水芹肉桂醇脱氢酶基因的克隆与表达特性分析
2018-07-19朱胜琪王永鑫刘洁霞熊爱生
仇 亮,朱胜琪,王永鑫,冯 凯,刘洁霞,熊爱生
(南京农业大学 园艺学院,作物遗传与种质创新国家重点实验室,农业部华东地区园艺作物生物学与种质创制重点实验室,南京 210095)
水芹[Oenanthejavanica(Blume) DC.]为伞形科水芹属多年生草本植物,常见于长江中下游地区,是重要的水生蔬菜[1]。水芹既可食用又具有药用价值[2]。除富含人体所需的营养元素外,水芹还富含抗坏血酸、维生素、膳食纤维、芹菜素、黄酮类化合物等多种成分,对人体具有很好的保健功能,可以降压安神、增强人体对外在环境的免疫能力、防治心脑血管等疾病[3]。
木质素是植物中肉桂醇脱氢聚合产生的酚类高分子聚合物,为沉积在高等植物特定细胞壁中第二丰富的天然化合物,是植物细胞壁的重要组成成分[4]。木质素的疏水特性使细胞壁不易透水,可以保证水分的长距离运输,提高植株的物质运输能力[5]。木质素的生物合成过程可以总结为苯丙氨酸或酪氨酸在一系列酶的催化下,逐步转化为木质素单体,最终聚合成木质素的过程[6]。肉桂醇脱氢酶(cinnamyl alcohol dehydrogenase, CAD)参与木质素特异合成途径下游酶类的调控,是一类重要的限速酶[7]。木质素作为膳食纤维的重要组成部分,能改善人体健康状况,降低一些疾病的风险,但含量增加后,植株细胞壁会加厚,植物组织变得木质化,很大程度上会影响蔬菜口感。作为以幼嫩叶和茎为主要食用部位的水芹,木质素含量对其品质有较大的影响[8]。
目前,相对于模式植物和大田作物而言,水芹的分子生物学研究相对滞后。本试验选用‘八卦洲水芹’及其紫色叶柄突变型‘南选八卦洲紫水芹’为材料,从水芹中克隆得到1个编码肉桂醇脱氢酶的基因OjCAD,对其进行了生物信息学分析,并利用荧光定量PCR技术分析了OjCAD基因在‘八卦洲水芹’和‘南选八卦洲紫水芹’叶片和叶柄中的表达特性,为进一步研究水芹木质素合成途径及OjCAD响应机制提供了实验基础。
1 材料和方法
1.1 材料及处理
本实验以‘八卦洲水芹’和紫色叶柄突变型水芹‘南选八卦洲紫水芹’为植物材料[9],于2017年2月下旬种植于南京农业大学作物遗传与种质创新国家重点实验室人工气候室,生长条件为:25 ℃光照12 h/20 ℃黑暗12 h,光照强度300 μmol·m-2·s-1。在水芹生长至抽薹后取样,样品用液氮速冻后保存于-80 ℃冰箱,用于总RNA的提取。大肠杆菌(Escherichiacoli)菌株DH5α保存于本实验室,pMD19-T质粒载体、ExTaqDNA聚合酶、Prime Script RT reagent Kit试剂盒等为大连TaKaRa公司产品。
