基的保守序列。FT亚家族的保守基序为X2DPDXPXPSXPX3EXLHWXVTDIPX7GXEX3YE XPXPX2GXH,MFT亚家族的保守基序为MTDPDAP SPSXPXMREX2HWXVX2IPGGTX4GX5YMXPRPXVG IH,TFL1亚家族的保守基序为EX2GXDXRSFFTLX MXDPDXPXPSX5EXLHWXVTXIPGTTDX2FGXE。FT和TFL均有一个决定其功能的关键氨基酸残基,FT蛋白的第85位是一个酪氨酸残基,对控制开花时间具有重要的作用[22],TFL1蛋白第88位的组氨酸对于维持顶端分生组织的生长和延迟开花起主要作用,MFT蛋白的色氨酸也能促进拟南芥开花,但是相对于组氨酸和酪氨酸作用较弱,这几个主要的氨基酸残基与其他残基共同作用,调控拟南芥成花转变[23]。
2.3 FT/TFL1家族启动子序列分析
利用PlantCARE软件在线分析谷子FT/TFL1基因启动子区顺式作用元件,结果发现,除了启动子和增强子区常见的顺式作用元件CAAT-box和核心启动子元件TATA-box外,还具有光响应元件sp1和G-box,胚乳表达顺式作用元件Skn-1_motif以及与逆境胁迫相关的顺式作用元件,如脱落酸响应元件ABRE、低温响应元件LTR和厌氧诱导响应元件ARE等(表1)。说明FT/TFL1基因家族成员在胚乳表达、响应光信号和抵抗逆境过程中起重要作用。

表1 谷子FT/TFL1基因家族启动子顺式元件预测
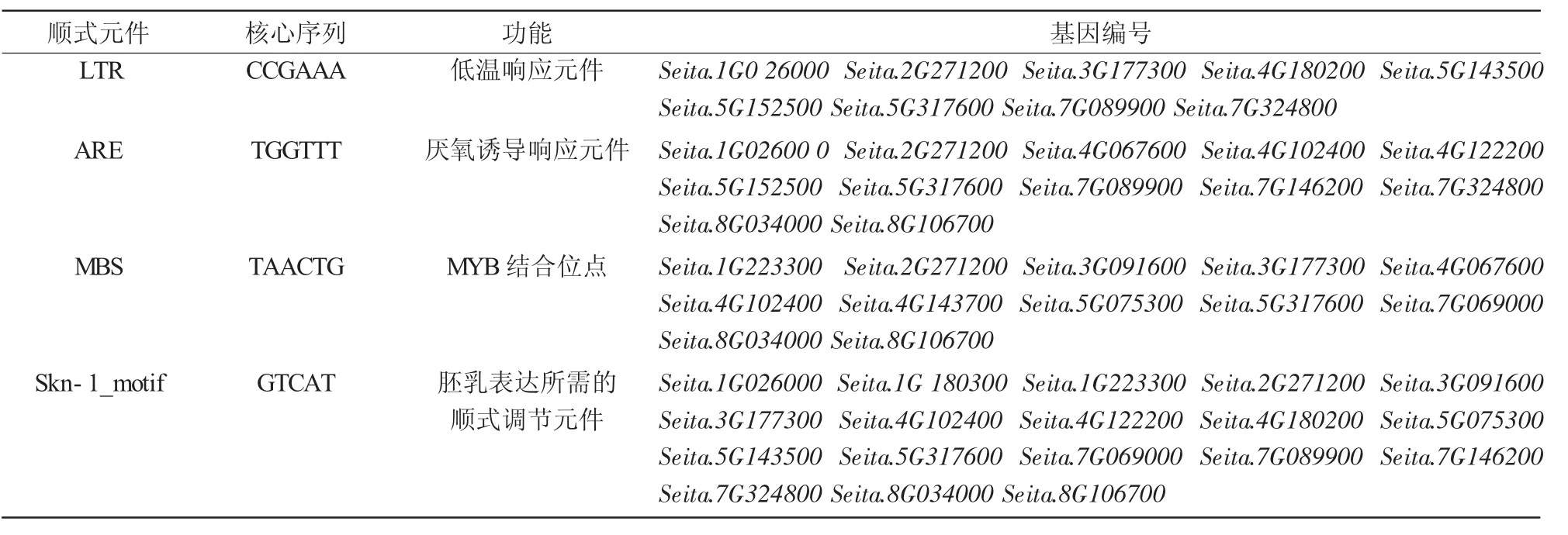
续表1
2.4 FT/TFL1家族表达模式分析
通过NCBI和Phytozome网站检索到29个谷子FT/TFL1基因家族成员,通过Phytozome网站,统计了FT/TFL1基因家族成员在各个组织中的表达量,通过R语言中的heatmap函数绘制热图,结果发现,Seita.4G180200基因在穗中的表达量最高,Seita.4G122200基因在穗中的表达量也较高,Seita.4G122200基因在根中的表达量最高;TFL1亚家族的Seita.1G180300和Seita.7G324800基因在根中的表达量较高,Seita.4G122200的在叶中和苗中的表达量也较高。FT/TFL1家族不同成员在谷子不同组织中的表达量存在显著差异(图3)。

3 结论与讨论
植物FT/TFL1基因是植物由营养生长向生殖生长转变的关键基因,通过对FT/TFL1基因家族进行生物信息学分析,为深入研究谷子FT/TFL1基因家族对成花转变的调控奠定了基础。
通过生物信息学分析发现,谷子FT/TFL1家族可以分为3个亚类,分别为FT亚类,MFT亚类和TFL1亚类,其中,FT亚类和MFT亚类对开花起促进作用,TFL1亚类抑制开花,3个亚类的基因相互协调,共同调控谷子开花。
谷子、水稻和拟南芥的FT,MFT和TFL1亚类的蛋白序列高度保守,均具有一段含有50个氨基酸残基的保守序列,FT蛋白第85位的酪氨酸残基、TFL1蛋白第88位的组氨酸残基分别对调控拟南芥开花和维持顶端分生组织的生长具有重要作用。
通过对谷子FT/TFL1基因家族起始密码子上游约1 500 bp左右的启动子顺式作用元件分析发现,它主要有光响应元件sp1、胚乳表达顺式作用元件Skn-1_motif以及与逆境相关的元件,如低温响应元件LTR等。说明FT/TFL1基因家族成员在胚乳表达、响应光信号和抵抗逆境过程中起重要作用。
通过对谷子表达模式分析发现,Seita.4G122200基因在根、叶和苗中的表达量都最高,Seita.4G180200基因在穗中的表达量最高,FT/TFL1家族在不同组织中的表达量存在显著差异。
综上所述,本研究通过对谷子FT/TFL1家族的生物信息学分析,初步探究了谷子FT/TFL1家族在谷子开花途径中的作用,结果为以后研究FT/TFL1家族成员控制谷子开花的具体调控机制提供了理论依据。
