60Co诱变大豆滞绿突变体M6变异分析
2018-04-19温宏伟李贵全
温宏伟,王 鹏,李贵全
(山西农业大学农学院,山西太谷 030801)
在植物衰老的过程中,叶片叶绿素降解,出现黄化现象,植株的生物产量减少。有些植株在生育末期叶片衰老过程中,叶绿素不降解或降解不明显,叶片和果实保持绿色,这种现象称为滞绿(stay-green)现象[1]。豆科植物中普遍存在滞绿现象[2],如菜豆[3]、豌豆[4]等突变体,滞绿突变体是研究叶绿素代谢、植物衰老进程等过程的理想材料。在大豆滞绿突变体中,存在2个隐性滞绿基因,分别为子叶滞绿基因D1和种皮滞绿基因D2,其中,D1被定位在大豆第1条染色体上;D2基因被定位在第11条染色体上[5],Gm SGR1和Gm SGR2基因分别负责大豆子叶和种皮滞绿[6]。
大豆种皮色及大豆子叶色会影响大豆的外观,而大豆外观与其商品质量存在着密切关系。有研究发现,导致种皮颜色差异的自然产物包括类黄酮和花青素[7],对抗氧化性的药用价值和营养价值产生一定的影响,因此,种皮色是一个育种后代评估的重要形态标记,有 5 个位点(I,T,W1,R,O)参与控制其性状的形成[8]。
不同株系、不同品种农艺性状差异较大[9-10],滞绿突变体外在表型特殊具有一定的研究价值。本试验利用60Co-γ射线辐射处理Z-94320(双青豆)大豆风干种子,在M6获得108个突变系,对其中的40个突变系从农艺性状、生物学性状、衰老时期叶片叶绿素含量及蛋白质方面进行分析,旨在为大豆滞绿突变体的利用及种皮颜色的研究提供理论依据。
1 材料和方法
1.1 试验材料
2012年,采用60Co-γ射线辐射处理滞绿突变体Z-94320(双青豆)大豆风干种子,剂量率为1.6 Gy/min,种植收获得到M1,2013年种植得到M2,在M2中选择优良变异单株以及特殊变异个体,经过几年的连续选育,到2017年获得M6,共108个突变体品系。
1.2 试验方法
2016年5月9日在山西农业大学农学院大豆育种试验田播种M5种子,收获M6突变体108种材料,行长5 m,行距0.5 m,株距0.25 m,重复3次,常规田间管理,生育期调查记录主要生物性状,利用SPAD-502叶绿素测定仪测定叶片叶绿素含量。成熟后,每株系随机抽取5株进行农艺性状室内考种,测产。测定的农艺性状主要有株高、株质量、结荚高度、茎粗、主茎节数、有效分枝、主茎荚数、分枝荚数、1粒荚数、2粒荚数、3粒荚数、4粒荚数、瘪粒荚数、总荚数、虫食数、百粒质量以及单株粒质量[11]。用InfratecTM 1241 Grain Analyzer V5.00品质分析仪对大豆的蛋白质含量进行测量。对照为滞绿突变体 Z-94320(双青豆)(CK)。
1.3 数据分析
利用SPSS19.0,Excel软件对数据进行分析。
2 结果与分析
2.1 滞绿突变体后代主要生物学性状的变异分析