1.2 方 法
1.2.1总RNA提取及cDNA合成将所取的水芹样品利用RNA Simple Total RNA Kit总RNA提取试剂盒提取总RNA(北京Tiangen公司),并按照 Prime Script RT reagent Kit试剂盒说明书提取水芹总RNA,然后反转录成cDNA。
1.2.2水芹肉桂醇脱氢酶基因的克隆根据本课题组的水芹转录组数据[10],检索得到水芹OjCAD基因,并对OjCAD序列进行克隆分析。利用Primer Premier 6软件设计1对克隆引物,OjCAD-F(ATGACGCAAACAACTCCCAA)和OjCAD-R(CTATGGTTTTCCAGCAATGT)。以水芹cDNA为模板进行PCR扩增。扩增体系总体积10.0 mL,包括0.30 ng·mL-1cDNA模板0.5 mL、2 × TaKaRaExTaq Mix (产品编号RR001A) 5.0 mL、双蒸水3.5 mL、0.10 mmol·L-1正向引物和反向引物各0.5 mL。扩增的反应条件为:94 ℃预变性5 min;94 ℃变性30 s,54 ℃退火30 s,72 ℃延伸80 s,共34个循环;最后在72 ℃延伸10 min。将反应所得产物用12 g·L-1琼脂糖凝胶电泳分离并回收。回收产物连接到pMD19-T载体后转化到大肠杆菌DH5α,菌液检测后送至南京金斯瑞生物科技有限公司测序鉴定。
1.2.3序列分析通过DNAMAN 6.0软件确定基因的开放阅读框(ORF)序列,翻译成相应的氨基酸序列后进行氨基酸序列的亲水性和疏水性分析。利用 NCBI数据库中的Blastp进行同源比对和保守域预测,采用MEGA 5.0软件绘制系统进化树[11]。不同物种CAD蛋白的氨基酸组成及理化性质分析均在ExPASy (http://www.expasy.org)上完成[12]。利用CPHmodels 3.2 Server (http://www.cbs.dtu.dk/services/CPHmodels/) 进行空间建模分析。
1.2.4基因表达特性分析根据OjCAD基因测序结果,使用Real-time PCR检测系统(Bio-rad, CFX96, USA)在96孔板中进行qPCR反应。以水芹actin基因作为荧光定量数据处理的内参基因[13],利用Primer Premier 6软件设计荧光定量PCR上游引物(5′-CTGGATTCTGGTGATGGT-3′)和下游引物(5′-GCAGTTGTTGTGAAGGAG-3′)。