干旱胁迫对八宝景天叶片生理的影响
2018-04-19刘紫娟李炳言贾斯淳宗毓铮郝兴宇
刘紫娟,袁 蕊,王 娜,李炳言,贾斯淳,宗毓铮,李 萍,郝兴宇
(山西农业大学农学院,山西太谷 030801)
工业革命以来,全球气温持续上升,气候变暖同时也影响全球水资源的分布、降水量的变化,导致部分地区干旱[1]。水分是植物生长的必需底物,植物的基因表达和新陈代谢都会受到干旱的影响。我国是缺水国家,在干旱和半干旱地区,由于降水量少、土壤水分缺乏,不仅造成作物减产,也严重影响了林业发展和生态环境[2]。因此,探究植物适应干旱胁迫的生理机制对干旱区植物生长和生态发展具有一定意义。
八宝景天(Hylotelephium erythrostictum)又名八宝、活血三七、对叶景天,为景天科八宝属多年生草本植物[3]。八宝景天是景天酸代谢(CAM)植物,叶肉质,喜强光和干燥,管理方式粗放,病虫害较少,是一种常见的植物,由于其具有较高的观赏价值和较低的养护成本,在城市建设中被广泛应用于园林绿化,特别是近些年快速发展的屋顶绿化中[4]。CAM植物由于其代谢方式特殊,均有一定的抗旱性,国内外学者针对CAM植物已有许多研究,多探讨CAM植物的光合代谢方式[5-7],针对其抗旱性的研究主要集中在铁皮石斛、垂盆草、东南景天、细叶景天、红景天等[8-10],对八宝景天的相关研究较少,特别是一天中白天和夜晚生理变化的研究还未见报道。
本试验拟探究干旱胁迫条件下八宝景天的叶片生理变化特征,旨在为CAM植物的抗旱机理研究奠定基础,为八宝景天今后的应用和发展提供理论依据。
1 材料和方法
1.1 试验材料
供试景天品种为八宝,由山西农业大学园艺学院提供。
1.2 试验设计
试验在山西农业大学试验基地(37.42°N,112.58°E)旱棚中进行,旱棚高3.5 m,面积为4 m×4 m。八宝景天于2016年4月24日移植于长×宽×高为60 cm×40 cm×35 cm塑料箱中,箱子底部打5个孔用于排水,箱内装土约28cm高,2016年10月20日割除八宝景天的地上部,留根于土中,2017年3月25日发芽。共栽植10盆八宝景天,前期正常灌水。5盆作对照处理(CK),植株发芽后控制土壤含水量为土壤饱和含水量的70%~80%;另外5盆作干旱处理(T),植株发芽30 d后(之前无水分胁迫)控制土壤含水量为土壤饱和含水量的30%~40%,其他管理措施一致。
1.3 生育期的确定
八宝景天开始发芽至发芽后107 d为营养生长期,期间植株茎伸长增粗,叶片增多,发芽后107~118 d陆续有植株长出花蕾,发芽后第119天有80%以上植株长出花蕾,进入现蕾期。
1.4 测定项目及方法
