光照强度对成熟红颜草莓果实着色和花青素生物合成的影响及可能的分子机制
2018-09-01邵婉璐李月灵李钧敏梁宗锁
邵婉璐 李月灵 高 松 李钧敏,3* 梁宗锁
(1.浙江理工大学生命科学学院,杭州 310018; 2.台州学院植物进化生态学和保护浙江省重点实验室,台州 318000; 3.台州学院生态研究所,台州 318000)
植物果实稳定、均匀的着色和香味是果实品质重要的特性。果实色素主要包括花色素、番茄红素、类胡萝卜素等,在自然界中花青素广泛存在于各种植物的花瓣、果实、种子等植物器官中,产生不同的颜色来帮助植物吸引传粉授粉的昆虫,保护植物免于紫外线等非生物的损伤以及病原体等的生物攻击[1~2]。花青素属于黄酮类色素,具有高效的抗氧化活性,可以提高视力[3],还具有预防慢性病如炎症[4]、肿瘤[5]、老龄化疾病[6]、心血管疾病[7]等的生理功效。在自然界中已发现400多种花青素,其中高等植物中普遍存在的有6种[2]。花青素生物合成的代谢途径是类黄酮合成途径中的一个分支,在拟南芥(Arabidopsisthaliana)、玉米(Zeamays)、金鱼草(Antirrhinummajus)以及矮牵牛(Petuniahybrida)等植物中已经被详细解释[8],并从中分离克隆了一些参与花青素合成与调控相关基因[9]。花青素合成途径中包括苯丙氨酸解氨酶基因(PAL),查耳酮合成酶基因(CHS),查耳酮异构酶基因(CHI),二氢黄酮醇4-还原酶基因(DFR),类黄酮-3′羟化酶基因(F3′H),花色苷合成酶基因(ANS)和类黄酮3-O-糖基转移酶基因(UFGT)等关键酶基因。Dubos等[10]对拟南芥花青素的合成途径进行分析,认为主要可以分为三个阶段,第一阶段起始于苯丙氨酸,在苯丙氨酸裂解酶(PAL)、肉桂酸羟化酶(C4H)、香豆酸CoA连接酶(4CL)的作用下生成4-香豆酰辅酶A;第二个阶段是在查尔酮合成酶(CHS)、查尔酮异构酶(CHI)、黄烷酮3-羟化酶(F3H)和类黄酮-3′羟化酶基因(F3′H)的作用下合成二氢黄酮醇的过程;第三阶段是在二氢黄酮醇4-还原酶基因(DFR)、花色苷合成酶基因(ANS)和类黄酮3-O-糖基转移酶基因(UFGT)的催化下生成稳定的花色素苷。而植物花青素的生物合成除了受结构基因的调控外,还受到调控基因的影响。参与花青素合成调控的转录因子主要分为3类MYB、bHLH和WD40,这些转录因子通过与结构基因启动子中相应的作用原件结合,一起调节花青素合成途径中的一个或者多个基因的表达,从而控制花青素的合成。
