马铃薯冠气温差变化特性与耐旱性的关系
2018-07-13杨宏羽刘玉汇白江平峰王张俊莲
余 斌 杨宏羽 王 丽 刘玉汇 白江平 张 峰王 蒂 张俊莲,*
1甘肃农业大学农学院 / 甘肃省遗传改良与种质创新重点实验室 / 甘肃省干旱生境作物学国家重点实验室培育基地, 甘肃兰州730070; 2甘肃农业大学生命科学技术学院, 甘肃兰州 730070
马铃薯(Solanum tuberosum)是世界第三大粮食作物[1]。中国是世界第一大马铃薯生产国, 马铃薯在中国各个生态区域均有种植, 尤其在西部地区种植面积更大[2]。中国西部属于典型的气候变化敏感区, 近 50年来气温显著上升, 降水明显减少, 干燥指数显著增加, 干旱灾害事件发生频率提高[3]。研究表明未来50年全球气候将继续变暖,湿润半湿润地区暖干化趋势明显, 对粮食安全产生重大影响[4]。选育适宜在干旱及半干旱区种植的耐旱马铃薯品种, 可以有效解决干旱恶化问题。
干旱条件下, 植物本身会通过一系列生理反应, 以维持体内正常生理代谢所需要的水分, 从而保护自身不受干旱伤害, 这种途径是植物耐旱的主要方式之一, 这些生理反应可作为植物耐旱性的评定指标, 其中叶片冠层温度、光合效率以及蒸腾速率均是抗旱鉴定的重要指标, 并有研究表明, 三者之间具有密切的关联性。Lourtie等[5]和Laury等[6]研究发现, 植物冠层温度在正常条件下通过蒸腾作用保持稳定, 在干旱条件下植物气孔关闭, 蒸腾强度减弱, 改变了冠层热量损失程度, 导致冠层温度变化, 而这些变化均会引起叶片光合效率的改变。植物冠层温度会受到空气温度的直接影响, 因此, 利用植物冠层温度与周围空气温度的差值即冠气温差作为植物耐旱性指标的研究已多见报道。Amani等[7]和 Fan等[8]研究发现, 遭受干旱胁迫时, 小麦冠气温差能够显著反映小麦的耐旱性, 冠气温差可以作为小麦耐旱基因型筛选的一个重要指标。研究人员也将冠气温差作为高效水分利用型小麦及耐旱型珍珠粟的有效筛选工具, 其中 Singh等[9]通过比较不同基因型珍珠粟的冠气温差对其进行了准确的耐旱性分类。