1.4.1 叶片表面气孔的测定 在营养生长期(发芽后102 d)10:00和22:00,每个处理分别选取有代表性的八宝景天5株(每箱1株),每株选取倒数第7片完全展开叶,用指甲油粘片法测量并计算气孔密度[11];在40倍物镜下选取视野内8个气孔,测量其长度、宽度,并测量气孔开度的长度和的宽度,计算气孔开度。气孔开度(c)=π·ab,其中,a=1/2 气孔开度的长度,b=1/2气孔开度的宽度[12]。
1.4.2 叶片抗逆指标的测定 在营养生长期(发芽后 102 d)和现蕾期(发芽后 119 d)的 10:00和22:00,每个处理分别选取有代表性的植株5株(每箱1株),每株选取其倒数第5片完全展开叶,用于测定抗逆生理指标。采用愈创木酚比色法测定过氧化物酶(POD)活性,采用硫代巴比妥酸显色法测定丙二醛(MDA)含量,采用酸性茚三酮比色法测定脯氨酸(Pro)含量[13]。
1.4.3 叶片含水量的测定 利用1.4.2选取的叶片,剪成0.5 cm小段后测鲜质量,然后于60℃烘箱中12 h烘干,测干质量。含水量=(鲜质量-干质量)/鲜质量×100%。
1.4.4 叶片苹果酸含量的测定 利用1.4.2选取的叶片,采用Goodban法进行苹果酸含量的测定[7]。
1.4.5 叶片色素含量的测定 利用1.4.2选取的叶片,用丙酮∶乙醇∶蒸馏水为4.5∶4.5∶1的混合液作为叶绿素提取液,再用分光光度计测定吸光值,并计算光合色素含量。
1.4.6 叶片糖类物质含量的测定 利用1.4.2选取的叶片,采用3,5-二硝基水杨酸法测定还原糖含量,采用蒽酮-硫酸法测定可溶性糖、淀粉和纤维素含量[14]。
1.4.7 各项形态指标的测定 于植株现蕾期将地上部分收获,对未取样植株分别测量其株高、茎粗、叶片数,于60℃烘箱48h烘干,测定单株总生物量。
1.5 数据处理
利用Excel 2007软件对数据进行统计分析和作图。用SPSS19软件对数据进行显著性分析(α=0.05)。图表中数据均为平均值±标准差。
2 结果与分析
2.1 干旱胁迫对八宝景天叶片气孔的影响
干旱胁迫使八宝景天叶片上下表皮细胞变小,上下表皮的气孔长度和开度减小,但气孔密度增加(图1)。与对照相比,干旱胁迫使八宝景天叶片上表皮的气孔长度在10:00和22:00分别显著降低19.9%和17.3%,10:00和22:00的气孔开度分别显著降低30.7%和16.6%,但是气孔密度分别显著增加36.8%和28.8%;下表皮的气孔长度在10:00和22:00分别显著降低23.7%和20.4%,气孔开度在10:00和22:00分别显著降低49.2%和11.5%,但 气孔密度增加(表1)。

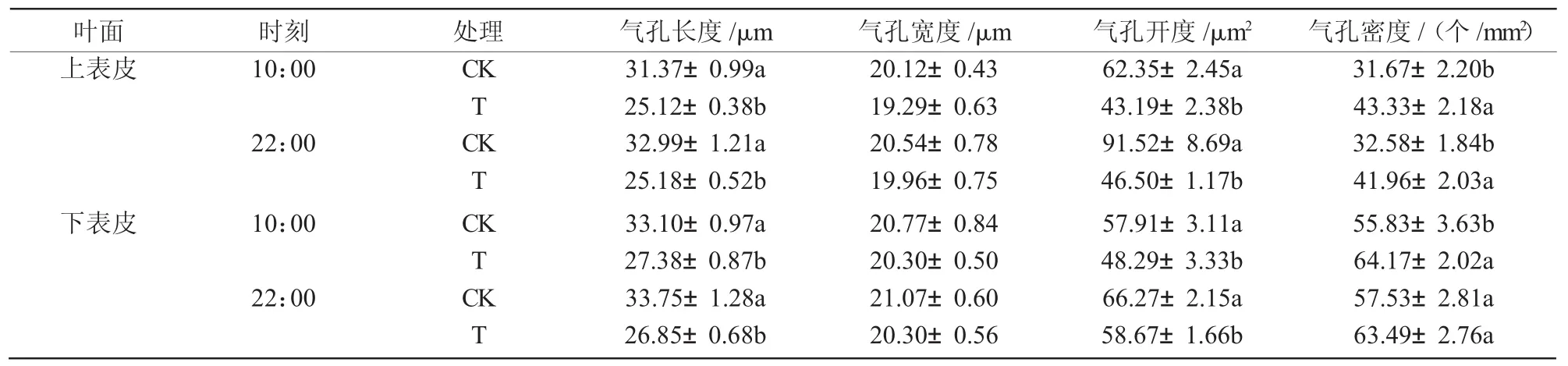
表1 干旱胁迫对八宝景天气孔的影响
2.2 干旱胁迫对八宝景天叶片含水量的影响