植物生存的环境对果实着色有着很大的影响,其中光是很重要的影响因子之一。光和花青素生物合成之间的关系是园艺育种与栽培过程中一个重要的研究方向[11]。研究苹果(Malus×domestica)光照强度实验时发现,一天中相对光照大于30%高光照区域,三主枝开心形的单果质量、花青苷含量、硬度、可溶性固形物含量显著高于其他冠形结构[12]。在对兔眼蓝莓(Vacciniumashei)的研究中发现,果实的总酚物质、类黄酮、花青素、维生素C含量整体随光照减弱而降低[13]。遮荫对大樱桃(Cerasuspseudocerasus)的研究结果表明,与对照处理相比,遮荫降低了枝条上果实的可溶性固形物含量和硬度,而且果实着色变差[14]。研究还发现,在果实生长的不同时期对果实进行弱光处理,都会不同程度的降低果实中糖、酸和花青素的含量[15]。有相关研究认为,弱光对果实的影响主要是因为光照不足导致光合作用下降,从而降低了提供给果实的同化物量,最终导致果实的品质降低[16]。
草莓(Fragaria×ananassaDuch)是广泛栽培的重要经济植物。其外观呈果状圆形或心形,具有极高的营养价值,富含丰富的维生素C、花青素等[17],有特殊且浓郁水果芳香,被人们誉为“果中皇后”,深受市场欢迎。研究表明光质对丰香草莓(Fragaria×ananassa‘Toyonoka’)叶片的光合特性有较大影响[18]。而不同光质也会影响草莓果实的香气成分[19]。近年来,草莓大多数花青素合成相关功能基因和转录因子逐步被分离出来,其中Duan[20]等研究表明FpCHS、FpCHI、FpF3H、FpDFR、FpANS、FpUFGT及转录因子MYB10在五叶草莓红果和白果之间的表达有很大差异,表明不同果色的果实中色素相关的基因及转录因子表达强度不同。而有研究表明转录因子MYB10的过表达显著提高了草莓植株的叶、根、果实、和柱头等植物器官的花青素苷水平[21]。但光照强度变化对草莓花青素的合成的调控机制的研究较少。因此利用光照来调控其花青素也成为草莓提高商业价值的重要目标之一。红颜草莓(Fragaria×ananassaDuch. ‘Benihoppe’)生长势强,是我国目前主要的早熟栽培品种,其栽培区占地面积很大,从南到北纬度跨度广,受光照影响较大,因此研究不同光照强度对红颜草莓果实着色、花青素含量及其生物合成相关基因的表达,可为红颜草莓生产中适宜的补光研发提供理论依据,具有重要的实践价值。