便携式红外成像仪的出现使得冠气温差指标更加广泛的应用于植物干旱胁迫响应研究以及耐旱基因型评价工作中。此外, 干旱对蒸腾速率和光合作用也会产生重要影响, 蒸腾速率变化是作物抵御干旱的一种重要生理反应, 光合作用是作物生长的物质基础。Jefferies[10]以干旱和非干旱条件下的马铃薯研究发现, 干旱导致马铃薯叶片气孔导度下降、蒸腾速率降低, 从而限制了光合作用。Basu等[11]研究发现, 水分胁迫下马铃薯净光合速率明显下降。
目前, 对马铃薯冠气温差在不同气候条件下及不同基因型之间的变化特征研究较少, 冠气温差与马铃薯耐旱性的关系仍不明确, 本研究以从国际马铃薯中心引进的耐旱型和干旱敏感型马铃薯种质为材料, 以耐旱型和干旱敏感型马铃薯品种为对照, 在半干旱与半湿润2种气候类型环境中, 对不同基因型马铃薯的耐旱性及冠气温差变化关系, 以及马铃薯冠气温差与蒸腾速率、光合效率及生物量积累间的关系进行研究, 探讨利用冠气温差评价马铃薯耐旱性的可行性, 为马铃薯耐旱育种技术提供理论依据。
1 材料与方法
1.1 试验材料与田间种植方法
供试材料由14份抗旱能力各不相同的四倍体马铃薯种质材料组成, 其中有来自于秘鲁国际马铃薯中心(International Potato Center, CIP)的种质材料10份, 西北旱作区大面积种植的耐旱型品种冀张薯8号、陇薯6号,以及干旱敏感型品种大西洋(Atlantic)和夏波蒂(Shepody)。

表1 试验品种和CIP马铃薯资源编号Table 1 Experimental varieties and code of potato germplasm resources from CIP
2015—2016年连续2年在 2种不同干湿类型地区甘肃省定西市鲁家沟镇(地理坐标: 104°35'14" E, 35°51'37" N,海拔1850 m, 年平均降雨量220 mm, 年平均蒸发量1620 mm, 属中温带半干旱区, 年平均气温 8.6℃, 无霜期 146 d)和甘肃省定西市五竹镇(地理坐标: 104°08'02"E,35°03'32"N, 海拔2182 m, 年平均降雨量650 mm, 年平均蒸发量 820 mm, 属中温带半湿润区, 年平均气温 4.7℃,无霜期130 d)进行田间试验。