干旱胁迫下,八宝景天叶片含水量在营养生长期和现蕾期的10:00和22:00均无显著变化(图 2)。

2.3 干旱胁迫对八宝景天叶片MDA含量的影响
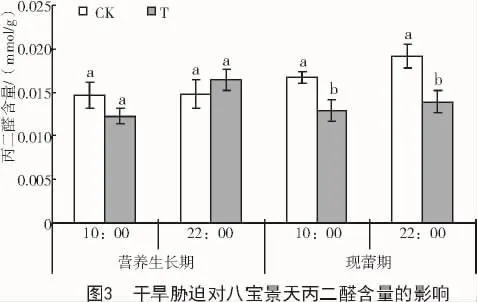
干旱胁迫使八宝景天叶片MDA含量在营养生长期 10:00,现蕾期 10:00 和 22:00 均有降低,在营养生长期22:00有所升高。其中,现蕾期10:00和22:00分别比对照显著降低22.6%和27.5%(图3)。
2.4 干旱胁迫对八宝景天叶片POD活性的影响
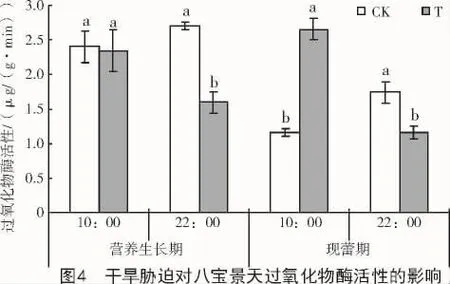
与对照相比,干旱胁迫使八宝景天叶片POD活性在营养生长期和现蕾期的22:00分别显著降低41.0%和33.7%,在现蕾期10:00显著升高128.3%,但在营养生长期10:00无明显变化(图4)。
2.5 干旱胁迫对八宝景天叶片脯氨酸含量的影响
与对照相比,干旱胁迫使八宝景天脯氨酸含量在营养生长期10:00和22:00分别显著升高3.5%和1.8%,但在现蕾期脯氨酸含量变化不显著(图5)。

2.6 干旱胁迫对八宝景天叶片苹果酸含量的影响
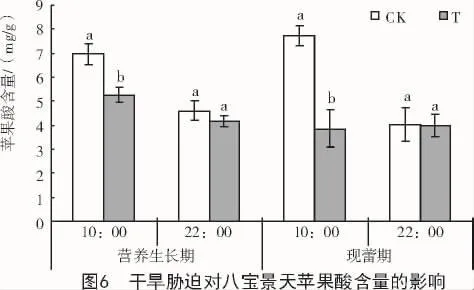
干旱胁迫条件下,八宝景天叶片苹果酸含量在营养生长期和现蕾期10:00分别比对照显著降低24.7%和50.1%,但在2个时期22:00均无明显变化(图 6)。
2.7 干旱胁迫对八宝景天叶片光合色素的影响
在干旱胁迫条件下,八宝景天叶片的叶绿素a、叶绿素b和类胡萝卜素含量在营养生长期和现蕾期的10:00均为升高趋势,其中,叶绿素a含量在营养生长期和现蕾期10:00分别比对照显著升高36.4%和32.5%;而在2个时期的22:00均呈下降趋势,其中,营养生长期22:00叶绿素b含量显著降低24.4%,现蕾期22:00叶绿素a含量显著降低23.8%(表 2)。
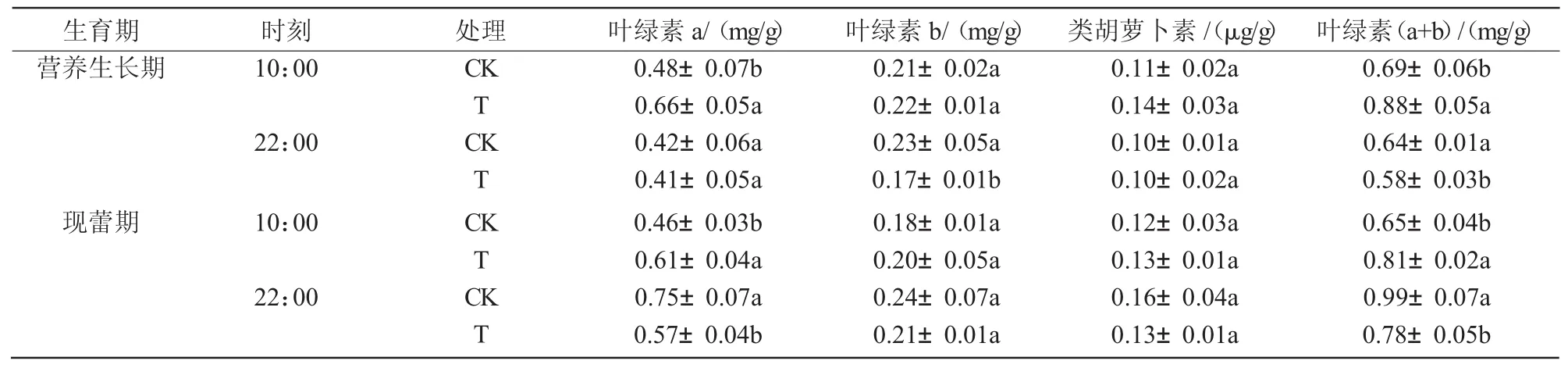
表2 干旱胁迫对八宝景天光合色素的影响
2.8 干旱胁迫对八宝景天叶片糖类物质含量的影响
与对照相比,干旱胁迫使八宝景天叶片中还原糖、可溶性糖、淀粉和纤维素在整个生育期全天总体上呈上升趋势,其中,还原糖含量在2个时期的22:00分别升高24.6%和45.8%,可溶性总糖含量在现蕾期10:00和22:00分别显著升高25.3%和24.7%,淀粉含量在营养生长期10:00,22:00和现蕾期10:00分别显著升高40.3%,35.9%和147.1%,纤维素含量在营养生长期10:00和22:00分别显著升高31.7%和25.2%,但在现蕾期却呈下降趋势,其中,10:00 时显著下降 36.6%(表 3)。