根据扩增基因的序列设计表达检测上游引物(5′-CAGGCACAATCACTCCTTACG-3′)和下游引物(5′-TCACACTTCAAACACGACGC-3′)。反应采用20 mL体系,每个PCR反应物体系中包含2.0 mL稀释15倍的cDNA,0.4 mL上下游荧光定量引物,10 mL SYBR Green I mix和7.2 mL ddH2O。PCR反应条件如下:95 ℃预变性30 s,95 ℃变性5 s,60 ℃ 退火延伸30 s,共进行40个循环。每个PCR反应重复3次,分析3个重复。使用Excel软件进行不同组织内表达水平的分析,CAD基因在水芹不同品种叶片和叶柄中相对表达量的计算采用2-ΔΔCT方法[14]。
2 结果与分析
2.1 水芹OjCAD基因的克隆
提取正常生长的水芹植株叶的RNA,并反转录成cDNA,然后以此为模板,利用OjCAD-F和OjCAD-R引物进行PCR扩增,获得1 000 bp左右的扩增片段。测序结果显示,水芹的OjCAD基因包含一个长度为1 074 bp开放阅读框(ORF),编码357个氨基酸(图1)。其中,碱基A、T、G、C的总量分别为305、276、272和221个,分别占总碱基数的28%、26%、25%和21%。
2.2 水芹OjCAD蛋白的进化分析
利用 MEGA 5.0 软件绘制CAD蛋白在不同物种间的进化树(图2)。结果显示,水芹OjCAD蛋白与伞形科的胡萝卜(Daucuscarota)CAD蛋白进化关系最近,其次是地星(Dorcocerashygrometricum);水芹的OjCAD蛋白与芝麻(Sesamumindicum)、甜樱桃(Prunusavium)和藜麦(Chenopodiumquinoa)CAD蛋白进化关系较远。水芹的OjCAD蛋白与同为伞形科的胡萝卜在同一分支上,表明CAD蛋白的氨基酸序列在同科植物的进化中比较保守。

*. 终止密码子图1 水芹OjCAD基因核苷酸序列及其编码的氨基酸序列*. Stop codonFig.1 The nucleotide and putative amino acid sequences of OjCAD gene from O. javanica

括号内为氨基酸登录号;其他物种来源的CAD氨基酸序列皆来源于NCBI数据库;标尺表示遗传距离图2 水芹与其他物种CAD蛋白的系统进化树The accession No. was given in brackets; The amino acid sequences of CAD proteins from other species were derived from NCBI database; The scale bar represents genetic distanceFig.2 Phylogenetic tree of CAD protein from O. javanica and other species