按小区种植, 每小区2垄, 单垄两行, 每行10株, 株距为0.3 m, 垄距0.8 m, 每小区间隔1.0 m的走廊, 设 3次重复。在两地均施氮肥 210 kg hm–2, N∶P2O5∶K2O 比例为 1.4∶1.0∶2.0, 分别用 15∶15∶15的复合肥、尿素和硫酸钾。采用机械化一次性基施, 不追肥, 人工点播后覆土。参照《马铃薯实验研究方法》[12]测定株高、叶面积、叶鲜重指标。
1.2 试验区平均降水量和气温状况
2015—2016年鲁家沟镇年平均降雨量258.8 mm, 年平均气温9.8℃, 五竹镇年平均降雨量604.2 mm, 年平均气温 5.6℃, 这两年两地试验区的平均降雨量和气温与历史水平值接近, 无特殊气候现象发生(图1)。
1.3 光合指标测定
使用光合仪(LI-COR, LI-6400XTP, USA)测定净光合速率Pn、叶片气孔导度Gs、蒸腾速率Tr。从每小区选择3株能够反映该品种植株表型形态的具有代表性的健康植株, 从播种 40 d后(苗期)开始, 选择晴天 9:00—12:00测定, 以自然日光为光源, 连续测定 3个晴天, 每个指标重复3次, 取3 d的平均值。
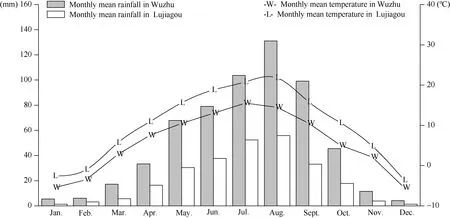
图1 试验区月平均降雨量和气温状况Fig. 1 Dynamic of monthly mean rainfall and temperature in experiment areas
1.4 叶绿素测定
使用便携式叶绿素仪(Minolta, SPAD-502Plus, Japan)测定叶绿素含量。从每小区选择 3株有代表性健康植株,测定第3个展开叶, 对每个叶片测3次, 取平均值。
1.5 植被覆盖指数测定
使用NDVI植被指数测定仪(Spectrum, CM1000, USA)测定植被覆盖指数(normalized difference vegetation index,NDVI)。
1.6 冠气温差的测定
