长沙藤蕹开花的光周期特性研究
2018-07-24朴立喜辛戴思慧
朴立喜 刘 娟 艾 辛戴思慧
(湖南农业大学园艺园林学院,湖南长沙 410128)
藤蕹为旋花科甘薯属蕹菜(Ipomoea aquatic Forsk)中不易开花或不能开花的品种类型,是华南、华东、西南及华中地区夏季主栽绿叶蔬菜之一(周淑荣 等,2014)。藤蕹一般不能开花,个别年份虽然能在生长晚期形成花蕾,但不能开花结实,故生产中只能采用成熟的茎秆窖藏越冬留种(谈敏等,2017)。由于藤蕹种茎窖藏技术复杂,环境不易控制,容易出现缺种、失种的情况。一般采用大棚越冬留种(殷武平 等,2017)、南繁早春漂浮留种(童辉 等,2017)和种苗组培快繁技术(杨博智 等,2014)等进行藤蕹种苗繁殖,提高种苗繁殖效率。但是,长期采用无性繁殖,容易导致藤蕹种性退化严重;同时,由于不能开花结实,也限制了藤蕹种质创新和新品种的选育。有性杂交育种是藤蕹种质创新和新品种选育的最佳途径,通过有性繁殖能够实现品种间、类型间优良性状基因的有效重组和聚合。开展藤蕹杂交育种的前提是诱导藤蕹开花,初步研究表明藤蕹为严格的短日照植物(龚明辉 等,2015),但有关藤蕹的光周期特性却鲜见报道。短日照植物只有在日照长度小于临界日照长度的条件下进行光周期诱导才能成花(付建新,2014),但能否诱导成花还与植株大小、处理时间、光质、温度等多种因素有关。光周期对植物开花调控的有效起始时间取决于植物幼龄期结束的时间,而终止时间则由限界性诱导光周期来确定(胡惠蓉 等,2007)。光周期诱导时间小于限界性光周期,已分化的花芽会发生逆转,在紫苏(Lam &Leopold,1961)、大豆(Liu et al.,2009)中表现明显。本试验以不开花的长沙藤蕹为试验材料,探讨藤蕹开花的光周期诱导特性,确定藤蕹的临界光周期、幼龄期以及限界性光周期,建立藤蕹开花诱导技术,为今后开展藤蕹种质创新和新品种选育奠定理论基础。
1 材料与方法
1.1 试验材料
试验于2017~2018年在湖南农业大学园艺园林学院人工气候室进行。试验材料为采集于湖南农业大学耘园基地的长沙藤蕹,是长沙地区种植历史悠久的地方优良品种,在长沙地区不能进行花芽分化和开花结实。藤蕹的繁殖采用茎段扦插法,将老熟的藤蕹植株剪成2个茎节的茎段,扦插于装满基质(园土∶草炭=2 V∶1 V)的直径为25 cm的塑料盆中,放置在FYS-10型智能人工气候室培养,培养条件为光照强度270 μmol·m-2·s-1、昼/夜温度30 ℃/26 ℃、光照时间15 h·d-1、空气湿度70%;每隔2 d浇灌1次有机肥水。
