无核香味葡萄胚挽救育种研究
2018-07-20史文静骆强伟王跃进
史文静,骆强伟,王跃进*
(1 西北农林科技大学 园艺学院,陕西杨陵 712100;2 旱区作物逆境生物学国家重点实验室,陕西杨陵 712100;3 农业部西北地区园艺作物生物学与种质创制重点实验室,陕西杨陵 712100;4 新疆维吾尔自治区葡萄瓜果研究所,新疆鄯善 838200)
目前,市场上大部分的无核品种,如‘底莱特’、‘波尔莱特’、‘红宝石无核’等是通过常规杂交选育而来[1]。常规杂交育种一般以有核葡萄品种为母本,无核性状遗传能力强的葡萄品种为父本,通过杂交选育无核新种质。然而,该方法获得的后代无核比率较低[2-4]。1982年,Ramming等[5]首次以无核葡萄为母本利用胚挽救技术获得了无核后代。相较于传统育种,胚挽救技术子代无核率高、育种周期短,因而已被广泛应用于葡萄无核种质创制中[6-12]。研究表明,葡萄中的芳香物质属于萜烯类化合物,其遗传与表达受到至少5对优势互补基因和1个修饰基因调控[13-14]。已有研究结果表明,相较有香味亲本,杂种F1代玫瑰香表型呈现明显减弱趋势,即浓香型与中等、淡香味或者无香味的品种杂交,其杂交果实多数无香味;浓香型品种与浓香型品种的杂交后代中,果实玫瑰香味的比例较大,且后代变异系数较小,组合遗传力较大[15]。综上,利用胚挽救技术培育香味无核新种质具有极高可行性。
前期研究发现,从胚珠中分离的合子胚可在培养基中继续离体发育直至成熟,这一观点也成为运用胚挽救技术选育新品种的重要理论支撑[16]。胚挽救技术涉及以下3个主要过程:(1)胚珠离体培养,使杂种胚在胚珠中得以继续发育成熟;(2)在无菌条件下剥取杂种幼胚置于萌发培养基萌发成苗;(3)胚挽救杂种苗炼苗移栽[17]。然而,目前胚挽救技术中仍面临着胚发育率低、成苗率低等问题[18-21]。大量研究表明葡萄胚挽救效率受到父母本基因型、采样时间、培养基、培养条件等因素影响[17]。其中,无核品种前期的胚珠与胚挽救过程中幼胚的发育都严格受到其父母本基因型影响[6-7, 17, 22-23]。Cain等[23]通过将13个无核品种部分进行自交,发现不同的父母本间的胚发育率和成苗率差异较大。另外,一些研究表明相比于‘无核白’和‘波尔莱特’,‘火焰无核’更适合作为母本进行胚挽救育种[24-25]。胚挽救过程中的采样时间同样作为影响胚挽救效率的重要因素[17]。自然状态下,假单性结实无核葡萄中只有少数合子胚发育成熟,大部分合子胚逐渐败育直至消失[19]。采样时间影响胚囊中杂种胚的发育程度,进而影响胚挽救结果[26]。除此之外,培养基作为胚挽救过程中离体培养胚珠和胚的媒介,其培养基类型、基础培养基种类、培养基中添加物等对胚挽救效率有着重要影响[17]。研究胚珠离体培养过程中发现,高浓度糖可以促进幼胚发育[6, 9, 12]。同时,一些研究表明某些添加物如植物生长调节剂、氨基酸等对于胚发育具有促进作用[11, 26-29]。
葡萄育种中利用胚挽救技术已近四十年,但目前市场上通过胚挽救技术释放的新品种较少[17]。因此,利用胚挽救技术选育优良葡萄新品种仍是目前育种工作者不断探索和研究的重点。
1 材料和方法
1.1 植物材料