使用红外线热像仪(FLIR, T660, USA)从播种40 d后(苗期)开始, 选择晴天中午12:00—15:00, 从每小区选择3株具有代表性的健康植株采集冠层红外热图像, 每株采集3次。将远红外镜头垂直置植株上方, 对最上部展开叶(距成像仪0.8 m)拍摄图像, 利用ThermaCAM Researcher 2002软件读取样本叶片温度, 选择每株样本 6片均匀无病虫危害、完全展开的叶片, 读取每片叶6处不同位置的温度, 取平均值为冠层温度(canopy temperature,Tc), 在采集每个红外热图像的时间内, 使用电子数显温度计(Dretec, O-230, Japan)重复3次读取空气温度, 取平均值计为空气温度(air temperature,Ta)。冠气温差(canopy temperature depression, CTD) =Ta–Tc。
1.7 耐旱系数DTC值计算
参考 Bouslama等[13]的公式, 耐旱系数(drought tolerance coefficient, DTC)= SA/SH。SA为半干旱条件下(鲁家沟镇)某指标的实测值; SH 为半湿润条件下(五竹镇)某指标的实测值。
1.8 数据分析
采用Microsoft Excel 2003和IBM SPSS Statistic 19.0进行数据统计分析及 SPSS皮尔逊(Pearson)相关系数分析。
2 结果与分析
2.1 不同环境下植株表型性状的耐旱性分析
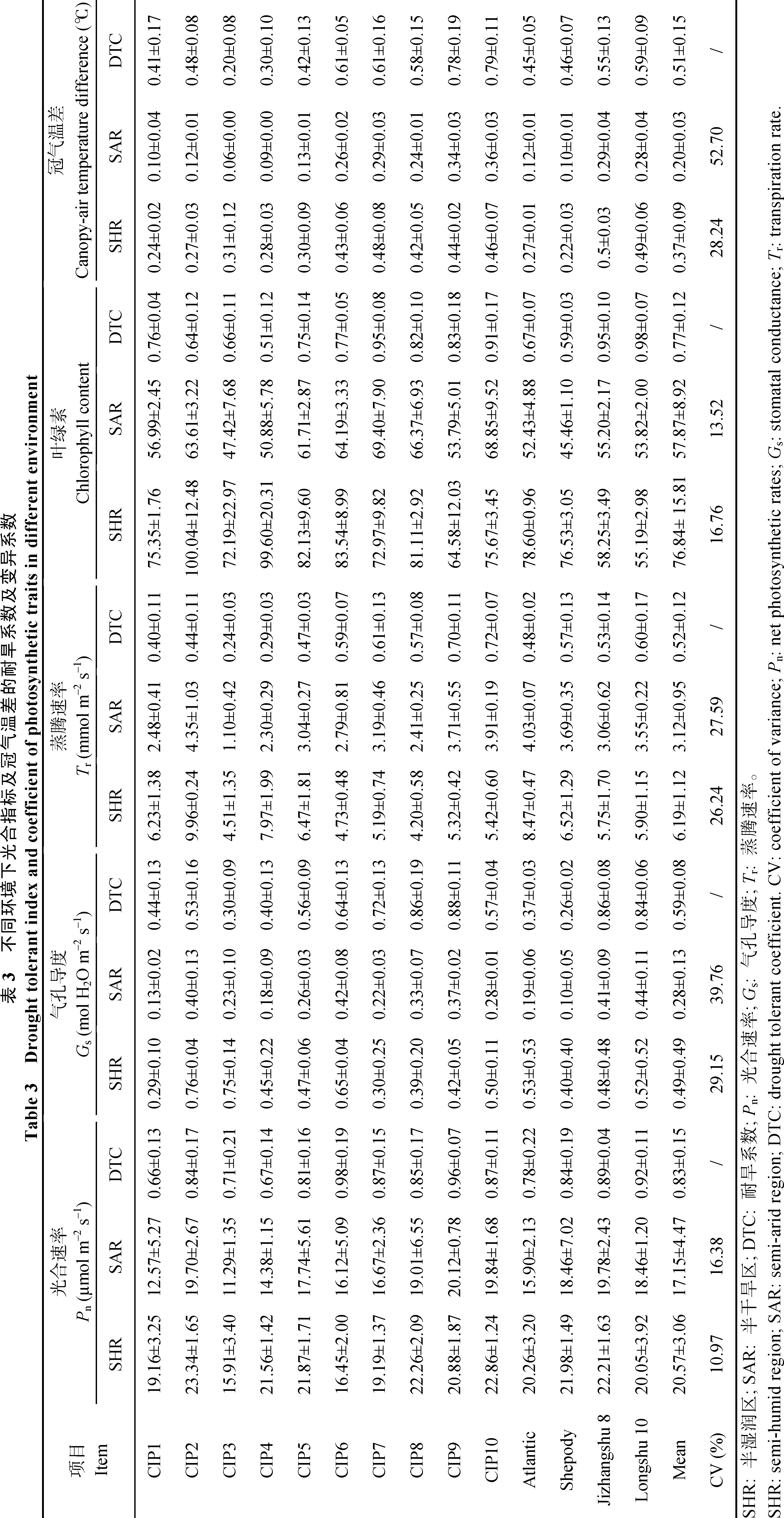
干湿两种种植环境下, 供试马铃薯材料的植株表型性状表现出不同程度的差异。从表2可以看出, 半干旱环境对供试材料的株高、叶面积、叶鲜重、植被覆盖指数均表现出抑制作用。株高、叶面积、叶鲜重、植被覆盖指数在半湿润环境中的变异范围分别为 15.67~28.00、6.14~10.93、0.15~0.35和0.83~0.90, 在半干旱环境中的变异范围分别为 9.00~17.33、5.11~9.32、0.08~0.27 和 0.61~0.75,各性状的平均值在半干旱环境中比在半湿润环境中分别降低34.83%、25.45%、38.36%和19.77%。通过计算得到的耐旱系数表明, 不同供试马铃薯材料的各项表型性状也表现出不同的耐旱性, 其中植被覆盖指数的平均耐旱系数最高, 为0.8, 叶鲜重的平均耐旱系数最低, 为0.6。