1.2 藤蕹光周期处理方法
1.2.1 藤蕹临界光周期的测定 将生长在光照时间15 h·d-1条件下,长势整齐一致的藤蕹植株(8片真叶,平均株高30 cm)分别放置在10 L∶14 D、11 L∶13 D、12 L∶12 D、13 L∶11 D、14 L∶10 D的光周期条件下进行处理,观察记录植株现蕾开花情况,以全部植株出现花芽和开花为确定光周期长度的依据。每处理3次重复,每重复10株。
1.2.2 藤蕹限界性光周期的测定 依据临界光周期的测定结果,将11 h·d-1作为藤蕹限界性光周期测定的光周期长度。将生长在15 h·d-1条件下、长势整齐一致的藤蕹植株(8片真叶,平均株高30 cm)分别放置在光照时间11 h·d-1条件下处理1、4、7、10、13、16、19、22、25 d后,重新将植株迁移至15 h·d-1条件下培养,观察记录植株现蕾和开花情况。每处理3次重复,每重复10株。
1.2.3 藤蕹幼龄期的测定 分别将生长在长光照(15 h·d-1)条件下具有1、2、3、4、5、6、7、8片叶的藤蕹植株迁移至短光照周期11 h·d-1生长4 d和7 d后,重新迁移至长光照(15 h·d-1)条件下培养,观察记录植株现蕾开花节位、时间。同时,选取在长光照周期15 h·d-1生长的叶数(茎节数)为0、2、4、6、8、10、12、14片的藤蕹植株,迁移至短光照周期11 h·d-1条件下培养直至开花,观察记录植株现蕾及开花节位和时间。每处理3次重复,每重复10株。
1.2.4 藤蕹光周期暗夜光中断试验 挑选长势整齐一致的藤蕹植株(8片真叶,平均株高30 cm)分别进行4组处理:第1组,11 h·d-1光照处理;第2组,11 h·d-1光照并在暗夜时进行强闪光处理;第3组,13 h·d-1光照处理;第4组,13 h光照/13 h黑暗处理。处理10 d后,观察记录植株现蕾开花情况。每处理3次重复,每重复10株。
2 结果与分析
2.1 藤蕹临界光周期特性
从表1可以看出,藤蕹在光照时间≤12 h·d-1能现蕾开花,光照时间>12 h·d-1不能现蕾开花,其临界光周期为12 h光照/12 h黑暗;在12 h·d-1的光照条件下现蕾、开花所需要时间较长,与10、11 h·d-1处理的现蕾、开花时间差异显著,而10 h·d-1和11 h·d-1处理的现蕾、开花所需时间均较短且差异不显著。综合考虑植株的生长和现蕾开花时间,诱导藤蕹开花的最佳光周期为11 h·d-1。

表1 不同光周期处理对藤蕹现蕾、开花时间的影响
2.2 藤蕹限界性光周期特性
