利用GBS-cpDNA揭示花生属花生区组内种间亲缘关系
2018-08-29李春娟闫彩霞石程仁赵小波单世华
李春娟,闫彩霞,石程仁,赵小波,王 娟,单世华
(山东省花生研究所,山东 青岛 266100)
栽培种花生是大约3500年前由二倍体AA(A.duranensis)和BB(A.ipaensis)野生种基因组重组进化形成的异源四倍体植物[1]。在植物分类系统上,栽培种花生(ArachishypogaeaL.)隶属于豆科(Leguminosae),蝶形花亚科,花生属(Arachis)。目前,花生属已经命名的有81个种,绝大部分是二倍体野生种(2n=20),少量四倍体种(2n=40)和非整倍体种(2n=18)。1994年,Krapovickas和Gregory根据近代分类学研究结果,将花生属植物进一步划分为9个区组(花生区组、大根区组、直立区组、围脉区组、异形花区组、匍匐区组、根茎区组、三叶区组和三子粒区组)。其中,花生区组(sect.Arachis)包含花生栽培种(A.hypogaeaL,2n= 40)、 25个二倍体野生种和一个四倍体野生种,研究表明它们之间杂交亲和[2-3]。
栽培种花生是我国重要的经济作物和油料作物。我国是花生种植大国,已有500多年的种植历史。孙大容等发表的《中国栽培花生品种的分类》中指出,我国种植的花生品种主要由2个亚种组成,即密枝亚种ssp.hypogaea和疏枝亚种ssp.fastigiata[4]。栽培种花生种植面积广泛,但是遗传基础相对狭窄,因此拓宽栽培种花生的种质资源遗传基础显得尤为重要[5]。花生属野生种质资源丰富,基因型多样,在热带亚热带气候环境下有广泛的适应性。和栽培种花生相比,野生种质具有叶斑病、锈病、青枯病等抗病虫害基因,是用于花生品种改良的重要基因源。尽管有些种质资源并无栽培价值,但可根据不同的目的,通过杂交选育利用其有利基因,获得新种或达到改良现有栽培种的目的。对花生属种间亲缘关系的准确揭示将有助于实现这一目标。
简化基因组测序(Genotyping-by Sequencing;GBS),是一种高通量、高性价比的二代测序技术。通过对花生属的不同物种进行测序,获得酶切位点附近基因序列信息,能够检测出大量的遗传变异位点[5-7]。本研究利用GBS测序结果获取能够比对到叶绿体全基因组的序列信息,由于叶绿体基因组具有单倍性、母系遗传、结构和功能高度保守等特点,因此通过比较花生栽培种与其近缘野生种叶绿体基因组序列差异,能够揭示系统演化关系。同时,叶绿体基因组有一些高突变区,在漫长的进化中会有差别,可以解决低阶分类单元问题。此外,Prabhudas等首先发表了一个花生栽培品种(A.hypogaeaL. Co7 variety)的叶绿体全基因组,指出基因组大小有156 391bp,注释到110个基因,这为花生属各物种叶绿体遗传信息的获得提供了参考信息[8]。因此,本研究能够较好地揭示栽培种花生与花生属花生区组其他野生种之间的亲缘关系,对野生资源的引入和遗传资源的保存有重要参考。
1 材料与方法
1.1 试验材料
55份花生种质材料包括11份花生区组的野生种质(A.monticola,A.duranensis,A.stenosperma,A.paraguariensis,A.helodes,A.hoehnei,A.chacoensis,A.villosa,A.correntina,A.batizocoi,A.cardenasii),11份来自印度、美国及阿根廷等地的国外种质以及33份(普通型10份,龙生型9份,珍珠豆型8份,多粒型6份)国内不同产区的花生种质材料。每份材料随机选取3粒完整种子,培养皿中浸种1~2 d后,置入培养盒,28℃下培养10 d。使用植物基因组DNA提取试剂盒(天根,北京),依次取每个样品的鲜叶进行基因组DNA的提取并检测DNA的完整性和纯度(单位浓度不少于50 ng/μL,总质量大于2μg;相关试剂来自US Everbright Inc.)。-80°C下保存备用。