本研究于2016 年5 月~2017 年5 月在新疆维吾尔自治区葡萄瓜果研究所与西北农林科技大学农业部西北地区园艺作物生物学与种质创制重点试验室完成。供试材料中,母本选择7个无核品种,分别是‘火焰无核’、‘昆香无核’、‘红宝石无核’、‘莫莉莎无核’、‘克瑞森无核’、‘奇妙无核’和‘波尔莱特’;父本材料为4个香味葡萄品种,‘贵妃玫瑰’、‘玫瑰香’、‘巨玫瑰’和‘昆香无核’。
1.2 方 法
1.2.1胚挽救流程1) 田间杂交授粉。本研究中围绕香味无核的育种目标,在当年母本植株成花情况基础上,结合本课题组前期研究结果,配置了9个杂交组合(表1)。根据杂交组合配置,收集父本花粉,研磨晾干后待用。参照当年物候,开花前2~3 d(2016年5月9日~12日),对杂交组合中母本进行去雄处理。去雄后的花穗,立即套袋以防止外来花粉污染,并挂牌记录去雄日期。去雄后的3 d,柱头前端出现透明黏液,即可在晴天上午10:00以前对去雄花穗进行人工授粉,连续授粉3 d,记录授粉时间。
2)大田采样。参考实验室前期研究,对杂交组合进行连续取样,以确定不同亲本最佳采样时间。其中,对以‘火焰无核’、‘昆香无核’、‘红宝石无核’、‘莫莉莎无核’、‘克瑞森无核’、‘奇妙无核’和‘波尔莱特’为母本的组合采样时间分别为授粉后39~45 d、49~55 d、54~60 d、53~57 d、39~42 d、48~50 d和51~57 d。
3)胚珠离体培养与发育。取回的杂交果实,统计果粒数后,置于流水下冲洗2~3 h。在超净工作台中,将冲洗后的果实装进高压蒸汽灭菌过的丝口瓶中,按照以下顺序消毒处理:70%酒精消毒30 s后,2%次氯酸钠消毒20 min,然后用无菌水冲洗幼果3~5次,期间不断摇晃瓶身以确保消毒充分。将消毒完成的果粒置于无菌的培养皿中,用手术刀及镊子小心剥取胚珠,接种于改良MM3培养基(MM3+60 g/L蔗糖+ 0.5 g/L水解酪蛋白+ 0.1 g/L肌醇+3 g/L活性炭)中,暗培养8周。每个培养皿接种15个胚珠。
4)离体幼胚萌发成苗。在无菌条件下,利用显微镜将经过离体发育的胚从胚珠中剥离后,接种于加入胚萌发培养基的摇菌瓶中(WPM+0.2 mg/L BA+0.1 g/L肌醇+7 g/L琼脂+1 g/L活性炭+20 g/L蔗糖),置于培养温度为(25±2)℃,光周期为12 h /12 h,光照强度2 000 lx的组培间进行萌发成苗。
5)炼苗移栽。将经过继代扩繁的杂种苗,选择其中生长茁壮、根系发达的壮苗进行炼苗移栽。具体为:将基质土与蛭石按照5∶1的比例混匀后装袋高压灭菌后,晾凉备用。小心取出杂种苗,洗干净培养基后植于装有4/5处理过基质的小花盆中,并罩上透明杯,标注移栽日期及组合类型。逐步掀开透明杯至彻底去掉,期间每2 d浇1次水,直至炼苗完成。在气温适宜的晴天将其移植至大田,浇透水,统计成活率。
6)分子标记辅助选择育种。CTAB 法提取杂交子代株系的葡萄基因组DNA,应用分子标记GSLP1-569[30]和SCF27-2000[31]对其进行无核标记检测。