2.3 不同物种CAD蛋白氨基酸序列比对与氨基酸理化性质分析
利用NCBI对水芹CAD蛋白序列进行Blastp检索,结果表明,克隆得出的水芹CAD蛋白具有一个典型的CAD1结构域,此外还包含CAD特征性的2个Zn结合位点(图3),与CAD酶学功能相符,属于MDR超级家族。选取来源于巴西橡胶树、菠菜、地星、藜麦、榴莲、葡萄、甜樱桃、胡萝卜、长春花、芝麻的CAD蛋白,与水芹OjCAD蛋白进行氨基酸序列的多重比对(图4)。结果显示,水芹OjCAD蛋白序列与已被证实的另外10个物种的氨基酸序列一致性达到了85.75%。其中,水芹OjCAD蛋白序列与胡萝卜CAD蛋白序列的一致性达到了95.24%,可见,水芹与同为伞形科的胡萝卜亲缘进化关系极近。进一步通过水芹和其他物种CAD氨基酸组成及理化性质比较(表1)可以发现,这 11种植物CAD蛋白氨基酸数在357~366之间, 相对分子质量均在3.9×104左右, 理论等电点在6.42~7.57之间,碱性氨基酸与酸性氨基酸数目相差不大, 脂肪族氨基酸比例较高, 芳香族较低, 蛋白质可溶性预测中平均亲水性在-0.164~-0.074之间。
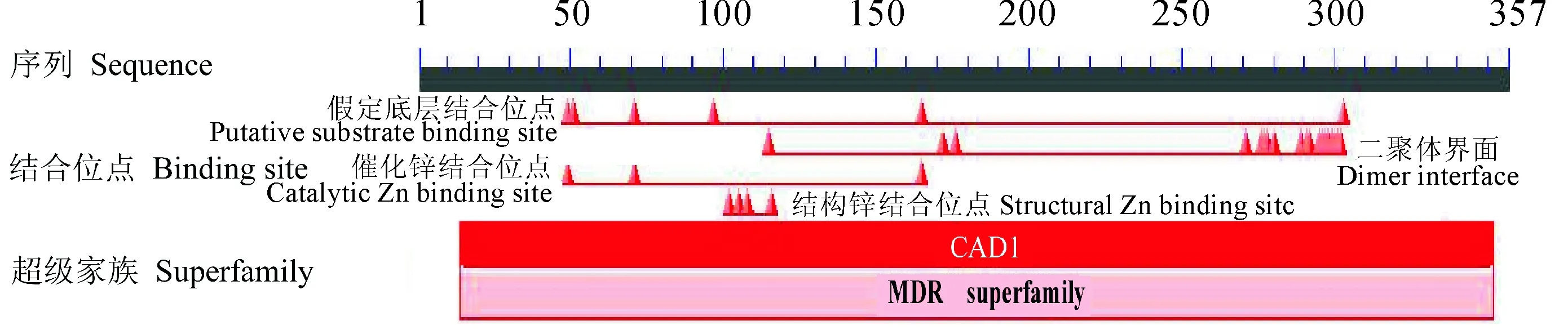
图3 水芹OjCAD氨基酸序列保守域预测Fig.3 Prediction of conserved domain of amino acid sequence of OjCAD from O. javanica
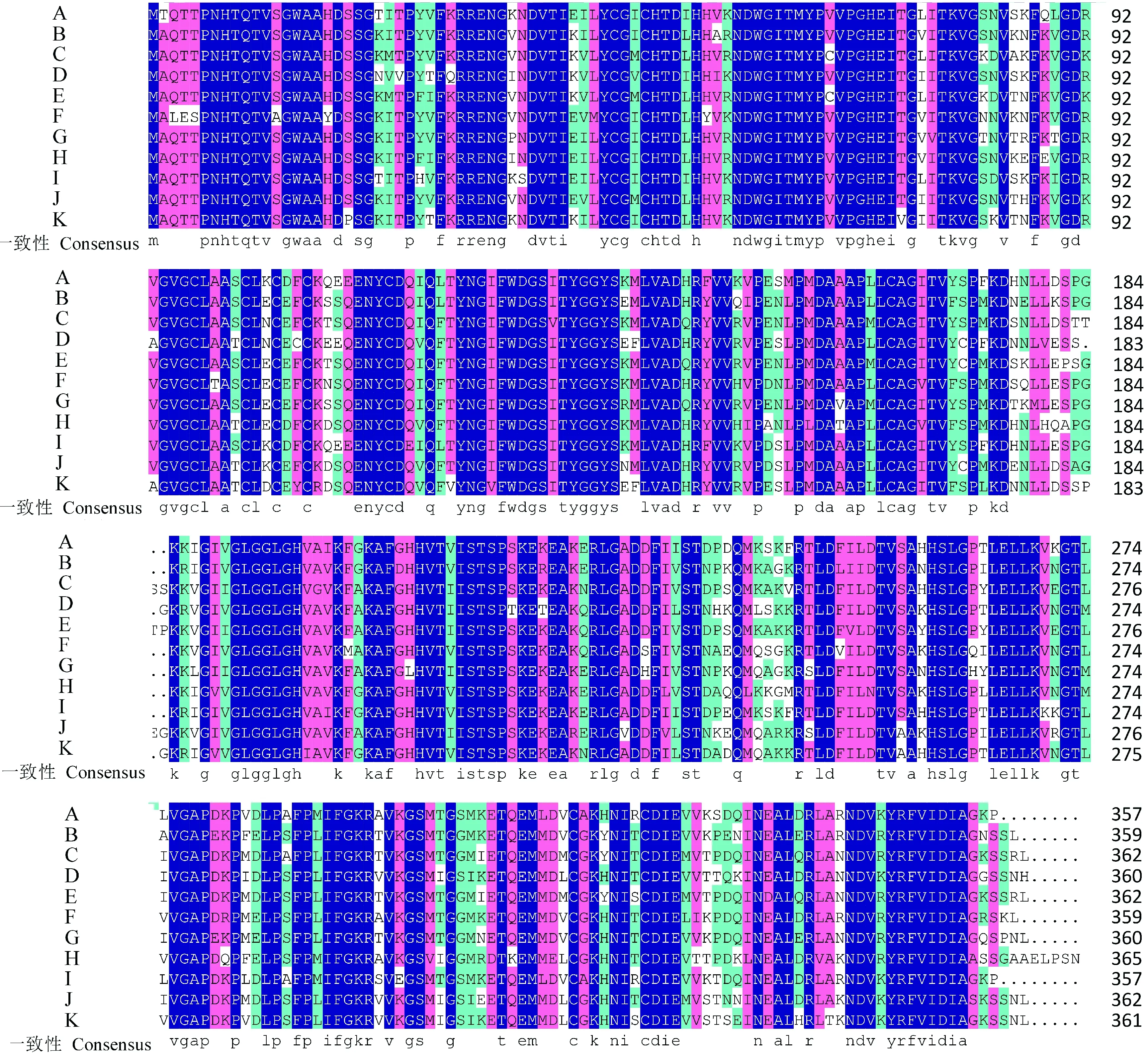
A. 水芹; B. 巴西橡胶树; C. 菠菜; D. 地星; E. 藜麦; F. 榴莲; G. 葡萄; H. 甜樱桃; I. 胡萝卜; J. 长春花; K. 芝麻;表1同。图4 水芹与其他物种CAD氨基酸序列的多重比对A. O. javanica; B. Hevea brasiliensis; C. Spinacia oleracea; D. Dorcoceras hygrometricum; E. Chenopodium quinoa; F. Durio zibethinus; G. Vitis vinifera; H. Prunus avium; I. Daucus carota; J. Catharanthus roseus; K. Sesamum indicum;The same as Table 1Fig.4 Multiple alignment of amino acid sequences of CAD from O. javanica and other species