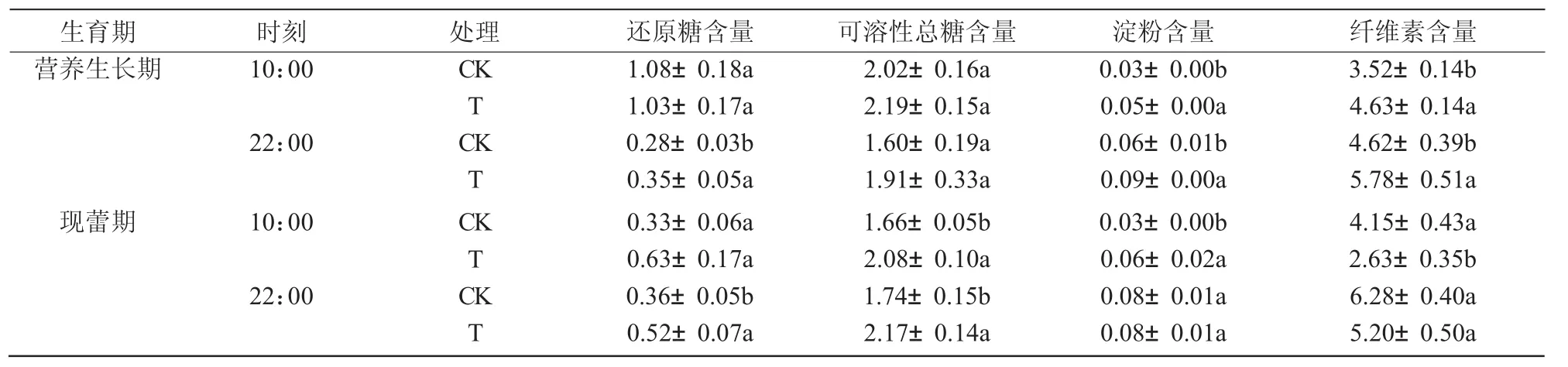
表3 干旱胁迫对八宝景天叶片糖类含量的影响 %
2.9 干旱胁迫对八宝景天各项形态指标的影响
与对照相比,干旱胁迫使八宝景天株高、茎粗、单株叶数和单株总生物量分别显著降低了36.2%,32.3%,24.6%和35.2%(表4)。

表4 干旱胁迫对八宝景天各项形态指标的影响
3 讨论
以往的研究通常认为,干旱胁迫会促使植物叶片气孔关闭或密度减小,从而减少植物蒸腾作用中水分的散失,提高植物的抗旱性[15];但也有研究显示,在干旱胁迫下,植物气孔密度增大,但其长度、宽度和开度均减小[16]。本试验中,八宝景天在干旱胁迫下叶片上下表皮气孔的长度、宽度和开度均减小,但气孔密度增加。气孔密度增加的原因可能是由于干旱景天叶面积减小,使单位叶面积气孔密度增加。而气孔长度、宽度和开度减小的原因是由于干旱影响了气孔的发生、分化和发育,使气孔变小。气孔变小使蒸腾减弱,提高水分利用效率[17]。
干旱胁迫会破坏植物的细胞膜系统,使植物体内自由基增加,从而引起膜脂过氧化作用,膜脂过氧化的产物是MDA,其含量的多少可表示膜脂的过氧化程度和植物的抗逆性[18]。本试验中,干旱胁迫使八宝景天MDA含量在营养生长期10:00和现蕾期均降低,可能是由于八宝景天具有较强的抗旱性,干旱对叶片的膜脂过氧化作用影响不大。植物中的膜脂过氧化防御系统会保护细胞免遭伤害,POD是植物体内清除H2O2的主要酶类之一,能催化H2O2氧化后产生H2O[19]。本试验中,干旱胁迫使八宝景天POD活性在22:00时均下降,现蕾期10:00升高,白天温度较高,可能需要更多的POD防御膜脂的过氧化,夜间温度降低,干旱胁迫有所缓解,所以,POD活性下降。Pro是一种渗透调节物,可以增强植物的抗旱能力,本试验中,干旱胁迫下八宝景天叶片的Pro含量除现蕾期10:00外均增加,这也反映出八宝景天是一种抗旱植物。干旱胁迫下植物叶片的含水量可以反映植物保水能力的大小。本试验中,干旱胁迫对八宝景天叶片的含水量并无显著影响,说明八宝景天在通过气孔调节和Pro、糖类物质等渗透调节物质的调节后,在干旱胁迫下有较好的保水能力。
八宝景天是CAM植物,光合方式有其特殊性,表现为:夜间气孔开放,吸收CO2,以苹果酸的形式贮存,白天苹果酸分解,释放CO2,进行光合作用[7]。本试验中,干旱胁迫使八宝景天叶片苹果酸含量在上午10:00显著下降,说明干旱胁迫抑制了夜间苹果酸的积累,这是因为干旱胁迫使气孔开度减小,影响了CO2的吸收。
