植物内生菌活性代谢产物最新研究进展
2018-08-23张晓梅李文均
靳 锦, 赵 庆, 张晓梅*, 李文均
(1.云南中医学院 基础医学院 中药学院,云南 昆明 650500;2.中山大学生命科学学院 广东省热带亚热带植物资源实验室,广东 广州 510275)
植物内生菌(Endophyte)一般是指那些在其生活史的一定阶段或全部阶段生活在健康植物的组织内但不会引起宿主植物产生侵染现象或明显病害症状的一类微生物[1]。1898年Vogle[2]从黑麦草(Loliumtumeletum)种子中分离出第一株内生真菌,1993年Stierle等[3]从短叶紫杉的内生真菌中发现紫杉醇(Taxol),内生菌的研究自此引起了国内外学者的重视。内生菌是一类非常重要的微生物资源,在自然界中广泛存在,几乎每一种植物中都能发现内生菌,它们具有独特的生理和代谢机制,使其能够适应植物内部特殊的环境,同时还能够编码产生多种生物活性物质。此外,内生菌因长时间与宿主协同进化使其产生某些与宿主植物相似或相同的具有药用价值的代谢物质。有学者对植物内生菌次生代谢产物进行整理和总结[4-8],但多数仅涉及特定的活性或结构类型。为了更好地研究和开发植物内生菌次生代谢产物的活性成分,本文对2010年至今从植物内生菌中分离得到的具有新颖结构或生物活性的化合物研究进展进行综述。
1 植物内生菌的次生代谢产物
研究发现,内生菌分布广、种类多,几乎存在于所有研究过的植物(含药用植物)中,研究内生菌代谢产物所涉植物来源较为广泛。近年来从植物内生菌发酵产物中分离得到的代谢产物结构类型多样,包括生物碱、甾体、萜类、蒽醌类、环肽类、黄酮类等,其中不乏与宿主植物次生代谢产物结构类似的,如表1所示。目前,对植物内生菌代谢产物的研究大多集中在真菌和放线菌,细菌较少。其中,研究较多的植物内生真菌类群主要包括拟茎点霉菌属(Phomopsis)、镰刀菌属(Fusarium)、青霉菌属(Penicillium)、曲霉属(Aspergillus)和毛壳菌属(Chaetomium)等,少见拟层孔菌属(Fomitopsis)、链格孢属(Alternaria)及单格孢属(Ulocladium),近年来植物内生真菌次生代谢产物研究见表2。植物内生放线菌仍以链霉菌属(Streptomyces)为研究最多的类群,链霉菌属也是活性化合物出新率较高的类群;此外,小单孢菌属(Micromonospora)、诺卡氏菌属(Nocardia)和拟诺卡氏菌属(Nocardiopsis)等也是植物内生放线菌代谢产物研究中较常见的类群,表3归纳了近年来分离自植物内生放线菌的主要代谢产物。不难看出,植物尤其药用植物内生菌中有大量结构新颖的代谢产物有待于进一步的研究和发现。