1 材料与方法
1.1 试验材料处理及采样
供试材料为栽培红颜草莓(文中简称“草莓”)。实验于2017年4月24日在台州学院温室大棚外试验基地上进行。选取在正常光照条件下完成授粉的草莓,用不同遮荫网进行遮荫处理。用照度计在一段时间内分别不同时间段多次进行测定自然条件下光照和各处理遮荫网下的光照强度,取平均数确定不同遮荫下的透光率。试验设计分别为:自然光照(无遮荫网,透光率100%);轻度遮阴(2层白色遮荫网,平均透光率75%);中度遮荫(1层黑色遮荫网,平均透光率25%)。每种处理材料不少于10株。25 d后,待各处理的草莓成熟后摘取果实,样品采集拍照后,一部分马上用于花青素含量测定,另一部分液氮速冻后存于-70℃保存,用于后续分子实验。每个处理取6个果实,以单株为重复。
1.2 草莓果实花青素含量测定
采用日本岛津高效液相色谱仪(型号LC-20A)进行花青素含量的测定。试样制备[22]:取2.5 g草莓果实放入液氮中研磨成粉末,加入10 mL含0.1% HCl的甲醇,室温萃取24 h,然后在6 650 r·min-1条件下离心15 min,上清液过0.45 μm滤膜,待上样测试。紫外检测器为SPD-20A紫外检测器;色谱柱:ODS-2 C18柱(型号250 mm×4.60 mm 5 μV);紫外检测波长:520 nm;流速:0.7 mL·min-1。柱温:35℃。进样量:20 μL;A相:0.05%甲酸水溶液(体积比);B相:0.05%甲酸+乙腈;梯度洗脱:0~5 min:5%B,5~10 min:5%~10%B,10~25 min:10%~90% B,25~30 min:90~5%B。以标样出峰时间和峰高叠加定性,外标法峰面积定量。每个浓度进样3次,取3次峰面积平均值。以矢车菊素-3-O葡萄糖苷(C3G)(购自上海诗丹德生物技术有限公司)为标准品(图1)进行线性回归。标准曲线为y=2.659 28e-005×x-0.369 148(R2=0.999 180 2),线性关系良好。花青素含量以平均数±标准误表示。