从表2可以看出,短日照处理4 d以上的藤蕹植株现蕾率在53.3%以上,但短日照处理天数≤7 d植株分化的花蕾黄化死亡,不能成花;当短日照处理10 d时,20.0%的植株可以开花;短日照处理天数达到16 d时,植株的开花率达到100.0%。由此确定藤蕹的限界性光周期为16 d。
2.3 藤蕹幼龄期特性
由图1可见,4片叶以上的藤蕹植株能感受短光照的诱导产生花蕾,但4片叶时现蕾率低于50%;植株越大现蕾率越高,8片叶植株处理4 d的现蕾率达到88.89%,处理7 d现蕾率达到100.00%。但由于处理时间没有达到藤蕹限界性光周期,所有的花蕾都不能发育成花。
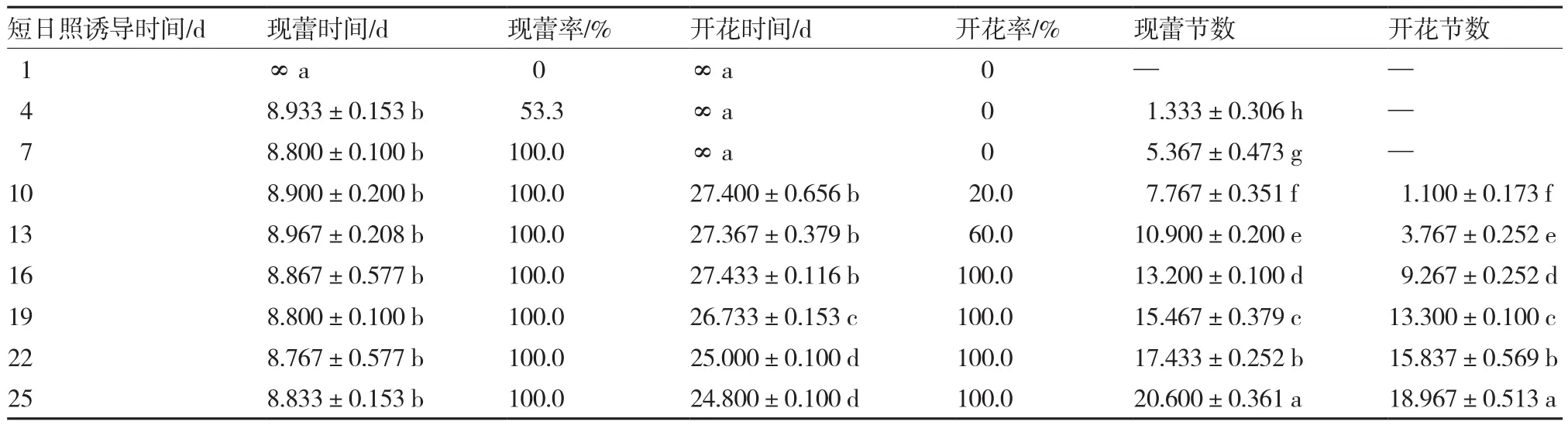
表2 不同短日照诱导时间对藤蕹现蕾、开花的影响
从表3可以看出,不同大小的藤蕹植株在11 h光照/13 h黑暗条件下培养直至开花,现蕾时间、开花时间以及现蕾起始节位均具有显著性差异,但现蕾率无差异。随着叶片数的增加,现蕾、开花时间逐渐减少,8片叶以后现蕾时间差异不显著;现蕾起始节位在4片叶以前趋于稳定,4片叶以后显著增加。由于藤蕹是在每个节位上开花,因此衡量藤蕹幼龄期的标准是最小现蕾起始节位的最短时间所对应的叶片数目。
综上,藤蕹植株在4片真叶时幼龄期结束。
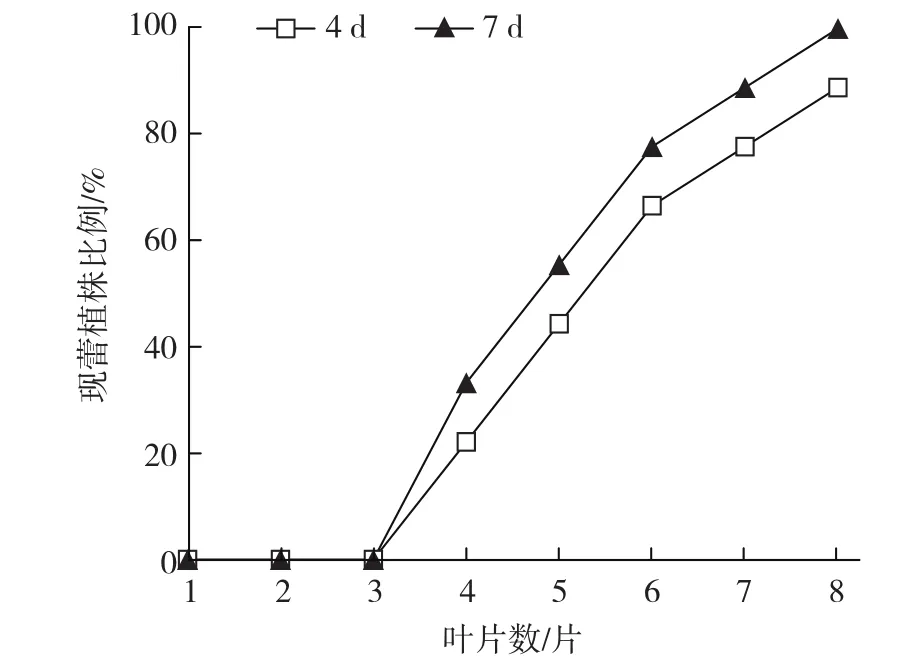
图1 植株大小和短日照诱导天数对藤蕹现蕾的影响
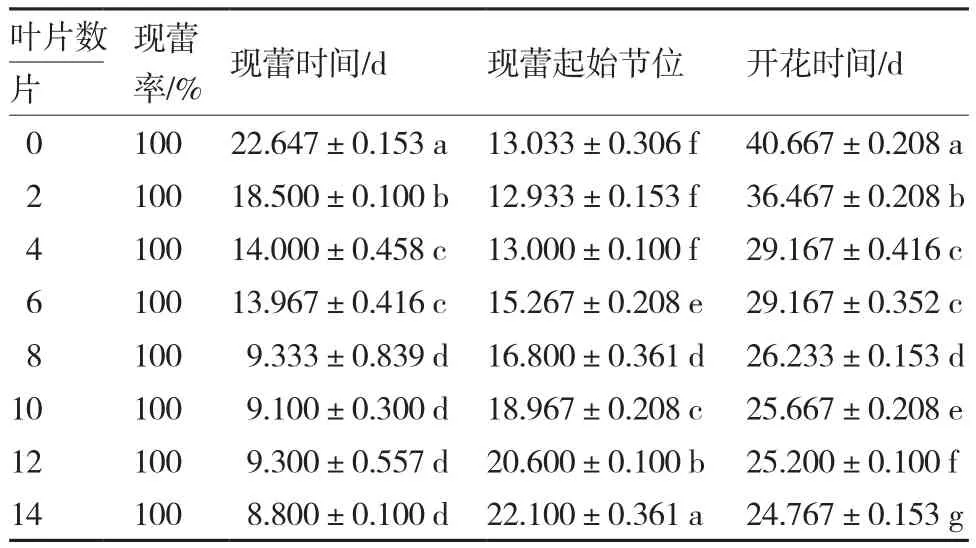
表3 短日照条件下植株大小对藤蕹现蕾、开花的影响

图2 不同光照处理对藤蕹开花的影响
2.4 光周期暗夜光中断对藤蕹开花的影响
长日植物的开花诱导过程主要取决于日照长度,而短日植物的开花诱导过程主要取决于暗期长度,只要达到暗期长度则可现蕾开花。从图2可以看出,当暗夜长度为11 h,藤蕹不能成花;当暗夜长度为13 h,即使延长光照时间,藤蕹也可以成花,但如果在暗夜期间进行强闪光处理则不能成花。说明藤蕹为短日植物,暗期长度是藤蕹成花的关键。
3 结论与讨论
植物对光周期的响应特性分为两种类型,一种对光周期的响应是兼性或量性的,即诱导条件可以促进开花,但非诱导条件下也能开花;另一种对光周期的响应属于专性或质性的,即只能在诱导条件下才能开花(Evenari & Gutterman,1966)。本试验结果表明,在光照时间≤12 h·d-1的光周期条件下藤蕹都可以现蕾开花,而光照时间>12 h不能诱导藤蕹现蕾开花,说明藤蕹的光周期临界日长为12 h光照/12 h黑暗,属于专性短日植物或质性短日植物;同时还发现,藤蕹在暗期长度为13 h时可以快速响应短日照诱导,植株生长健壮,确定诱导藤蕹开花的最佳光照时间为11 h·d-1。