对M6突变系中的108个株系的17个农艺性状进行测定,结果如表1所示。由表1可知,M6突变系中在株质量、株高、茎粗、主茎节数、2粒荚数、3粒荚数、总荚数、单株粒质量这8个农艺性状方面的均值和CK相比,变化较大。M6突变系中各农艺性状的变异系数从大到小分别为:4粒荚数(115.04%),分枝荚数(87.93%),瘪粒荚数(71.59%),虫食数(65.45%),3粒荚数(64.41%),有效分枝(52.22%),1粒荚数(49.51%),2粒荚数(48.04%),结荚高度(45.42%),单株粒质量(44.30%),株质量(43.60%),总荚数(39.68%),主茎荚数(27.92%),百粒质量(8.02%)。结果表明,在M6突变系中可以着重从4粒荚数、分枝荚数、3粒荚数以及结荚高度和单株粒质量这几个具有潜力的农艺性状中选择适合选育目标的材料。
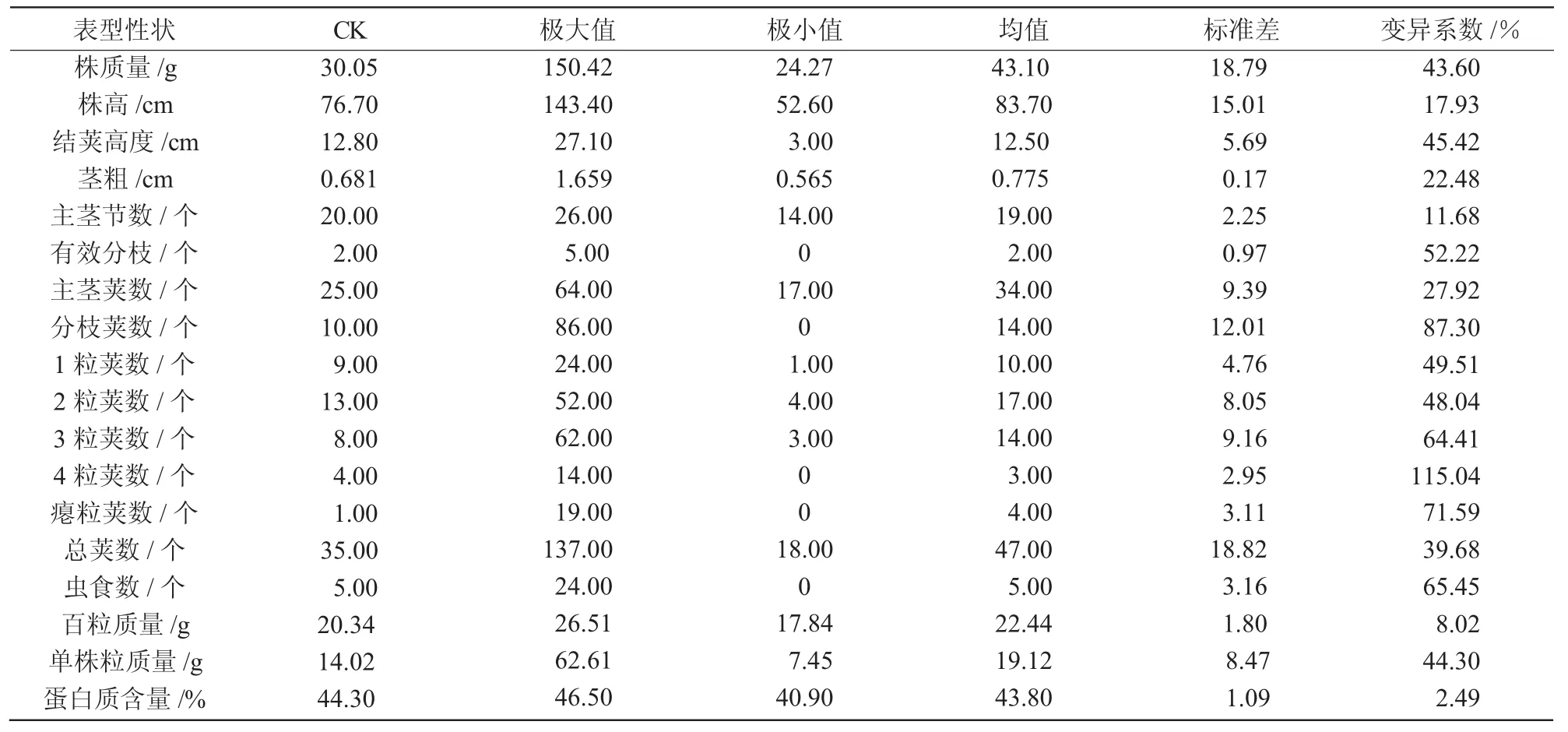
表1 突变体M6农艺性状及蛋白质均值和变异系数
从图1可以看出,大豆滞绿突变体诱变后代中,相对于对照的种皮,诱变后代中出现了种皮色的变异(A2),种皮颜色为绿到黄以及中间的过渡色(A1,A2),在M6的 108个突变系中,按种皮色分类可将其分为 4 类(I,II,III,IV),以 12 号为代表的突变系I中,种皮色为绿色,子叶色为绿色;62号代表的突变系II中,种皮色为浅绿色,子叶色为绿色;104号和74号代表的突变系III中,种皮色为浅绿色,子叶色为黄色;96号和106号代表的突变系IV中,种皮色为黄色,子叶色为黄色,而对照株系种皮色和子叶色均为绿色。M6突变系中豆荚色发生变异(A4),出现黄褐色(95)、浅褐色(98)、褐色(90)、深褐色(19)4种。在M6突变系中出现分枝数增多、茎秆变粗的株系(A5),16号材料可作为突变系中重点关注的材料。

2.2 滞绿突变体后代叶片叶绿素含量比较
从M6突变系中选出40个突变系,按种皮色和子叶色分为 4 类(I,II,III,IV),每一类 10 个株系。
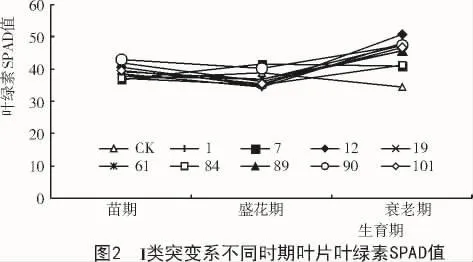

从图2,3可以看出,种皮色和子叶色均为绿色的CK和M6突变系在苗期和盛花期叶绿素含量变化不大,大部分材料在衰老期叶绿素含量相对于盛花期呈现上升的趋势。在II类突变系中,苗期到盛花期间叶绿素含量变化不大,在盛花期到衰老期大部分突变系叶绿素含量保持不变,少数突变系叶绿素含量呈上升趋势。
从图4可以看出,在III类突变系中,在苗期到盛花期间不同株系的叶绿素含量有升有降,其中,28,65号突变系在这个期间呈现上升的趋势。在盛花期到衰老期间,在衰老的过程中,74号材料的叶绿素含量呈现上升趋势,其他材料叶绿素含量均呈现下降趋势。

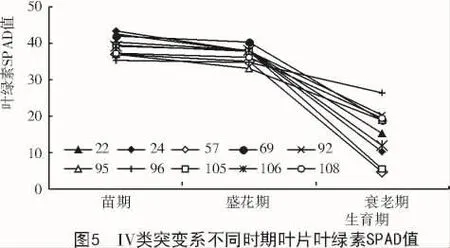