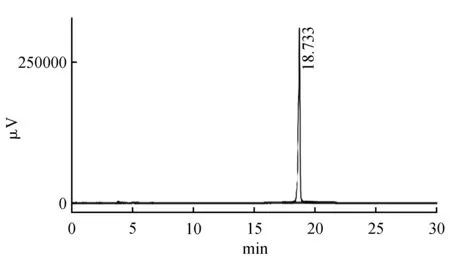
图1 矢车菊素-3-O葡萄糖苷(C3G)色谱图Fig.1 Chromatography profiles of C3G
1.3 草莓花青素代谢途径相关基因的表达分析
1.3.1 RNA提取及cDNA合成
草莓果实总RNA参照RNeasy Plant Mini kit试剂盒(Qiangen)操作进行提取,总RNA提取后,分别用琼脂糖凝胶电泳和Nanodrop超微量分光光度计(Thermo)检测质量和浓度。然后每个样品取5 μL总RNA,参照PrimeScriptTMRT Reagent Kit with gDNA Eraser(TaKaRa)使用说明合成cDNA。使用Primer Premier 5.0软件,设计选取查尔酮合成酶基因(FaCHS)、查尔酮异构酶基因(FaCHI)、黄烷酮-3-羟化酶基因(FaF3H)、类黄酮-3′-羟化酶基因(FaF3′H)、二氢黄酮醇4-还原酶基因(FaDFR)、花色苷合成酶基因(FaANS)、类黄酮-3-葡糖基转移酶基因(FaUFGT)及转录因子MYB10、MYB1为研究对象,分别设计上述基因实时荧光定量PCR引物(表1)[20],引物由上海铂尚生物技术有限公司合成。
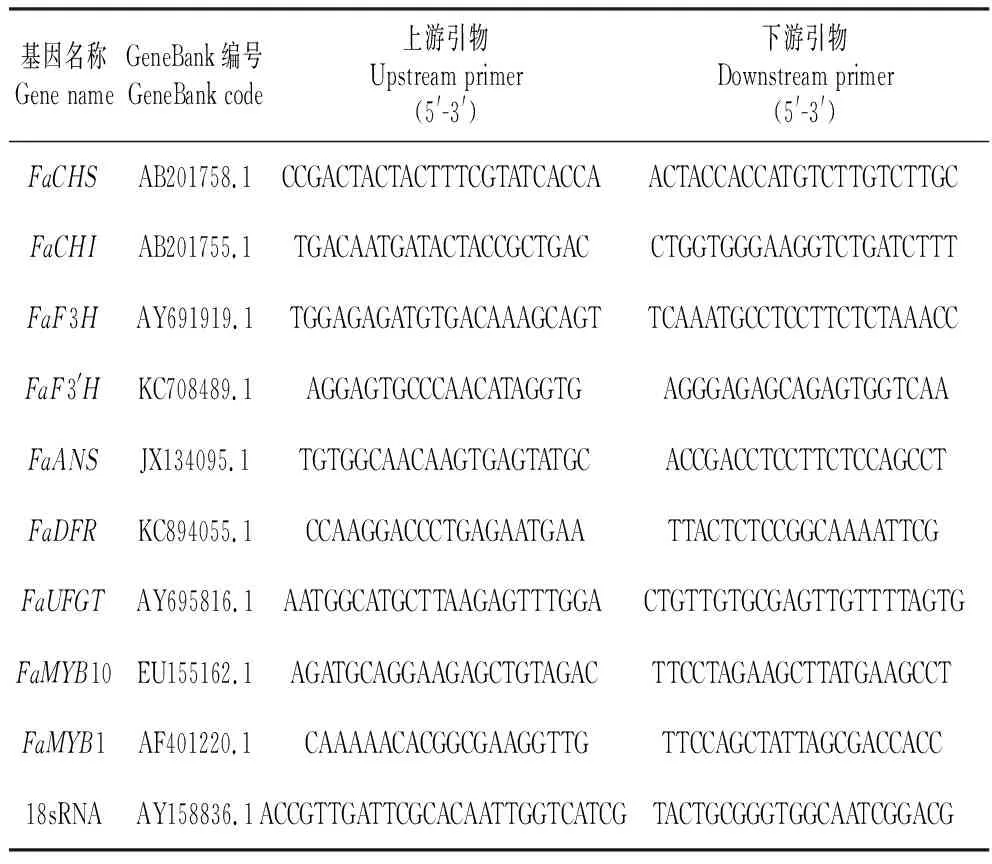
表1 荧光定量PCR引物序列
1.3.2 荧光定量PCR分析
实时荧光定量PCR在Eco定量PCR仪器(美国Illumina公司)上完成,参照SYBR Premix试剂盒(大连宝生物)的使用说明进行操作。反应总体积为:10 μL:5 μL 2×SYBR qPCR Mix,上、下游引物各0.2 μL(10 mmol·L-1),1 μL cDNA模板和3.6 μL灭菌双蒸水。反应条件:95℃预变性30 s;40个循环:95℃ 5 s,60℃ 30 s,72℃ 30 s。以草莓18S rRNA为内参基因[20],每个样品设3个重复。采用2-△△CT方法[23]计算FaCHS、FaCHI、FaF3H、FaF3′H、FaANS、FaDFR、FaUFGT基因以及转录因子FaMYB10、FaMYB1的表达量,所有结果重复3次,以平均数±标准误表示。
1.4 数据分析