1.2.2离体胚珠培养时间以杂交组合‘红宝石无核’ב克瑞森无核’的胚珠为材料,研究不同胚珠培养时间对后期胚挽救效率影响。实验中选择3个胚珠置于MM3暗培养的时间(64 d、79 d和85 d),统计5种不同发育时期胚(A 球形期胚;B 心形期胚;C 鱼雷形期胚;D 子叶形期胚;E 畸形胚)比率,并记录其萌发过程。从而从胚发育角度解释培养时间对胚挽救结果的影响。试验数据来源于3次重复试验,通过 SPSS Statistics 22软件进行方差分析和显著性检测(P<0.05)。
2 结果与分析
2.1 不同亲本对胚挽救效率的影响
在大田杂交后,对不同杂交组合的杂种果实分期分批取样,其胚挽救结果如表1所示。共培养香味无核杂交胚珠6 733个,胚珠经过离体培养8周后,剥取获得胚832个,胚发育率为12.36%;获得正常发育的胚挽救幼苗365 株,成苗率为5.42%;对杂交后代中长势良好的234株幼苗进行炼苗后移栽大田,182株杂种胚挽救苗成活(图1)。图1为通过胚挽救育种获得无核香味葡萄新种质的整个操作流程。
通过比较, ‘红宝石无核’ב玫瑰香’和‘莫莉莎无核’ב玫瑰香’2个组合胚发育率最高,分别为24.03%和17.19%;组合‘波尔莱特’ב玫瑰香’、‘昆香无核’ב玫瑰香’和‘昆香无核’ב巨玫瑰’胚发育率次之,分别为14.33%、13.04%和11.31%; ‘克瑞森无核’ב巨玫瑰’和‘火焰无核’ב贵妃玫瑰’胚发育率最低,分别是1.84%和3.83%。组合‘红宝石无核’ב玫瑰香’成苗率最高,为14.63%; ‘波尔莱特’ב玫瑰香’、‘昆香无核’ב玫瑰香’和‘昆香无核’ב巨玫瑰’成苗率次之,分别为7.74%、5.64%和3.09%;组合‘莫莉莎无核’ב玫瑰香’、‘火焰无核’ב贵妃玫瑰’、 ‘火焰无核’ב巨玫瑰’成苗率较低,分别为1.56%、1.03%和0.58%;‘克瑞森无核’ב巨玫瑰’、‘奇妙无核’ב昆香无核’成苗率都为0(表1)。
2.2 采样时间对不同亲本胚挽救效率的影响
采样时间影响胚发育程度,因此以胚发育率作为主要判断最佳采样时间的因素,正常苗率及畸形苗率辅助说明结果。表1中,以‘火焰无核’为母本的2个组合(‘火焰无核’ב巨玫瑰’和‘火焰无核’ב贵妃玫瑰’)在授粉后的43 d时采样,胚发育率和成苗率较高。其中,‘火焰无核’ב贵妃玫瑰’胚发育率最高,为8.54%;组合‘火焰无核’ב巨玫瑰’胚发育率为6.96%。以‘昆香无核’为母本的两个组合(‘昆香无核’ב玫瑰香’和‘昆香无核’ב巨玫瑰’)在授粉后的51~52 d期间采样较为合适。其中,组合‘昆香无核’ב玫瑰香’授粉后51 d采样条件下胚发育率和成苗率最高,分别为18.13%和8.75%;组合‘昆香无核’ב巨玫瑰’在授粉后51 d采样条件下胚发育率最高,为16.92%。组合‘红宝石无核’ב玫瑰香’授粉后59~60 d采样条件下,胚挽救效果最佳,胚发育率分别为34.21%和32.77%,此时成苗率也较高(17.37%和20.85%)。‘莫莉莎无核’ב玫瑰香’授粉后54 d采样最佳,胚发育率和成苗率分别为16.67%和10%。组合‘克瑞森无核’ב巨玫瑰’、‘奇妙无核’ב昆香无核’分别在授粉后39 d和48 d之前采样胚挽救结果较好。而在组合‘波尔莱特’ב玫瑰香’中,采样时间控制在授粉后52 d,可获得最佳胚挽救结果(胚发育率18.53%、成苗率11.85%)。