表1 与宿主结构相同或相似的次生代谢产物及其活性
注:表中标【药】的原植物为药用植物,下表同

表2 源自植物内生真菌的次生代谢产物

宿主植物内生真菌次生代谢产物茶茱萸科植物(M. dentate)拟层孔菌Fomitopsis sp. MTCC 10177Camptothecine (11)、9-MeO-Camptothecine (12)、10-OH-Camptothecine (13)[18]槲寄生【药】(Viscum coloratum)毛双孢菌Lasiodiplodia sp. ME4-2cyclo-(Trp-Ala)、indole-3-carboxylic acid、indole-3-carbaldehyde、mellein、2-phenylethanol[19]胡黄连【药】(Picrorhiza kurroa)大豆茎溃疡病菌Diaporthe phaseolo-rum PR4薄荷醇、苯乙醇、柠檬烯、3-戊酮和3-羟基丙酸[20]龙血树属植物(Dracaena draco L.)可可球二孢Botryodiplodia theobro-mae Pat生物碱、香豆素、异香豆素、倍半萜、二肽、酰胺类,(14)为结构新颖的生物碱[21]苦橙(Citrus aurantum L.)顶多毛孢菌Bartalinia pondoensis MarinN-(ethyloxy, hydroxymethyl) phenylethyl-amine[22]甘草属植物(Glycyrrhiza glabra Linn.)茎点霉菌Phoma sp. GG1F1硫代二酮哌嗪类化合物(15、16)[23]红海榄(Rhizophora stylosa)产黄青霉Penicillium chrysogenum PXP-55脑苷脂类化合物(17)、Chrysogendones A(18)和B[24]美国白松(Pinus strobus)DAOM 242779和242780呫吨类化合物、苯并呋喃类化合物(19~23)[25]诃子【药】(Terminalia chebula Retz.)