2.2 不同环境下冠气温差及光合生理指标的耐旱性分析
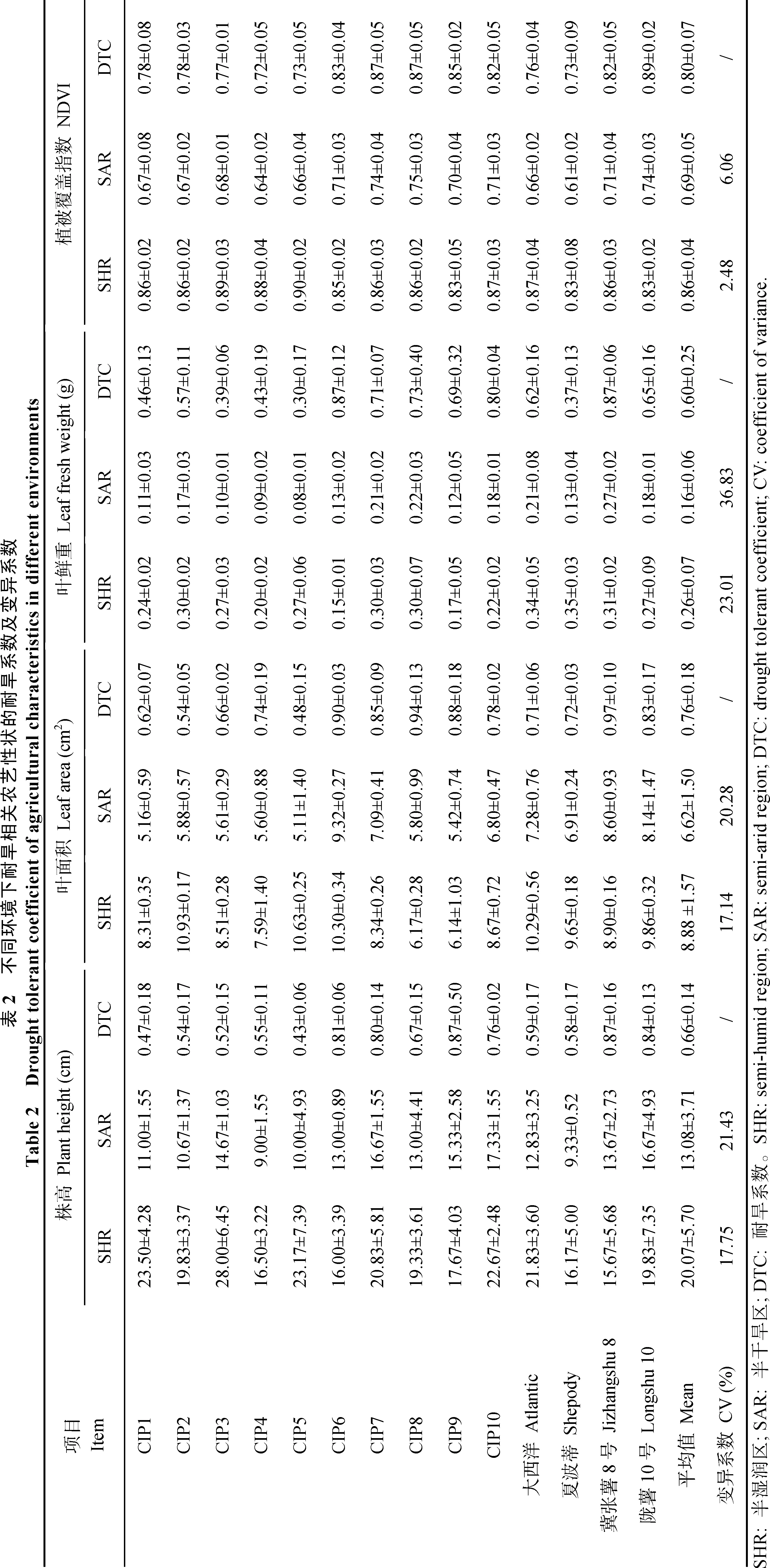
在通过远红外热成像仪拍摄的热成像图中, 根据色板颜色可分辨图像中不同位置的温度情况, 越接近底部蓝黑色代表图像中相应位置温度越低, 越接近顶部黄白色代表图像中相应位置温度越高。从图2可以看出, 不同马铃薯品种在同一环境下叶片温度差异明显, 在半干旱环境下, 马铃薯植株的生物量积累明显受到抑制, 并且在半干旱环境中耐旱品种的叶片温度明显低于干旱敏感型品种。
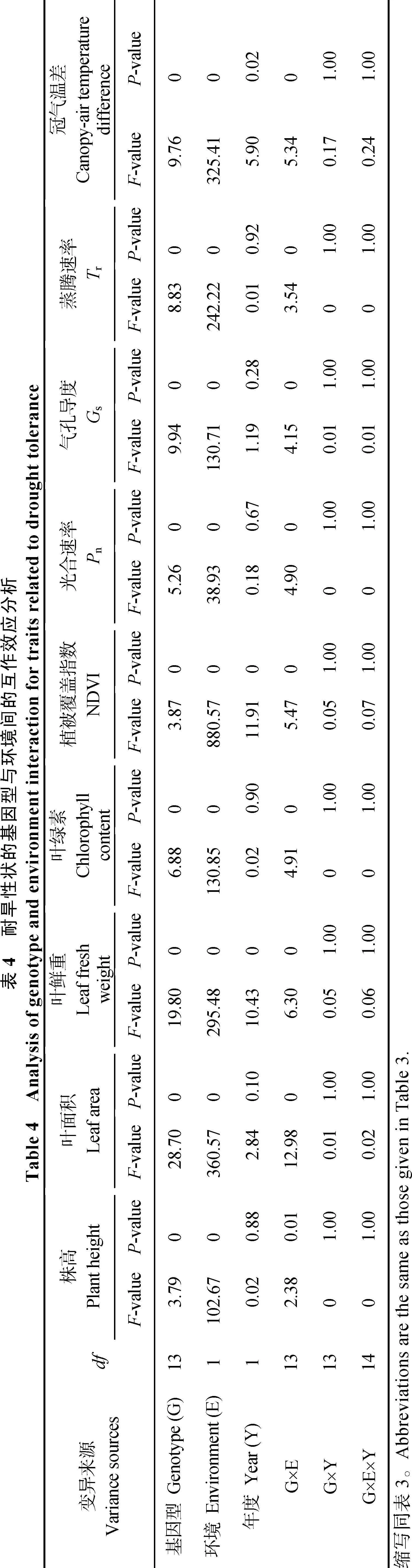
从表3可以看出, 在半干旱环境中, 不同供试马铃薯材料的光合生理指标及冠气温差比在半湿润环境中均有所降低, 光合速率、气孔导度、蒸腾速率、叶绿素、冠气温差在半干旱环境中的平均值相比在半湿润环境中分别降低16.63%、42.86%、49.60%、24.69%和51.52%。各指标平均耐旱系数值表明, 马铃薯光合速率、气孔导度、蒸
腾速率、叶绿素、冠气温差在干旱环境中具有不同程度的耐旱性, 其中光合速率的平均耐旱系数最高, 为 0.83, 其次是叶绿素、气孔导度和蒸腾速率, 冠气温差的平均耐旱系数最低均, 为 0.51。耐旱型品种冀张薯 8号和陇薯 10号以及耐旱型材料CIP6~CIP10在干旱种植条件下表现出较高的冠气温差值。
?

图2 远红外热成像仪拍摄的热成像图Fig. 2 Thermal images taken by infrared imaging systems
2.3 各性状在基因型与环境、年际间的互作效应分析