a. 大田杂交: a-1. 母本人工去雄,a-2. 父本花粉采集(箭头指示去雄后柱头);b. 人工授粉;c. 采样;d. 消毒后将胚珠接种至改良MM3培养基(箭头指示离体剥取胚珠);e. 从胚珠中无菌剥取胚接种于胚萌发培养基 (WPM+ 0.2 mg/L BA+0.1 g/L肌醇+7 g/L琼脂+1 g/L 活性炭+20 g/L蔗糖);f. 胚萌发;g. 成苗;h. 人工气候室中炼苗;i. 次年5月移栽至大田图1 葡萄胚挽救技术选育无核葡萄新种质基本流程图a. Treatments of the parents: a-1. Artificial emasculation; a-2. pollen collection, Arrow: a larger version of pistil after emasculation; b. Artificial pollination; c. Sampling; d. Inoculation into MM3 medium after sterilization, Arrow: a larger version of embryo excised from ovule in vitro; e. Sub-culture embryo stripped into medium (WPM supplemented with 0.2 mg·L BA, 0.1 g/L inositol, 7 g/L agar, 1 g/L AC, and 20 g/L sucrose) from ovule for embryo germination; f. Embryo germination; g. Developing into seedling; h. Domestication in phytotron; i. Transplanting into the field in May of the second yearFig.1 General procedure of breeding seedless grape (‘Ruby Seedless’ × ‘Muscat’) by embryo rescue
2.3 胚珠离体培养时间对胚挽救结果的影响
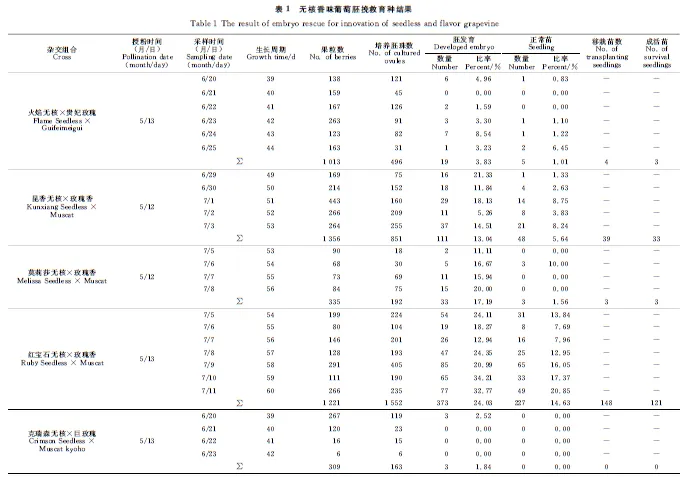
不同类型胚比率随胚珠培养时间变化结果显示(图2,Ⅰ),子叶形胚所占比率最高,为52.94%~64.52%,其次为鱼雷形胚。且胚发育时期不随胚珠培养时间延长而发生变化,然而随着胚珠在改良MM3培养基上暗培养时间的延长,畸形胚比率显著提高。
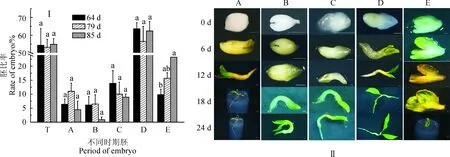
对于不同发育时期胚萌发成苗的进一步观察结果表明(表2),子叶形胚萌发率及成苗率都显著高于其他发育时期胚,分别为93.04%~97.78%、49.16%~58.73%。同时,畸形胚更易萌发为畸形苗,其畸形苗率为47.22%~66.67%。对于不同发育时期胚的在接种到萌发培养基24 d内的萌发情况观察结果显示(图2,Ⅱ),子叶形胚萌发成苗最快,在接种后的12 d即萌发成为正常苗,除畸形胚外其余发育时期胚也在24 d内发育成正常植株。