链格孢霉Alternaria alternateAltenusin(24)[26]香露兜(Pandanus amaryllifolius)间座壳菌Diaporthe sp. P133二氢苯并吡喃酮化合物(25、26)[27]马钱属植物(Strychnos cf. toxifera)粘帚霉Gliocladium sp. CR 2.3.1二酮哌嗪生物碱(27)[28]银杏【药】(Ginkgo biloba)多节孢菌Nodulisporium sp. A21Sporothriolide(28)[29]仙人掌(Opuntia dillenii)镰刀菌Fusarium sp. 伊快霉素衍生物(29)[30]瓶花木(Scyphiphora hydrophyllacea Gaertn. F.)红树植物内生真菌A1 (unidentified)A Fatty Acid Glycoside(30)[31]云实属巴西木(Caesalpinia echinata Lam.) 镰刀霉菌Fusarium sp. KF 611679白僵菌素[32]蒜【药】(Allium sativum)脐孢木霉Trichoderma brevicompac-tum 0248木霉素[33]木榄(Ruguiera gymnorrhiza)曲霉Aspergillus sp. Aspergillumarin A(31)和B[34]地衣(Lichens)单格孢菌Ulocladium sp. 联苯类化合物:Alterlactone(32)、Altenusin和5'-methoxy-6 -methyl-biphenyl-3,4,3'-triol[35]番红花【药】(Crocus sativus)青霉菌Penicillium vinaceum X17异喹啉类生物碱(33)[36]苦楝【药】(Melia azedarach L.)烟曲霉Aspergillus fumigatus LN-43-Hydroxyfumiquinazoline A(34)[37]罗勒属植物(Ocimum tenuiflorum L.)橘青霉Penicillium citrinum生物碱(35、36)[38]生地黄【药】(Rehmannia glutinosa)轮枝孢菌Verticillium sp.联苯醚类化合物(37)[39]孔石莼(Ulva pertusa)球毛壳菌Chaetomium globosum QEN-14Cytoglobosins A-G(38)[40]未知来源(A dead branch collected from Mt.)GassanNeonectria ramulariae Wollenw KS-246吡咯烷类生物碱(39)[41]