采用单因素方差方法分析不同处理间草莓色素含量及基因表达量的差异性。采用最小显著极差法进行不同光照强度处理之间的多重比较。采用Pearson相关系数分析色素含量与基因表达的相关性。
2 结果与分析
2.1 光照对草莓果色的影响
光照强度影响草莓果实的着色,随着光照强度的下降草莓果实的颜色由红色逐渐减弱,低透光率下果色呈现浅色(图2)。

图2 不同光照处理下草莓果实着色的影响 A. 100%透光下的果实;B. 75%透光的果实;C. 25%透光的果实Fig.2 Effects of different light conditions on the coloration of strawberry fruits A. Fruit of 100% transmittance; B. Fruit of 75% transmittance; C. Fruit of 25% transmittance
2.2 光照对草莓果实花青素含量的影响
HPLC检测显示3个处理在520 nm波段处均检测到花青素类物质,与标准品比对后确认为C3G。根据参考标样的滞留时间和峰面积可以看出,随光照强度降低草莓果实积累的花青素含量降低,草莓果实颜色越深色素积累含量越高。75%和25%透光率下草莓果实合成的花青素含量分别为100%透光率下的58.42%和7.46%(图3)。
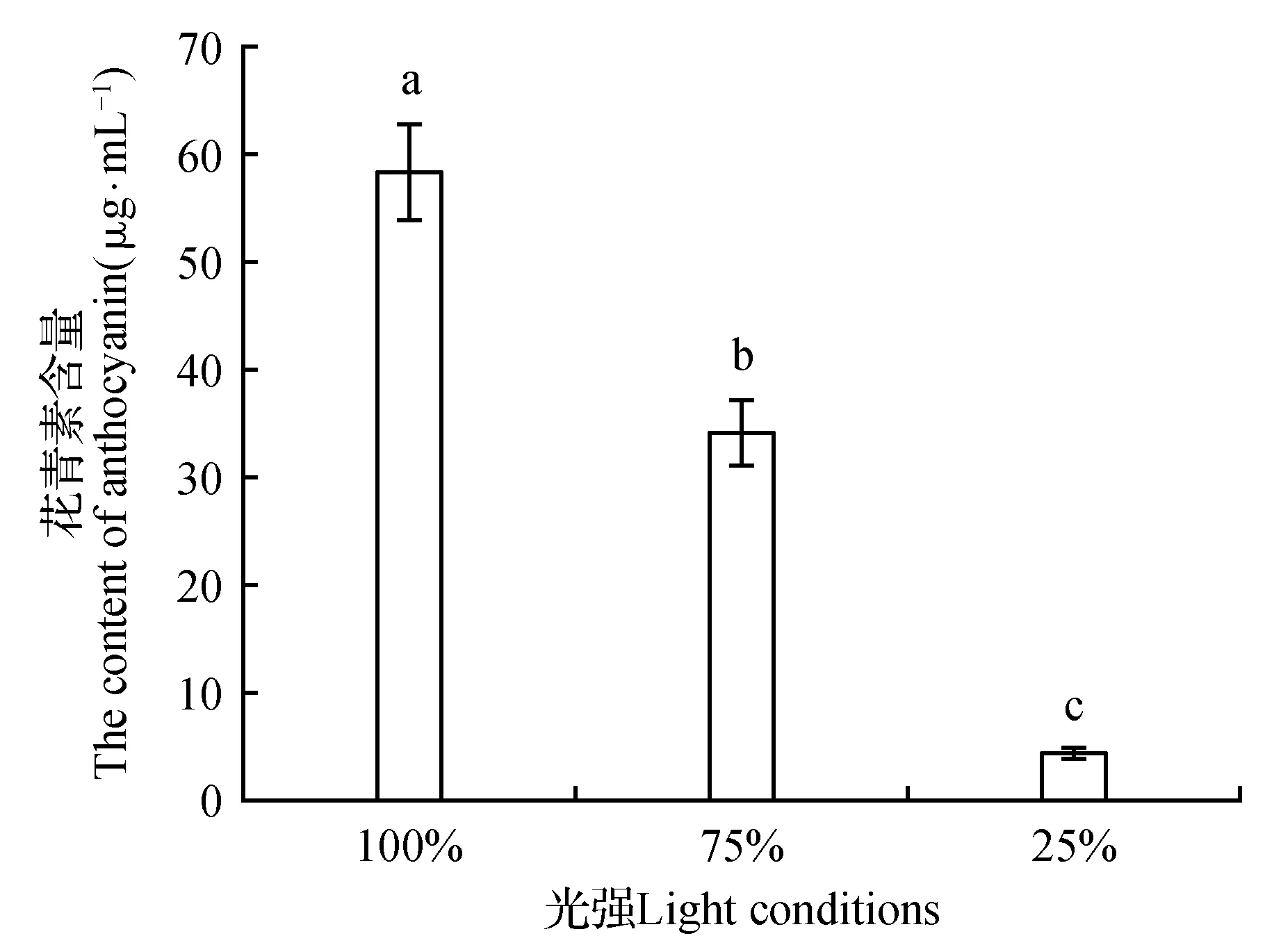
图3 不同光照下草莓果实花青素含量 100%. 透光率100%;75%. 透光率75%;25%. 透光率25% 不同字母表示不同处理之间存在显著性差异,下同。Fig.3 The content of anthocyanin in strawberry fruits under different light conditions 100%. Light transmittance 100%; 75%. Light transmittance 75%; 25%. Light transmittance 25% Different small letters indicate there is significant difference between different treatments,the same as below.