1.2 文库构建、高通量测序及原始测序数据过滤
首先,选取EcoRI和NiaIII酶切组合对质量合格的DNA进行酶切[9]。其次,GBS建库,酶切好的每个样本加不同的一级接头(adapter),区分混合测序中不同样本后,再通过PCR加入二级接头(adapter),并且通过控制PCR的退火时间,控制扩增片段大小(300~500 bp),实现片段选择的目的。最后,确认检测建库质量后,在Illumina Hiseq 4000平台上进行PE150双末端测序。
利用Q30标准对序列质量进行评估,采用FastQC软件(http://www.bioinformatics.babraham. ac.uk/projects/fastqc/)对所测得的原始数据进行过滤:①去掉接头;②去掉N的含量超过该条read长度比例 10%的reads;③去除单端序列低质量碱基(Q≤5)比例超过read长度 50%的reads。
1.3 全基因组水平遗传变异位点信息的获取
以花生叶绿体基因组(Genbank号为:KX257487)为参考基因组,全长为156391 bp。利用 BWA version 0.6.2[10]软件进行reads比对(mem-t4-k32-M)。输出文件通过SAM tools 软件和VCF tools软件获取叶绿体全基因组上的遗传变异位点。其中,参数“view”转化sam为bam文件;参数“sort”对bam文件进行排序;参数“rmdup”去除重复比对的reads(reads比对到多个位置);参数“mpileup”检测SNP。为了提高遗传变异位点的准确性,按照以下标准进行过滤:①双等位基因;②Q>20;③MAF(Minor Allele Frequency)≥0.05;④个体覆盖度不低于80%;⑤reads深度≥3。
1.4 PCA分析及系统树构建
R软件包中adegenet程序用于主成分判别分析(Discriminant Analysis of Principal Components,DAPC)[11]。在不设置种群的先验信息情况下,利用参数“find.clusters”在贝叶斯信息准则基础上(BIC)寻找最合理的簇数目。利用MEGA最大似然法(Maximum Likelihood,ML)和邻接法(Neighbor-Joining,NJ)两种方法分别构建系统进化树,缺隙(Gap)设为缺失状态,同时以自展法(Bootstrap)进行检验,重复5000次。其他均使用默认参数。
2 结 果
2.1 GBS测序和遗传变异位点检测
GBS测序reads与叶绿体全基因组比对,reads平均深度达到231x(113x ~922x),平均覆盖度为69%(54%~88%)。从55份花生材料中获得遗传变异位点1102个。在此基础上,经过上述过滤标准,剩余SNP位点约372个。此外,去除杂合SNP位点,共保留121个变异位点,其中包含10个InDel位点,111个SNP位点。筛选得到的SNP变异位点将用于后续分析。
2.2 系统演化关系揭示
首先,PCA分析将55份花生区组的材料分成了5个组: ① 栽培种花生和A.monticola;②A.duranensis,A.batizocoi,A.paraguariensist,A.correntina和A.hoehnei;③A.villosa和A.stenosperma;④A.chacoensis;⑤A.helodes和A.cardenasii。