从表4可以看出, 冠气温差、光合生理指标及植株表型性状在不同供试马铃薯材料之间, 以及干、湿2种种植环境之间均表现出极显著差异性(P<0.01)。在不同年度间,叶鲜重、植被覆盖指数差异极显著(P<0.01), 冠气温差差异显著(P<0.05), 其余性状均无显著性差异。此外, 除株高在基因型与环境(G×E)之间存在显著互作效应(P<0.05)外, 其他性状在基因型与环境(G×E)之间均存在极显著互作效应(P<0.01)。所测性状在基因型与年度(G×Y)之间, 基因型与环境、年度(G×E×Y)之间均无显著互作效应。
2.4 各性状耐旱系数间的相关性分析
从表5可以看出, 供试马铃薯材料在干、湿2种种植环境中冠气温差的耐旱系数与所有光合生理指标及表型性状的耐旱系数均呈极显著正相关, 气孔导度的耐旱系数与叶面积和植被覆盖指数的耐旱系数呈极显著正相关,与叶绿素含量的耐旱系数呈显著正相关, 与株高和叶鲜重的耐旱系数无显著相关性。光合速率和蒸腾速率的耐旱系数分别与叶面积和植被覆盖指数的耐旱系数呈极显著正相关, 与叶鲜重的耐旱系数呈显著正相关, 与其他表型性状的耐旱系数无显著相关性。
3 讨论
3.1 马铃薯耐旱性评价指标
耐旱鉴定指标是马铃薯耐旱育种工作中的重要内容和理论依据[14]。近年来, 研究人员对马铃薯的耐旱机制研究并提出了多项耐旱指标, 可在马铃薯育种评价中应用。在利用植株表型性状鉴定马铃薯耐旱性中, Deblond等[15]在水旱条件下对6个马铃薯品种的茎高、叶片数、叶长测定发现, 这些指标对干旱敏感, 认为可作为马铃薯耐旱性评价指标。Lahlou等[16]调查了4个马铃薯品种的耐旱性,发现叶片干重、叶面积、株高可以作为马铃薯耐旱性评价指标。Richards[17]在干旱条件下分析各性状对马铃薯产量不同贡献率, 发现开花数、早期叶面积可作为马铃薯耐旱性评价指标。在利用光合指标进行耐旱性鉴定中, Bansal等[18]发现马铃薯在干旱胁迫下净光合速率明显下降, 可作为重要的马铃薯耐旱指标。Jefferies等[19]在水、旱2种条件下种植马铃薯, 发现干旱可使马铃薯气孔导度下降,进而限制光合作用, 导致产量降低, 从而发现气孔导度可作为耐旱指标对耐旱型马铃薯进行鉴定。本试验在干、湿2种种植环境下对14个不同马铃薯材料的耐旱性评价表明,