Ⅰ. 胚珠离体培养时间对胚发育时期的影响;标准误(SE)显示在柱型图上;不同小写字母表示各处理间0.05水平差异显著性。Ⅱ. 不同发育时期胚离体萌发成苗情况;标尺=200 μm;T. 总发育胚;A 球形期胚;B 心形期胚;C 鱼雷形期胚;D 子叶形期胚;E 畸形胚图2 胚珠离体培养时间对组合‘红宝石无核’ב克瑞森无核’胚挽救效率影响Ⅰ. Comparison of the proportion of embryo in vitro at different periods with the extension of ovule culture time. Means with different letters are significantly different (P<0.05). Ⅱ. Different stages of embryo germination and seedling in medium in vitro; Scale bars = 200 μm; T. Total; A. Globe-shape; B. Heart-shape; C. Torpedo-shape; D. Cotyledon-shape; E. MalformationFig.2 Effect of culture time in vitro of ovules from the cross Ruby Seedless × Crimson Seedless on embryo rescue
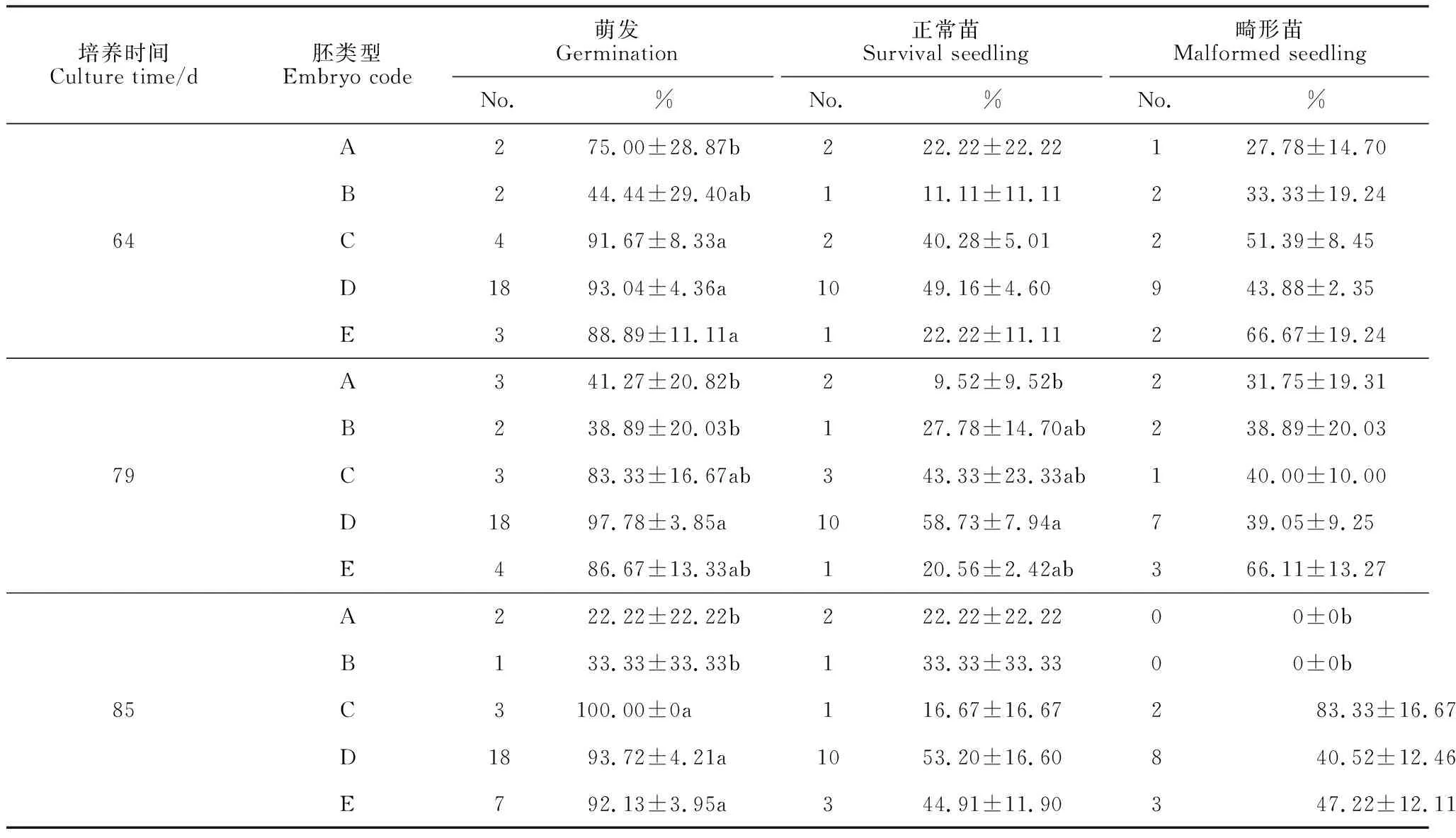
表2 不同发育时期胚的萌发成苗
注:百分率相对于各个培养时间内不同发育时期所对应的胚发育数计算得到;萌发、畸形苗、正常苗率表示为平均值±SE;同列不同小写字母表示各处理间0.05水平差异显著性Note: Percent was calculated relative to the number of inoculated embryos of the corresponding types respectively. Data represent mean±SE. Means with different letters are significantly different for each culture time (P<0.05)

A. 无核探针SCF27-2000对杂交亲本的检测;B. 无核标记GSLP1-569对杂交亲本的检测。M:2k plus Marker;1.昆香无核;2.巨玫瑰;3.玫瑰香;4.红宝石无核;5.波尔莱特;6.火焰无核;7.贵妃玫瑰;8.莫莉莎无核图3 无核标记(探针) 对杂交亲本的检测A. Schematic the detection of parents with grapevine seedless gene using GSLP1-569 probe; B. Schematic the detection of parents with grapevine seedless gene using marker SCF27-2000. M. 2k plus Marker; 1. Kunxiang