2.3 光照对草莓果实花青素合成相关基因表达的影响
如图4所示,FaCHS、FaCHI、FaF3H、FaF3′H、FaDFR、FaANS、FaUFGT等基因的表达均随光照强度的降低而下降,在75%透光率下各基因表达分别下降了41.87%、41.91%、46.47%、39.69%、24.57%、41.02%、31.81%,而在25%透光下各基因表达分别下降了39.21%、62.81%、71.57%、87.74%、46.64%、57.26%和50.31%。
2.4 光照对草莓果实花青素合成相关转录因子表达的影响
如图5所示,转录因子FaMYB10及FaMYB1在草莓果实中的表达也随光照强度降低而降低,FaMYB10在75%透光和25%透光下的表达率相对于100%透光下分别下降了25.85%和46.24%,而FaMYB1则分别下降了39.49%、35.47%。但FaMYB1在75%和25%透光下的表达差异不显著。
2.5 色素含量与基因表达量的相关性分析
从表2可以看出,在不同光照处理下草莓果实花青素(C3G)含量与基因FaDFR和FaUFGT的相关性达到显著水平;与FaF3′H的相关性达到极显著水平;而与其它基因FaCHS、FaCHI、FaF3H和FaANS以及转录因子FaMYB10和FaMYB1相关性不显著。
3 讨论
花卉、果实丰富的颜色是由花色素苷决定的,不稳定的花色素形成后,在UFGT的作用下进一步与葡萄糖糖苷形成稳定的花色素苷。而光照是影响着色的重要环境因子。孟祥春等[24]对非洲菊(Gerberehybrida)花色素苷积累的研究结果也表明阻断光照后,花不能正常着色。本研究发现遮光条件可以影响草莓果实的着色和花青素的含量,随着光照强度的降低,草莓成熟果实中花青素含量也减少,这表明光照的强弱是影响草莓果实着色和花青素合成的关键因子之一。
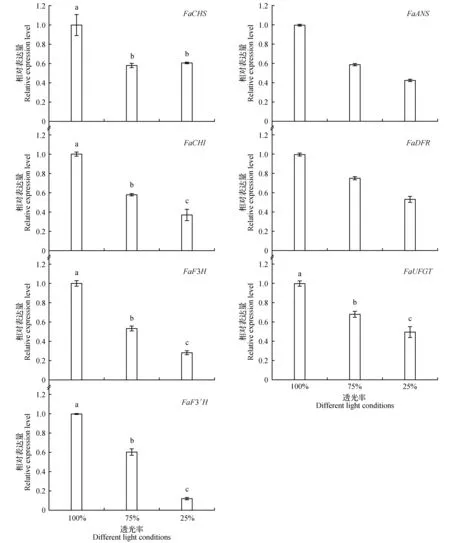
图4 不同光照下草莓果实花青素合成相关基因的表达Fig.4 Expression of anthocyanin biosynthesis related gene in strawberry fruits under different light conditions
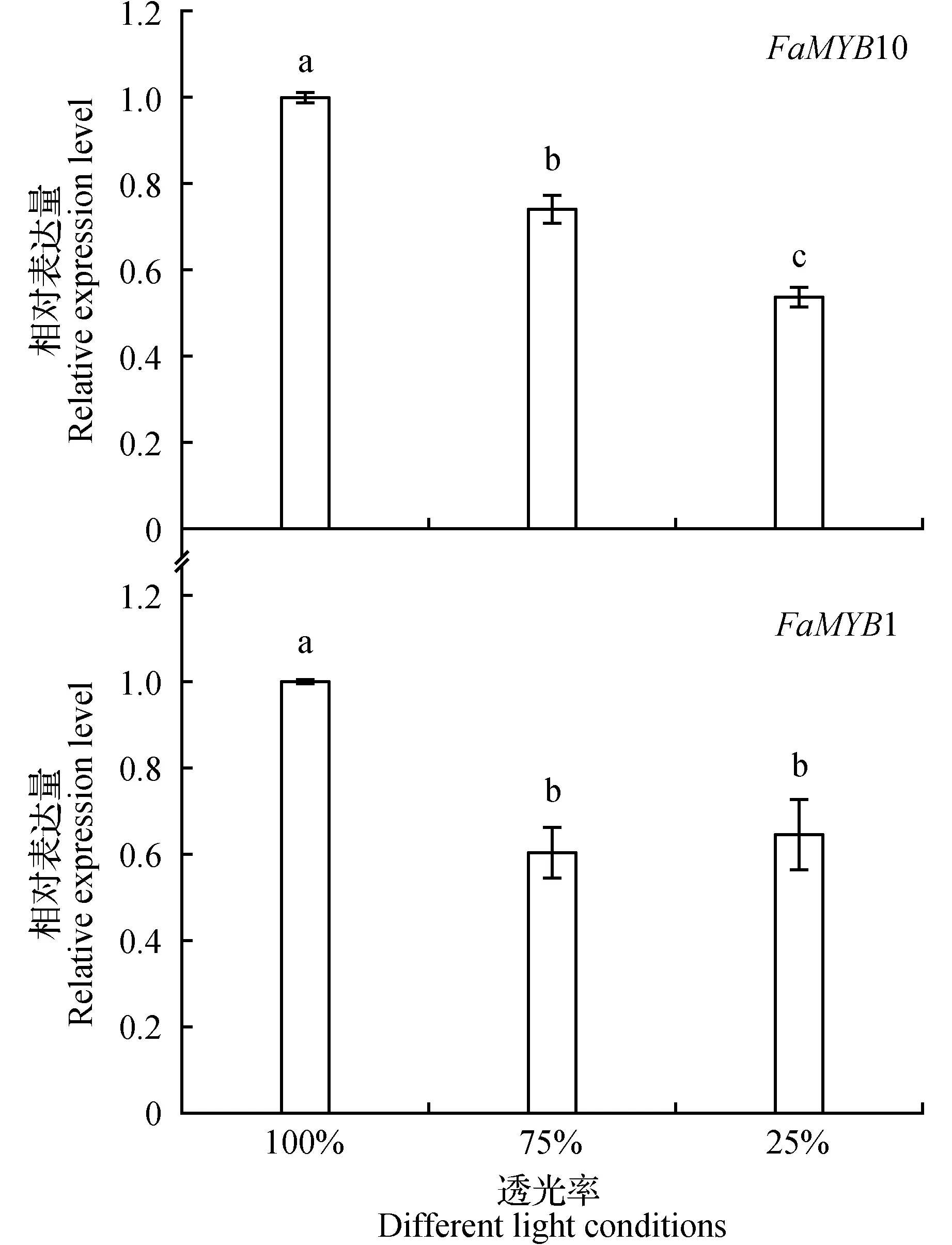
图5 不同光照下草莓果实花青素合成相关转录因子的表达Fig.5 Expression of anthocyanin biosynthesis related transcription factors in strawberry fruits under different light conditions