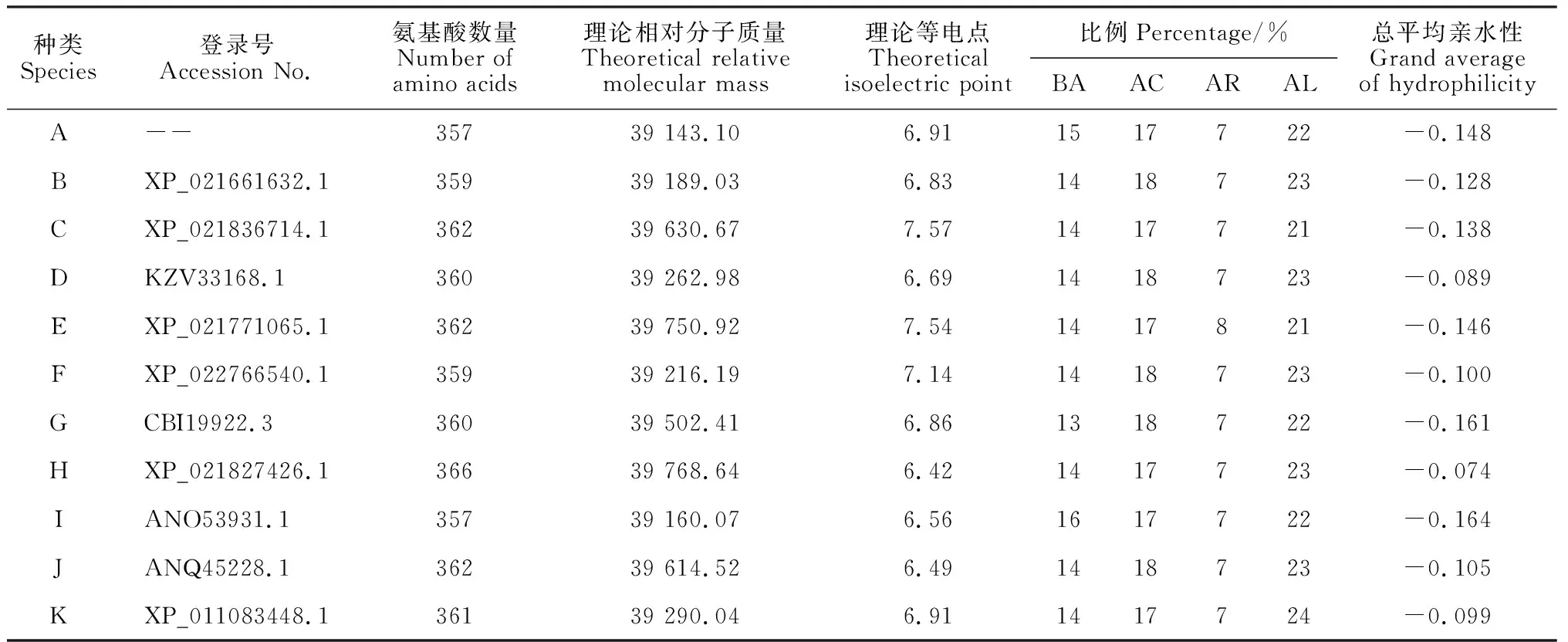
表1 水芹及其他物种CAD氨基酸组成及理化性质比较
注:BA. 碱性氨基酸; AC. 酸性氨基酸; AR. 芳香族氨基酸; AL. 脂肪族氨基酸Note:BA. Basic amino acids; AC. Acidic amino acids; AR. Aromatic amino acids; AL. Aliphatic amino acids

图5 水芹OjCAD蛋白氨基酸序列的疏水性和亲水性分析Fig.5 Analysis of hydrophobicity and hydrophilicity of amino acid sequence of OjCAD protein from O. javanica
2.4 水芹OjCAD氨基酸亲疏水性分析
利用DNAMAN 6.0软件进行蛋白亲水性疏水性分析,结果显示,水芹OjCAD氨基酸位于第189位的异亮氨酸疏水性最强,位于第221的赖氨酸和223位的丙氨酸亲水性最强。从总体来看,水芹OjCAD属于疏水性蛋白(图5)。

图6 水芹OjCAD蛋白的三级结构Fig.6 Tertiary structure of OjCAD protein from O. javanica
2.5 水芹OjCAD蛋白的三级结构预测及分析
通过CPHmodels 3.2 Server对水芹OjCAD蛋白的三级结构进行了预测及分析,以PDB ID: 1YQD.A为模型对OjCAD蛋白序列进行同源建模,其一致性为58.7%。结果显示,水芹OjCAD蛋白结构主要由7个α-螺旋和17个β-折叠组成(图6)。
2.6 OjCAD基因在紫色和非紫色水芹叶片和叶柄中的表达分析
利用qRT-PCR方法对‘八卦洲水芹’和‘南选八卦洲紫水芹’的叶片和叶柄中OjCAD基因的相对表达量进行分析。如图7所示,OjCAD基因在紫色和非紫色水芹叶片和叶柄中的表达量有明显的差异。其中,OjCAD基因在‘八卦洲水芹’中的相对表达量是‘南选八卦洲紫水芹’的2倍;OjCAD基因在叶片中的表达量相对较高,且品种差异显著,而在叶柄中的表达量极低,且品种间无显著差异。
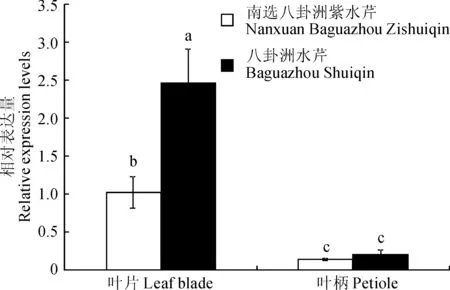