干旱对试验中所测定的马铃薯植株表型性状和光合生理指标均产生了抑制效应, 光合速率和植被覆盖指数的耐旱性最强, 分别为0.80和0.83, 叶面积、叶绿素、株高、叶鲜重耐旱性中等, 介于 0.60~0.77之间, 冠气温差、蒸腾速率和气孔导度耐旱性最弱, 分别为0.51、0.52和0.59,说明马铃薯的冠气温差、蒸腾速率和气孔导度对干旱胁迫的响应最敏感。在以往研究中, 植株表型性状和光合生理指标已被广泛用于马铃薯耐旱性鉴定研究, 近年来, 冠气温差也常被用于植物的耐旱鉴定。冠气温差被认为是反映植物生理适应性的有效指示器, 可以很好地反映某一环境条件下植物的综合生理状态[20], 已有研究证明, 冠气温差可作为有效工具对小麦、水稻、大豆、棉花、玉米等作物进行耐旱型种质筛选[21-26]。本试验中干旱对供试材料的冠气温差均产生了抑制效应, 冠气温差在干、湿2种种植环境之间均表现出极显著差异性, 表明干旱对冠气温差会产生显著影响, 说明冠气温差也可作为鉴定指标在马铃薯耐旱性研究中应用。基因型与环境互作效应可反映品种性状的稳定性, 本试验发现试验中所测性状指标在基因型与环境之间均存在显著互作效应, 说明这些性状受基因型和环境条件的双重影响, 需在多环境条件下进行表型鉴定, 才能获得性状的全面评价。
?
?
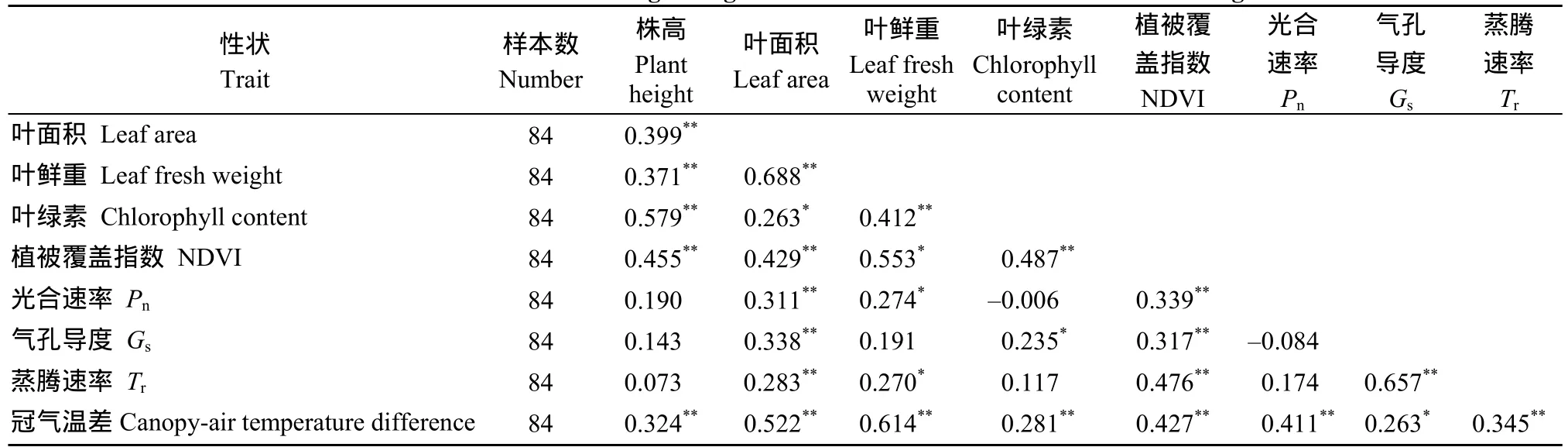
表5 各性状耐旱系数间的相关系数Table 5 Correlation coefficients among drought tolerant indexes of traits related to drought tolerance
3.2 冠气温差与马铃薯耐旱性的关系
作物在田间生长过程中, 能够通过自身对冠层温度的调节来适应干旱、高温等不利环境[27]。冠层温度的变化实际是作物适应不同环境的反应信号, 干旱胁迫下, 作物气孔关闭、蒸腾强度降低, 从而减少水分散失, 维持植株正常生理代谢, 保持作物生物量的积累, 在气孔关闭、蒸腾强度降低后, 植株的一个重要表象就是冠层温度升高, 而这一表象往往提前于植株生物量积累的变化, 且相对于生理代谢的变化更易于观察[28], 所以冠层温度可视为马铃薯在田间受旱时的快速响应指标。