对花青素合成途径相关基因的表达分析显示,光照对草莓果实花青素合成过程中涉及到的酶基因FaCHS、FaCHI、FaF3H、FaF3′H、FaANS、FaDFR、FaUFGT以及转录因子FaMYB10、FaMYB1的表达均有影响。周波[25]研究草莓果实对光的响应时发现,光敏感性不同的草莓果实在花青素合成过程中最终合成的花青素含量不同,花青素合成相关基因CHS、DFR、ANS在光不敏感型草莓果实“女峰”果实遮光部分和受光部分的表达没有明显差异,而部分光敏感型草莓“丰香”果实遮光部分比受光部分表达量减少。本实验中得到的FaCHS、FaDFR、FaANS在不同光照下的表达结果与光敏感型草莓结果一致,表明本实验采用的红颜草莓为一种光敏感型草莓。C3G是一类3′-4′-羟化化合物,而F3′H是控制花青素B环羟化的重要功能酶,F3′H基因的上调会导致Penstemonbarbatus其花从蓝色到红色的颜色变化[9],是C3G和花翠素合成的重要决定因子。本研究中,在强遮光条件下FaF3′H基因的表达量呈现出显著下调,说明草莓的果实的着色与其密切相关。DFR处于花青素代谢途径中的中下游,催化二氢黄酮醇形成不稳定原花色素,我们发现FaDFR基因表达与色素含量呈现显著相关性,这与Espley RV[26]及Yoshio Itoh[27]等报道的DFR基因的表达活性与苹果果皮色素含量呈紧密相关性相一致。UFGT是花青素合成过程中最后一个的关键酶,主要作用是在花青素形成后对碳骨架进行糖基化修饰[28],它能使不稳定的花青素通过糖基化作用转变为稳定的花青苷,在色素形成过程中发挥重要作用。我们结果显示FaUFGT基因的表达同色素含量呈现显著相关性,这一结果同一些在苹果、梨、葡萄中的报道也相一致[29~30]。FaF3′H、FaDFR和FaUFGT基因表达都和C3G的合成密切相关。色素合成途径中各基因的表达趋势在果实的不同发育阶段常不同[31],进一步的研究可以分析不同光照条件下草莓不同发育阶段果实花青素含量及与合成相关基因表达的相关性,以深入阐明光照对草莓花青素合成的影响分子机制。
表2不同光照强度下草莓色素含量与花青素合成基因的相关性分析
Table2Thecorrelationcoefficientsbetweenthecontentofanthocyaninandtherelativeexpressionoftheanthocyaninsynthesisrelatedgenesunderdifferentlightconditions