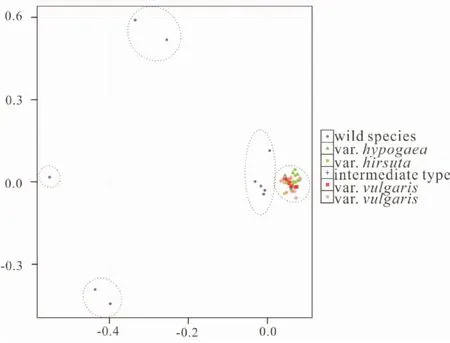
图1 PCA分析显示五个相似的遗传组分 Fig. 1 PCA analysis revealed five genetic groups
其次,构建了最大似然法(ML)和邻接法(N-J)两棵系统演化树。与花生区组的其他物种相比,栽培种花生种内遗传变异较少,聚为平行枝。该枝还包括野生四倍体A.monticola。此外,野生种A.duranensis,A.batizocoi,A.paraguariensist和A.hoehnei聚为一枝,其余6个野生种聚为一枝。与NJ树不同的是,ML树的平行枝中还包含野生种A.correntina。
3 讨 论
花生属花生区组种间亲缘关系的揭示对利用野生种优良基因改良栽培种花生具有重要意义。研究提出,区组内种间杂交具有一定亲和性,区组间种间杂交不亲和或杂种完全不育[2]。由于种间形态差异的复杂性,通过细胞水平和分子水平揭示它们之间的亲缘关系更为准确[12]。唐荣华对花生属六个区组九个种进行核型分析,根据着丝点位置区分花生遗传距离。指出A.batizocoi与花生区组中其它二倍体核型差异较大,遗传距离较大[13]。Singh从种子蛋白质电泳谱带分析中指出花生区组大多数二倍体种都有一个共同的A染色体组,而A.batizocoi则是B染色体组[14]。Moretzsohn等根据SSR聚类结果分析,认为栽培种花生与花生区组的二倍体种A.duranensis、A.villosa和A.diogoi的关系较近[15]。唐荣华等用SSR和AFLP两种分子标记技术证明A.cardenasii、A.batizocoi与栽培种的亲缘关系较远,不大可能是花生栽培种的祖先,而认为A.duranensis是花生栽培种的祖先之一[16]。杨森以花生栽培种四粒红及其与野生种光叶花生的种间杂种为材料,根据ITS1-5.8S-ITS2序列扩增获得长度约800bp的单一条带,在对 PCR 产物进行克隆测序的基础上构建了花生属进化树,具备鉴定种间杂种的优势[17]。
由于叶绿体基因组具有单倍性、母性遗传、结构和功能高度保守等特点,使得通过比较栽培种花生与其近缘野生种叶绿体基因组序列差异成为阐明种间亲缘关系的理想研究方法。本研究聚类结果表明,44份栽培种花生材料尽管来自不同国家,属于不同花生类型,但种内遗传变异程度较小,明显聚在一起。不同的野生种间遗传距离不同。花生属中与栽培种花生亲缘关系最近的是A.monticola。A.monticola是花生区组中唯一的四倍体野生种,基因组类型AABB。Grabiele et al. 推测这个物种是通过自然杂交和自然加倍后,但未经人工选择的四倍体物种,即可能是栽培种花生祖先种[18]。亲缘关系较近的有A.duranensis,A.batizocoi,A.paraguariensist和A.hoehnei。其中,A.duranensis被证实是栽培种花生的野生亲本之一(A染色体组供体)[1]。通过上述文献报道推测A.batizocoi与另一个亲本(A.ipaensis)的亲缘关系较近,因为它们都具备B染色体组。系统树结果也支持以上聚类关系。
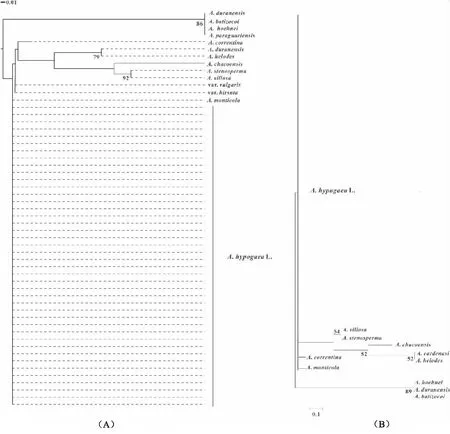
图2 花生属花生区组内种间亲缘关系树 Fig. 2 Phylogenetic tree among the species of sec. Arachis 注:(A)N-J树;(B)ML树。 Note: (A) N-J tree; (B) ML tree
由于花生区组存在杂交亲和性,根据栽培种花生与野生种亲缘关系远近,通过杂交、选育利用其有利基因,能够获得新种或改良现有栽培种,从而实现对野生资源的引入和遗传资源保存的目标。