图7 OjCAD基因在水芹叶片与叶柄中的相对表达量Fig.7 The relative expression levels of OjCAD gene in the leaf blades and petioles of O. javanica
3 讨 论
在高等植物中,木质素是细胞壁的基本组成成分,对植物具有形态塑成、机械强度支撑等重要作用[15]。在蔬菜中,木质素常与纤维素、半纤维素互相连接,形成天然屏,阻止各种病原菌的侵入,增强植物应对干旱、病害等环境胁迫的能力,对植物抗病虫和抗逆性等有很大的影响[16]。适宜的木质素含量对于蔬菜作物的品质很重要,尤其是叶菜类蔬菜。有研究表明,木质素合成的增多会导致鸭儿芹食用器官木质化,严重影响鸭儿芹的品质[17]。水芹是中国重要的水生蔬菜,研究水芹木质素生物合成,能更有效提高水芹的品质,改善水芹的口感。
高等植物中,肉桂醇脱氢酶(CAD)作用于木质素单体合成的最后一步,在植物木质素生物合成中起着重要的作用。在黑麦草[18]、陆地棉[19]、高粱[20]等植物中都已获得相关基因。在木质素合成过程中,肉桂醇脱氢酶以NADPH为辅酶将肉桂醛还原成相应的肉桂醇,合成不同的木质素单体,是调控木质素合成的关键控制点之一[21]。有研究证实,在拟南芥中将CAD4和CAD5这2个基因双突变之后,拟南芥茎部柔软,有倒茎现象,整个植株呈现倒伏状。经测定,其与野生型拟南芥相比,植株中的维管组织含量降低,木质素含量下降了94%[22]。石榴种皮中的木质素含量是影响石榴籽粒硬度的重要因子[23],石榴肉桂醇脱氢酶基因PgCAD的表达量与种皮总木质素含量呈正相关,PgCAD基因在石榴籽粒中表达量最高,肉桂醇脱氢酶对石榴籽粒的硬度起重要作用[24]。
水芹除了维生素和蛋白质等含量很高外,还含有芸香苷、水芹素和檞皮素等物质。为保证其鲜嫩的口感,适当降低叶和茎中的木质素含量至关重要。本研究以本课题组水芹转录组数据为基础,从水芹中克隆获得1个编码肉桂醇脱氢酶的OjCAD基因。OjCAD氨基酸序列含有典型的CAD1结构域,且具有特征性2个Zn结合位点[25]。系统进化分析显示,水芹OjCAD与同科植物胡萝卜CAD具有最近的亲缘关系。OjCAD氨基酸序列属于MDR超级家族[26]。水芹与同一科植物CAD蛋白氨基酸理化性质相近,同科植物的CAD蛋白同源性也较高。另外,有研究表明,在叶柄中,木质素只能在维管束、厚角组织和表皮细胞中可以观察到[27]。本研究荧光定量结果显示,OjCAD在水芹叶片中的表达量高于叶柄,推测OjCAD基因在水芹木质素合成中起重要作用。本研究结果为水芹木质素的合成与调控等研究奠定了理论基础。