Seedless; 2. Muscat kyoho; 3. Muscat; 4. Ruby Seedless; 5. Perlette; 6. Flame Seedless; 7. Guifeimeigui; 8. Melissa SeedlessFig.3 Molecular marker-assisted selection of parents with grapevine seedless gene
2.4 胚挽救杂交子代无核性状的早期分子标记检测结果
2.4.1分子标记筛选不同组合亲本利用目前已有无核标记对杂交组合中双亲无核性状进行早期检测,结果如下:(1)图3,A为无核标记基因SCF27-2000对8个杂交亲本的扩增结果。从图中可以看出,5个母本‘火焰无核’、‘昆香无核’、‘红宝石无核’、‘莫莉莎无核’和‘波尔莱特’都扩增出了2 000 bp 特异性条带。除‘昆香无核’以外其余3个父本‘贵妃玫瑰’、‘玫瑰香’、‘巨玫瑰’均未在2 000 bp 处出现特异性条带。因此用SCF27-2000 无核标记对3个杂交组合‘昆香无核’ב玫瑰香’、‘昆香无核’ב巨玫瑰’和‘红宝石无核’ב玫瑰香’的胚挽救杂交后代进行无核性状检测;(2)无核探针GSLP1-569对8 个杂交亲本扩增结果如图3,B所示,母本‘火焰无核’扩增出569 bp特异条带,而在其他无核亲本中并没有扩增出569 bp 特异条带,因此以‘火焰无核’为母本的杂交组合(‘火焰无核’ב巨玫瑰’和‘火焰无核’ב贵妃玫瑰’)可以选择无核基因探针GSLP1-569 对其杂交后代进行无核性状的早期鉴定。
2.4.2无核标记(或探针)对杂交子代的检测1)无核标记SCF27-2000对杂交子代的检测。无核标记SCF27-2000 对杂交组合‘昆香无核’ב玫瑰香’、‘昆香无核’ב巨玫瑰’和‘红宝石无核’ב玫瑰香’杂交子代的60 个株系进行无核性状的检测。结果如图4,A、B,其中组合‘昆香无核’ב玫瑰香’的15个株系中有11个株系在2 000 bp处扩增出特异性条带;‘昆香无核’ב巨玫瑰’15个株系中有13个株系具有2 000 bp 特异性条带;组合‘红宝石无核’ב玫瑰香’30个株系中有14个株系在2 000 bp 处出现特异性条带。
2)无核探针GSLP1-569对杂交子代的检测(图4,C)。无核探针GSLP1-569 对杂交组合‘火焰无核’ב巨玫瑰’和‘火焰无核’ב贵妃玫瑰’的12个株系进行无核检测,其中‘火焰无核’ב巨玫瑰’7株系,有4个株系具有569 bp 特异性条带;‘火焰无核’ב贵妃玫瑰’5个株系中,4个株系扩增出569 bp的特异性条带。
3 讨 论
胚挽救育种过程中,亲本基因多样性是导致表型、胚珠及胚发育、成苗等方面差异的首要因素。1982年以来,国内外育种工作者不断尝试各种无核或有核品种以期获得更高的胚挽救效率,从而得到较多胚挽救杂种后代[17]。胚挽救结果受母本基因型影响较多,同时父本也对胚挽救效率有一定影响[17]。Emershad等[4]在研究中以‘无核白’为亲本,成苗率仅为10%~15%。前期研究中,以‘底莱特’为母本的杂交组合胚挽救效率较高,其胚发育率高于母本材料为‘红脸无核’的组合[19]。以 ‘火焰无核’、‘爱神玫瑰’、‘无核白鸡心’和‘红宝石无核’为母本,父本为‘双优’、 ‘北醇’的杂交组合成苗率相对较高[18, 25]。本文试验中,同样培养条件下,母本为‘红宝石无核’、‘火焰无核’、‘昆香无核’的杂交组合胚挽救效果较好,其中‘红宝石无核’为母本的组合成苗较高(14.63%),是胚挽救中很好的母本材料,这与前人结果一致[32-33]。香味品种中‘玫瑰香’更适合作为父本材料来选育香味无核种质。