研究表明,植物在适合的光周期下诱导一定时间后终止光周期诱导,仍然可以启动花芽分化,花芽能继续发育并最终完成开花,但也有些植物如短日植物茼蒿(Chrysanthemum morifolium)在15 d短日诱导之后,移入长日条件下,花序的花芽发生了逆转不能成花;如果延长短日照处理天数则分化的花芽不会被逆转(Battey & Lyndon,1990)。本试验发现,藤蕹在短日照诱导少于16 d时,所分化的花芽能被长日照逆转,花芽停止发育,黄化死亡,这种现象在大豆(李晓梅 等,2005)和裂叶矮牵牛(King & Evans,1969)上也同样存在,说明这些植物诱导成花需要最低限的光周期诱导周期数(常以天为单位),称为限界性诱导光周期(Damann,1993)。根据环境因素、成花决定的程度与成花逆转的关系,植物成花逆转有3种类型,第1种是成花决定一旦启动就不会发生逆转;第2种是成花决定在开花的任何时候都能发生逆转;第3种是成花决定完成之前可发生逆转,成花决定完成后不能逆转(白素兰 等,2000)。藤蕹的花芽分化不在其生长点的顶端,每个叶腋着生1个花芽或花序,花芽分化随着植株生长持续进行,其成花节位数与短日照光周期诱导时间呈正相关,从低节位至高节位顺序开花。藤蕹花芽成花与成花逆转的第3种情况相似,在完成成花决定之前可逆转,但花芽逆转的方式与其他植物不同,花芽不会转化为营养生长,而是黄化死亡。藤蕹在花芽分化完成后,需要持续的短光周期处理,才能完成成花决定,否则已分化的花芽会停止发育,说明藤蕹的花芽分化和成花是可以通过光周期调控进行逆转的。长沙藤蕹的限界性光周期为16 d,但处理16 d时开花数较少,不足10朵。为了保证有足够多的节位开花结实,建议将藤蕹置于持续的短日照下20 d以上,甚至更长时间,这也增加了藤蕹诱导开花的成本。
本试验通过对诱导藤蕹开花的光周期特性进行研究,确定了诱导藤蕹启动花芽分化的临界光周期、诱导藤蕹开花的最佳光周期、限界性光周期,确定了4叶期以上的植株才能感受短日照光周期诱导。通过调控光周期,成功地解决了藤蕹有性繁殖的难题,为开展藤蕹种质创新和遗传育种奠定了理论和技术基础。诱导藤蕹开花既可以在设施条件良好的人工气候室进行,也可以考虑冬季在海南三亚南繁,三亚11月下旬至翌年1月中旬(大约60 d)的光照时间少于12 h,很接近长沙藤蕹的临界光周期,理论上长沙藤蕹在三亚是可以开花的,但缺少实证,能否在海南三亚顺利开花还有待进一步试验。