续表2
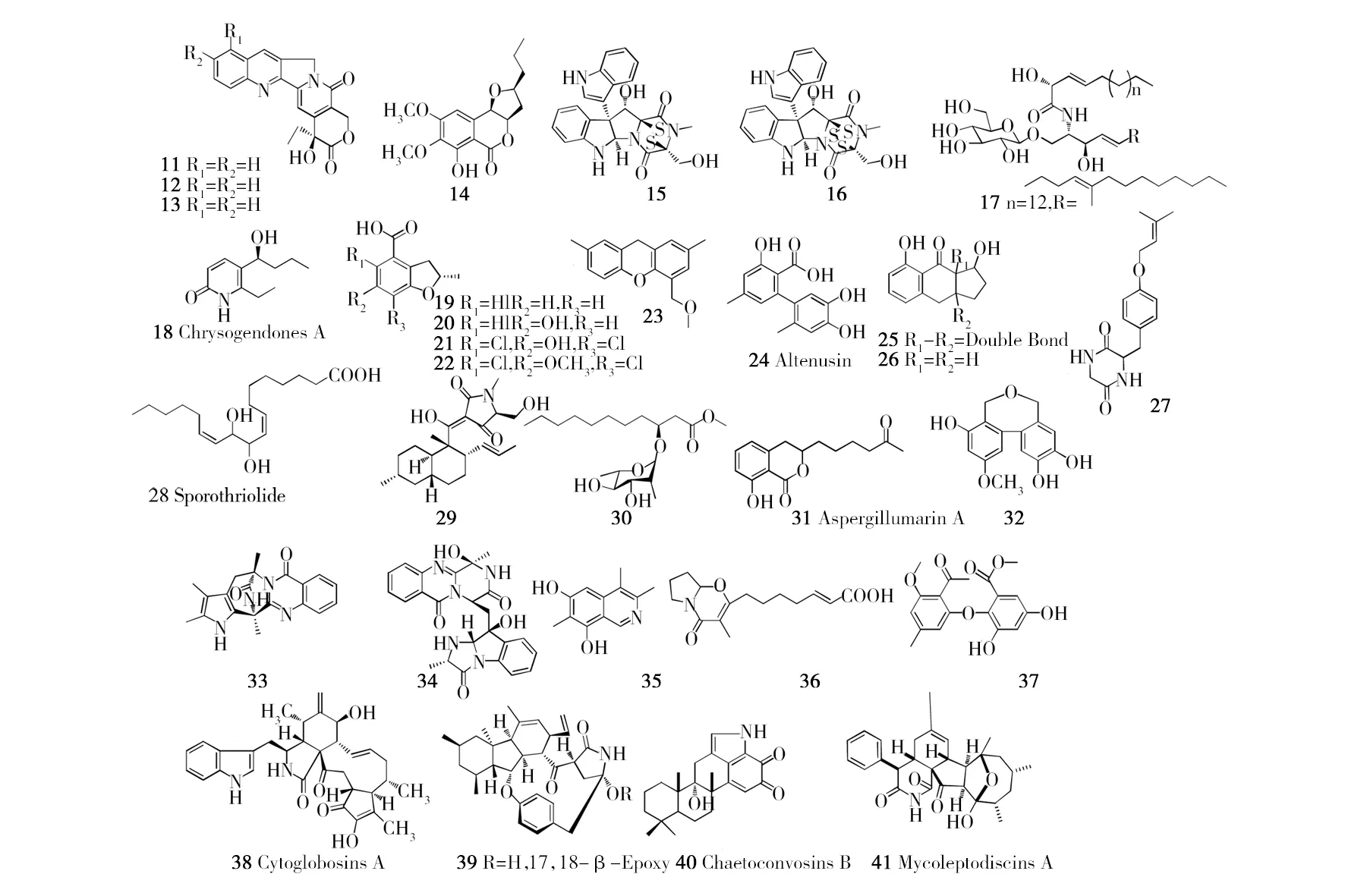

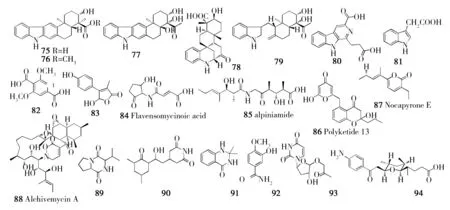
表3 源自植物内生放线菌的次生代谢产物
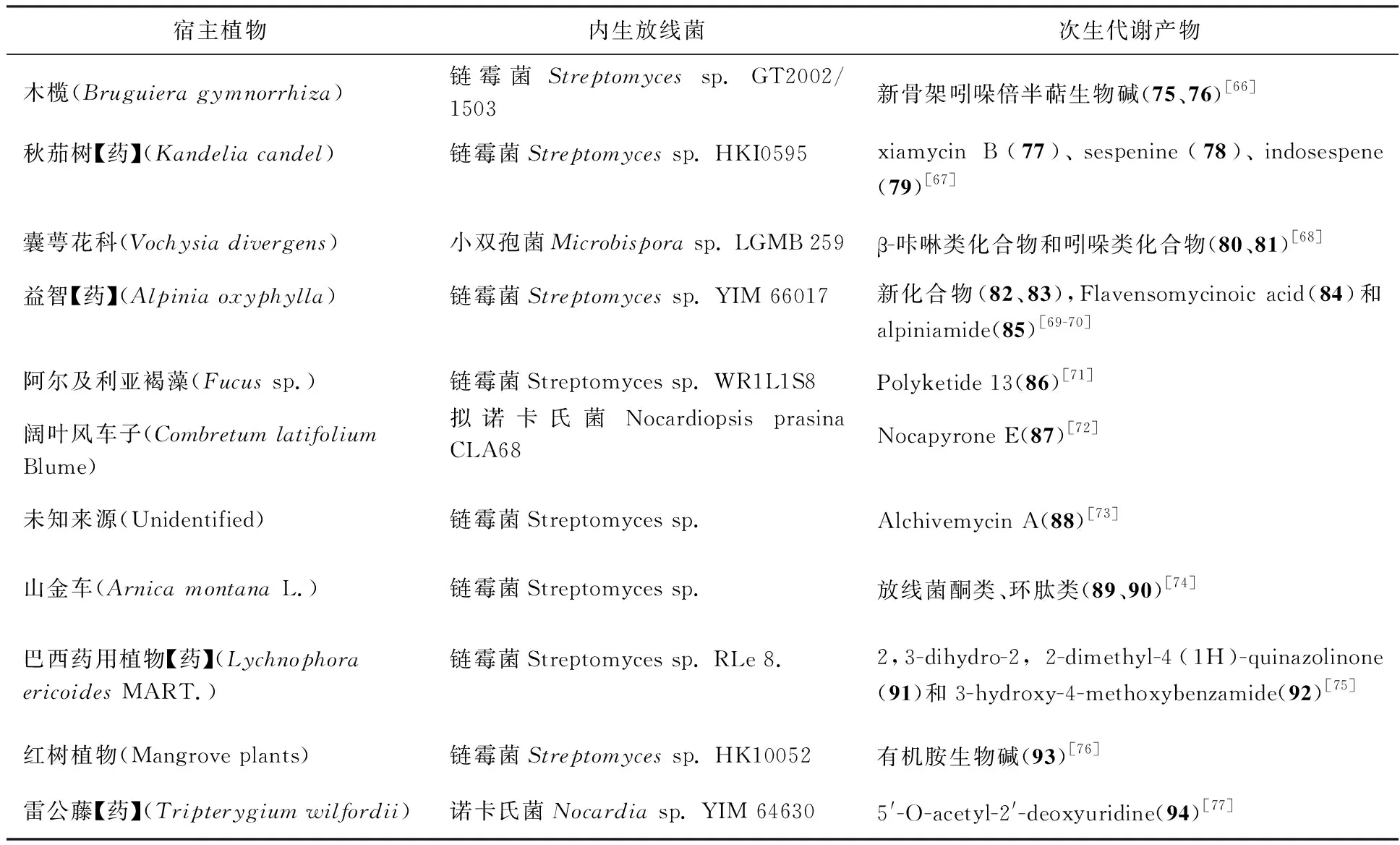
宿主植物内生放线菌次生代谢产物木榄(Bruguiera gymnorrhiza)链霉菌Streptomyces sp. GT2002/1503新骨架吲哚倍半萜生物碱(75、76)[66]秋茄树【药】(Kandelia candel)链霉菌Streptomyces sp. HKI0595xiamycin B(77)、sespenine(78)、indosespene(79)[67]囊萼花科(Vochysia divergens)小双孢菌Microbispora sp. LGMB 259β-咔啉类化合物和吲哚类化合物(80、81)[68]益智【药】(Alpinia oxyphylla)链霉菌Streptomyces sp. YIM 66017新化合物(82、83),Flavensomycinoic acid(84)和alpiniamide(85)[69-70]阿尔及利亚褐藻(Fucus sp.)链霉菌Streptomyces sp. WR1L1S8Polyketide 13(86)[71]阔叶风车子(Combretum latifoliumBlume)拟诺卡氏菌Nocardiopsis prasina CLA68Nocapyrone E(87)[72]未知来源(Unidentified)链霉菌Streptomyces sp.Alchivemycin A(88)[73]山金车(Arnica montana L.)链霉菌Streptomyces sp.放线菌酮类、环肽类(89、90)[74]巴西药用植物【药】(Lychnophora ericoides MART.)链霉菌Streptomyces sp. RLe 8.2,3-dihydro-2, 2-dimethyl-4(1H)-quinazolinone(91)和3-hydroxy-4-methoxybenzamide(92)[75]红树植物(Mangrove plants)链霉菌Streptomyces sp. HK10052有机胺生物碱(93)[76]雷公藤【药】(Tripterygium wilfordii)诺卡氏菌Nocardia sp. YIM 646305'-O-acetyl-2'-deoxyuridine(94)[77]