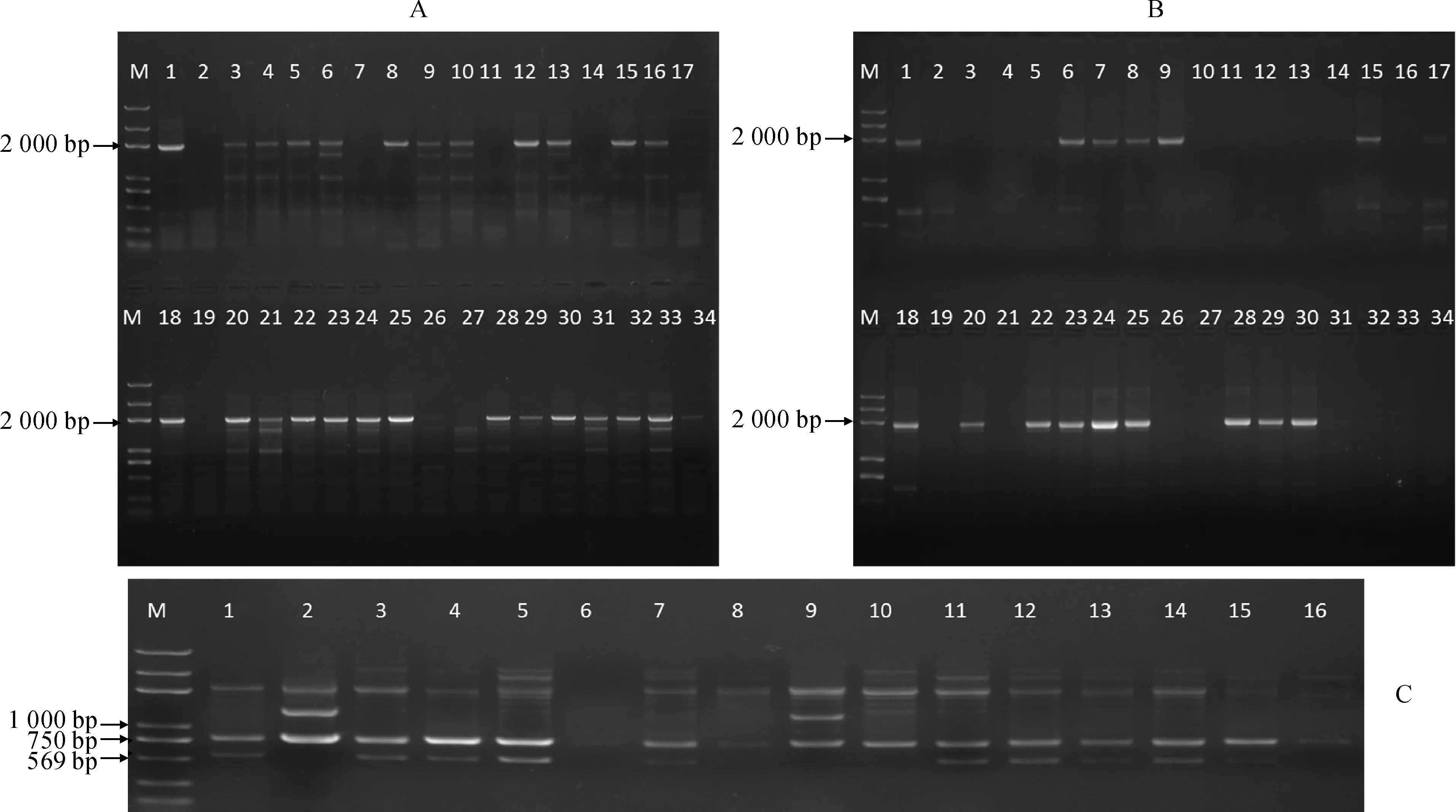
M. 2k plus Marker;A. 无核标记SCF27-2000 检测组合‘昆香无核’ב玫瑰香’和‘昆香无核’ב巨玫瑰’子代无核性状: 1和18. 昆香无核;2. 玫瑰香;3~17. 杂交组合‘昆香无核’ב玫瑰香’不同杂交株系;19. 巨玫瑰;20~34. 杂交组合‘昆香无核’ב巨玫瑰’不同杂交株系。B. 无核标记SCF27-2000 检测组合‘红宝石无核’ב玫瑰香’子代无核性状:1和18. 红宝石无核;2和19. 玫瑰香;3~17和20~34. 杂交组合‘昆香无核’ב玫瑰香’不同杂交株系。C. 无核探针GSLP1-569检测组合‘火焰无核’ב巨玫瑰’和‘火焰无核’ב贵妃玫瑰子代无核性状:1和10. 火焰无核;2. 巨玫瑰;3~9. 杂交组合‘火焰无核’ב巨玫瑰’不同杂交株系;11. 贵妃玫瑰;12~16. 杂交组合‘火焰无核’ב贵妃玫瑰’不同杂交株系图4 利用无核标记SCF27-2000和无核基因探针GSLP1-569鉴定胚挽救杂交苗无核性状M. 2k plus Marker; A. Schematic the detection of hybrid progeny seedlings with grapevine seedless gene using marker SCF27-2000: 1 and 18. Kunxiang Seedless; 2. Muscat; 3-17. hybrid progeny seedlings of the cross Kunxiang Seedless × Muscat; 18. Kunxiang Seedless; 19. Muscat kyoho; 20-34. hybrid progeny seedlings of the cross Kunxiang Seedless × Muscat kyoho. B. Schematic the detection of hybrid progeny seedlings with grapevine seedless gene using marker SCF27-2000: 1 and 18. Ruby Seedless; 2 and 19. Muscat; 3-17 and 20-34. hybrid progeny seedlings of the cross Ruby Seedless × Muscat. C. Schematic the detection of hybrid progeny seedlings with grapevine seedless gene using GSLP1-569 probe: 