而冠层温度会受到所在环境空气温度的显著影响, 用同一时间内的空气温度减去作物冠层温度获得的冠气温差可反映作物在不同环境中的相对温度变化, 这有效排除了外界环境的干扰。在冠气温差与马铃薯耐旱性关系的研究中, 本试验通过公式计算获得了供试马铃薯材料的植株、叶面积、叶鲜重、叶绿素和植被覆盖指数的耐旱系数, 这5个性状可反映马铃薯植株的生物量积累和生理代谢情况, 其耐旱系数是反映马铃薯耐旱性的重要指标, 通过相关性分析表明, 冠气温差的耐旱系数与植株、叶面积、叶鲜重、叶绿素和植被覆盖指数的耐旱系数呈极显著正相关, 此外供试马铃薯材料光合作用指标的耐旱系数与冠气温差的耐旱系数也均呈现出极显著正相关, 说明冠气温差可综合表明马铃薯植株在干旱胁迫下的生物量积累和生理代谢情况, 从而反映马铃薯的耐旱性。
3.3 冠气温差在马铃薯耐旱性鉴定中的应用
冠气温差不仅受环境影响, 也受作物遗传特性的作用, 具体表现在受到干旱胁迫时, 不同基因型作物的冠气温差变化程度差异显著[29]。在本试验中, 冠气温差在不同供试马铃薯材料之间差异极显著。有研究表明, 耐旱型作物品种的冠层温度通常低于干旱敏感型品种的冠层温度, 因而耐旱型作物品种的冠气温差高于干旱敏感型品种[7-8]。在相同条件下, 冠层温度在不同基因型间存在的差异称之为冠层温度分异特性。通过冠层温度分异特性可将作物品种分为冷型品种、暖型品种及中间型品种。通常干旱胁迫下冷型品种较暖型品种会表现出更好的生物学性状, 在耐旱型品种选育中, 利用冷型材料为亲本或以其作为主要选育目标的研究已在小麦中多有报道[30]。在本试验中, 耐旱型品种冀张薯8号和陇薯10号以及5个耐旱型材料在干旱种植条件下表现出较高的冠气温差值,并在植株表型性状和光合指标上表现出较好的耐旱性,从而说明冠气温差可以作为马铃薯的耐旱性鉴定指标予以应用。此外, 在作物对干旱胁迫的反应上, 冠层温度和空气温度相对较容易测定, 因此冠气温差是一个较为理想的田间鉴定指标。并且在本试验中使用的红外线热像仪可在不损伤马铃薯植株的条件下对冠层温度进行快速检测, 使马铃薯耐旱性鉴定的方法更为便捷高效。