2 植物内生菌次生代谢产物的生物活性
大量研究表明,无论是植物内生菌发酵产物的乙酸乙酯提取物还是甲醇提取物,均显示出一定的生物学活性,从中得到的一些单体化合物同时也显示出生物学活性。通过比较发现,近年来发现的活性物质中萜类及生物碱类占大部分,其主要表现出抗菌和抗肿瘤活性;醌类和酮类(包括放线菌酮、苯并吡喃酮、α-吡喃酮、黄酮、苝醌、蒽醌等)表现出抗菌、杀虫活性;内酯类和脂肪酸酯类则表现出酶抑制活性;此外,还有次生代谢产物具有抗病毒、抗氧化等生物活性。近年来得到的植物内生菌次生代谢产物的具体生物活性见表4~6。

表4 植物内生菌代谢产物的抗菌活性

表5 植物内生菌代谢产物的细胞毒活性

表6 植物内生菌代谢产物的其他活性
3 展 望
青霉素和链霉素作为微生物代谢产物在人类与感染性疾病的抗争中使用了很多年,但抗生素的不合理使用使得病原菌出现耐药性的时间间隔越来越短,同时生活水平的不断提高所带来的新疾病也严重威胁着人类的健康、生存和发展。植物内生菌尤其是药用植物内生菌作为重要的微生物资源,能产生结构多样的活性物质,具有多种重要生物学功能。从植物中分离内生菌并从中挖掘生物活性成分,尤其抗临床耐药菌如MRSA等的活性成分,是阻止人类退回到青霉素发现以前感染性疾病肆虐时代的有效措施,同时也为解决传统的天然产物药源——药用植物生长缓慢、资源紧缺等问题提供了新的启发。
虽然植物内生菌次生代谢产物的研究已经成为寻找结构新颖活性化合物的一个重要途径,但就目前的研究来看,大部分集中在菌株的分离及活性菌株的筛选,活性物质的追踪挖掘较少,得到活性物质后的作用机制及扩大应用则更少,植物内生菌的研究在许多方面仍有待加强:①在植物选材上不能盲目跟风,应选择新颖或具有一定特色的植物,如道地中药材和濒危灭绝的植物。②目标菌株的筛选相对简单,大都是常见的抗菌和抗肿瘤活性及常见的化学结构类型筛选,应建立一些快速、稳定、针对性强如抗耐药菌等的菌株筛选平台,同时进行发酵条件和生产工艺的优化,以服务生产为导向。③内生菌的研究真菌多放线菌少,放线菌是抗生素的主要来源,应进一步探索植物尤其是药用植物内生放线菌的分离培养方法,为活性代谢产物的深入挖掘提供更有效的菌种资源。