叶绿素a、叶绿素b和类胡萝卜素是植物进行光合作用所需的最主要的色素,其含量的多少会直接影响植物的生长发育[20]。本试验中,干旱胁迫使八宝景天的叶绿素a、叶绿素b和类胡萝卜素含量在营养生长期和现蕾期10:00升高,但22:00下降,这与之前的研究结果有所不同,可能是由于八宝景天在干旱条件下会产生更多光合色素促使光合作用的进行,但在夜间不进行光反应时,为维持自身正常生理活动,光合色素含量下降。但其调节机制有待进一步研究探索。
可溶性糖也是植物体内参与渗透调节的重要物质,植物在遭受胁迫时,可以通过调节可溶性糖的多少来维持正常的渗透压,淀粉在植物遭受逆境时可以与可溶性糖相互转化,纤维素是细胞壁的主要组成部分,可以使植物保持挺拔[21-23]。本试验中,还原糖、可溶性糖和淀粉含量在不同时期均有增加的趋势,这也是八宝景天抵抗干旱的一种形式。纤维素含量在营养生长期均增加,但在现蕾期降低,可能是由干旱后期植株缺水时间过长、碳素积累受限所致。
干旱胁迫会使植株的生长形态发生变化,表现在细胞、组织、器官个体等不同水平[24-25]。一般认为,干旱胁迫会使植物的株高、茎粗、生物量均降低,导致作物减产[26]。本试验中,干旱胁迫使八宝景天株高、茎粗、单株叶片数和单株生物量都降低,叶片的减少和植株矮小可以减少植株水分的散失和植物对水分的需求,有利于植株抵抗干旱环境。
在投融资方案上充分发挥政府投资对社会资本的引导放大作用,吸引社会资本参与。要坚持以市场为导向,以制度建设为重点,形成市场融资为主导、政府投资为补充、民间投资健康发展的投融资格局。在旅游、医疗、教育、停车等社会化产业服务领域,坚持以政策导向引领,社会资本主导参与建设为原则,通过出让特许经营权、第三方付费、使用者付费结合少量政府补助的方式,鼓励社会资本加大此类应用的建设力度和速度。利用社会资本的灵活性和企业服务的创新性实现智慧城市的长期可持续发展。
4 结论
本研究表明,干旱胁迫下,八宝景天叶片气孔变小,气孔开度降低,能够有效地减少水分散失,同时影响夜间CO2的吸收,降低了苹果酸的积累;干旱胁迫使八宝景天MDA含量降低,使POD活性白天升高,夜间降低。现蕾期Pro含量、可溶性总糖和淀粉含量均升高,以提高其抗旱性;干旱胁迫下,八宝景天叶片光合色素含量在白天增多,夜晚降低,有利于其光合效率的提高。干旱使八宝景天株高、茎粗、单株叶片数和单株总生物量均降低,可以有效地减少水分消耗。总之,八宝景天在干旱胁迫下通过气孔开度变小,POD活性增加进行膜脂过氧化防御,Pro和可溶性糖含量增加进行渗透调节,提高日间光合色素含量保证光合作用的有效进行等方式提高其干旱适应能力,表现了较强的抗旱性。
参考文献:
[1]WALLACE J S.Increasing agricultural water use efficiency to meet future food production[J].Agriculture,Ecosystems and Environment,2000,82:105-119.