1 and 10. Flame Seedless; 2. Muscat kyoho; 3-9. hybrid progeny seedlings of the cross Flame Seedless × Muscat kyoho; 11. Guifeimeigui; 12-17. hybrid progeny seedlings of the cross Flame Seedless × GuifeimeiguiFig.4 Molecular marker-assisted selection of hybrid progeny seedlings with grapevine seedless gene
采样时间同样对胚挽救结果有着重要的影响,采样过早胚发育不成熟,而采样过晚胚已经败育,最终导致胚挽救成苗率低[26]。而判断采样时间是否合适的标准则是胚发育率,从胚珠中剥取幼胚的数量即胚发育率[11]。李桂荣等[18]2010年胚挽救结果表明,‘火焰无核’、‘克瑞森无核’、‘红宝石无核’的最佳采样时期分别为授粉后45 d、60 d和65 d。本研究在2015年以‘火焰无核’、‘克瑞森无核’和‘红宝石无核’为母本的杂交组合分别在授粉后的43 d、39 d前、59~60 d采样,胚发育率和成苗率较高。本试验结果与之前研究有所差异,可能是由于不同年份、不同地区物候不同引起胚发育差异。试验中相同母本最佳采样时间相同或相近,表明母本材料对采样时间的确定影响较大,而父本基因型对其也有一定影响[19]。
通过胚挽救获得的植株只能在其结实后确定是否为杂种后代,而胚挽救过程中利用分子标记辅助可更早对后代表型进行早期判断,是生物技术与育种的完美结合。Mejia和王跃进等[30-31]分别开发出无核标记SCF27-2000和无核探针GSLP1-569,为分子标记筛选、检测无核性状奠定了基础。然而利用无核标记SCF27-2000 检测杂交子代时,若存在等位基因,检测结果会受到影响,并且不能很好区分亲本的有核、软核、无核性状[19]。前期研究发现无核探针GSLP1-569检测有些无核亲本如‘爱莫无核’和‘红宝石无核’时,在569 bp处未出现特异性条带[20, 34]。本试验选择无核探针GSLP1-569和无核标记SCF27-2000 对亲本和子代无核性状进行检测。亲本中只有‘火焰无核’在569 bp处出现特异条带,因此无核探针GSLP1-569可对以‘火焰无核’为亲本的组合杂交后代进行无核性状的早期鉴定;无核标记SCF27-2000检测亲本无核性时,5 个亲本(‘火焰无核’‘昆香无核’‘红宝石无核’‘莫莉莎无核’‘克瑞森无核’和‘波尔莱特’)扩增出2 000 bp 特异性条带,可用于对3个组合‘昆香无核’ב玫瑰香’、‘昆香无核’ב巨玫瑰’和‘红宝石无核’ב玫瑰香’的杂交后代的检测。检测的72个杂交株系中46个株系有特异条带,初步确定为无核株系。标记检测过的幼苗,炼苗移栽后加速杂种苗的生长发育,促进杂种苗尽快进入结果期以观察其性状表现。
综上,本研究在利用胚挽救技术创制无核香味葡萄新种质的同时,对影响葡萄胚挽救效率的亲本基因型、取样时间、胚珠培养时间与胚发育时期等进行研究,为后期葡萄胚挽救育种提供参考。