指标IndexFaCHSFaCHIFaF3HFaF3'HFaANSFaDFRFaUFGTFaMYB10FaMYB1色素Pigment0.8030.9690.9740.999**0.9540.996*0.994*0.9920.781
注:*表示0.05显著差异;**表示0.01极显著差异
Note:*indicates the significant level of 0.05;**indicates the significant level of 0.01
植物中对花青素合成途径中起调控作用的重要转录因子主要有MYB、bHLH和WD40,这些转录因子与结构基因的启动子结合,激活或抑制花青素生物合成途径中一个或多个结构基因的表达,影响色素的合成[32~33]。Yao等[34]通过对梨(Pyrus)果皮花青素合成的研究验证了PyMYB114的功能,发现转录因子PyMYB114和PyMYB10的功能存在着互作及叠加的效应,这两种基因在烟草和草莓上的共转化导致花青素的生物合成增强。目前,和草莓花青素合成有关的报道中,涉及到的MYB转录因子主要有FaMYB1和FaMYB10。Wang等[21]研究表明FaMYB10在烟草叶片体系中可直接激活拟南芥DFR基因。Medina-Puche等[35]研究报道MYB10基因在草莓红色果实发育过程中,与色素含量呈现显著正相关,且可影响花青素合成基因FaDFR的表达。Kadomura-Ishikawa等[36]发现FaF3′H基因的表达量也受到FaMYB10的调控。Ai等[37]研究菊花花青素调控关键基因时发现,将RsMYB1基因转入到矮牵牛中时,高表达的RsMYB1基因可以显著提高花青素生物合成过程中的结构基因的表达,从而增加菊花花瓣等植物器官中的花青素含量。这些结果表明草莓中的FaMYB10和FaMYB1转录因子在花青素合成途径上对这些结构基因有着直接或间接作用。而在我们的结果中在100%透光率下草莓果实色素含量最高且成熟度最高时转录因子FaMYB10及FaMYB1表达量最高,说明光照对转录因子也存在着显著的作用进而影响到了相关基因的表达。遮光实验说明光照对草莓中的影响可能是通过相关的光受体,进而影响转录因子的活性[36],最终影响了花青素的合成。本研究也发现了FaMYB1的表达在25%和75%光照强度下没有显著性差异,表明其与FaMYB10对花青素合成途径相关基因的调控及阶段可能不同,应在后续的研究中作为重点。
综上所述,红颜草莓为光敏感型草莓品种,光照强度可对其成熟果实的着色与花青素含量产生显著影响,其可能是通过转录因子影响花青素合成相关基因的表达来实现。红颜草莓栽培种在果实发育期需注重光照,在光线不足的情况下需人工补光以获得较好的果实品质。另外草莓中花青素的合成代谢具有复杂的网络调控机制,相关转录调控机制还需要进一步研究。