[2]鲁松.干旱胁迫对植物生长及其生理的影响[J].江苏林业科技,2012,39(4):51-54.
[3]中国科学院中国植物志编辑委员会.中国植物志 [M].北京:科学出版社,1984:54-55.
[4]陈江霞.八宝景天及应用领域[J].林木花卉,2009(11):46-47.
[5]李明启.景天科酸代谢的碳途径:植物代谢多条途径观点的一个典型例子[J].植物生理学通讯,1994,30(4):286-292.
[6]张泽锦.铁皮石斛的光合碳同化途径及其对环境变化的生理响应[D].北京:中国农业大学,2014.
[8]李云霞,张建生,吴永华,等.5种景天科地被植物抗旱性比较研究[J].干旱区资源与环境,2010,24(2):183-186.
[9]张斌,胡永红,刘庆华,等.几种屋顶绿化景天植物的耐旱性研究[J].中国农学通报,2008,24(5):272-276.
[10]李滨胜,周玉迁,潘杰,等.干旱胁迫下细叶景天生理生化指标的变化[J].北方园艺,2010(16):105-107.
[11]张立荣,牛海山,汪诗平,等.增温与放牧对矮嵩草草甸4种植物气孔密度和气孔长度的影响 [J].生态学报,2010,30(24):6961-6969.
[12]徐萍,李进,吕海英,等.干旱胁迫下水杨酸对银沙槐子叶表皮气孔开度的影响[J].植物生理学报,2014,50(4):510-518.
[13]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:164-165,260-261.
[14]李强,唐薇,石园园,等.蒽酮-硫酸法和3-5-二硝基水杨酸法测定杜仲水提液多糖含量 [J].食品工业科技,2010(10):370-371,374.
[15]张岁岐,李金虎,山仑.干旱下植物气孔运动的调控[J].西北植物学报,2011,21(6):1263-1270.
[16]于海秋,王晓磊,蒋春姬,等.土壤干旱下玉米幼苗解剖结构的伤害进程[J].干旱地区农业研究,2008,26(5):143-147.
[17]杨惠敏,王根轩.干旱和CO2浓度升高对干旱区春小麦气孔密度及分布的影响[J].植物生态学报,2001,25(3):312-316.
[18]胡学俭,孙明高,夏阳,等.NaCl胁迫对无花果与海棠膜脂过氧化作用及保护酶活性的影响 [J].西北植物学报,2005,25(5):937-943.
[19]邵艳军,山仑.植物耐旱机制研究进展[J].中国生态农业学报,2006,14(4):17-20.
[20]徐秀梅,张新华,王汉杰.四翅滨藜抗旱生理特性研究[J].南京林业大学学报(自然科学版),2004,28(5):54-58.
[21]牟凤娟,陈丽萍,李军萍,等.干旱胁迫下旱地油杉生理生化指标动态变化[J].干旱区资源与环境,2016,30(5):180-184.
[22] KLEIN T,HOCH G,YAKIR D,et al.Drought stress,growth and nonstructural carbohydrate dynamics of pine trees in a semi-arid forest[J].Tree Physiology,2014,34(9):981-992.
[23] O'BREIN M J,LEUZINGER S,PHILIPSON C D,et al.Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels[J].Nature Climate Change,2014,4(8):710-714.
[24]丛建鸥,李宁,许映军,等.干旱胁迫下冬小麦产量结构与生长、生理、光谱指标的关系 [J].中国生态农业学报,2010,18(1):67-71.
[25]何晓莹.初花期干旱对油菜农艺性状的影响 [J].山西农业科学,2016,44(9):1291-1293.
[26]王春乙,娄秀荣,王建林.中国农业气象灾害对作物产量的影响[J].自然灾害学报,2007,16(5):38-43.