由图5可知,在IV类突变系中苗期到盛花期叶绿素含量变化不大,在衰老期叶绿素含量降幅明显。结果表明,在大豆植株衰老的过程中,种皮色和子叶色与叶片叶绿素含量有着一定的关系,种皮色和子叶色均为绿色的突变系中,存在着滞绿现象,种皮色越绿叶片叶绿素含量越高,而黄色的突变系中存在黄化现象,导致衰老期叶片叶绿素下降。种皮色一致的情况下,在大豆衰老期,子叶色为绿色的突变系比黄色的突变系叶片叶绿素含量下降幅度小甚至出现叶绿素含量上升的趋势。
2.3 滞绿突变体后代蛋白质分析
由表1可知,M6的108个突变系中蛋白质含量的变幅为40.9%~46.5%,与对照株系的44.3%相比,有增有减,蛋白质的变异系数为2.49%。对M6选出的40个株系的蛋白质研究发现,M6-84的蛋白质含量最高,达到46.5%,其种皮色和子叶色均为绿色;M6-105的蛋白质含量最低,为40.9%,其种皮色和子叶色均为黄色。蛋白质均值从大到小排序为:I(44.06%)>II(43.91%)>III(42.84%)>IV(42.78%)(表2)。结果表明,大豆滞绿突变体中,绿色种皮、绿色子叶的大豆蛋白质含量相对高于黄色种皮、黄色子叶的大豆突变系,且滞绿大豆的蛋白质含量相对较高,均值达到44%左右。

表2 滞绿突变体M6蛋白质含量及其均值 %
3 讨论与结论
利用辐射诱变技术,育种家已经选育出大量的优良品种,如水稻脆性突变体[12]等。杨凯敏等[13]对60Co诱变晋大78的M5表型性状进行变异分析,为诱变育种帮助加快选育高产大豆品种提供一定理论依据。本试验对大豆滞绿突变体M6的农艺性状、生物性状、衰老期叶片叶绿素、蛋白质的变异及变化进行分析,结果表明,108个突变系中,4粒荚数、分枝荚数、3粒荚数及结荚高度和单株粒质量变异潜力大,例如,对照的单株粒质量为14.02 g,而诱变后代的突变系中单株粒质量变幅为7.45~62.61 g,变异系数为44.3%,在产量方面具有巨大的潜力,从中可选育出高产突变系。有研究通过对不同百粒质量野生大豆的生物学性状分析发现,随着百粒质量的增加,大豆粒色逐渐由以黑色为主到出现了多种颜色,脐色由以黑色为主到有褐色和深褐色[14]。在突变系中,种皮颜色出现了变异,呈现出从绿到黄色的过渡变异,将其分为4种类型:I.种皮色、子叶色均为绿色;II.种皮色为浅绿色,子叶色为绿色;III.种皮色为浅绿色,子叶色为黄色;IV.种皮色和子叶色均为黄色。子叶色出现了黄色变异,可能是控制种皮色和子叶色的基因位点发生了改变或缺失,有待进一步从基因层面上进行研究。
滞绿突变体诱变后代滞绿特性有减弱趋势[15],通过对突变系叶片叶绿素分析发现,在大豆衰老期过程中,叶片叶绿素含量变化与种皮色和子叶色有一定的关系,种皮色越绿,叶片叶绿素含量越高,且不降解,认为这类突变系为功能性突变体[16];而黄色的突变系中,衰老期叶片叶绿素下降。种皮色一致的情况下,在大豆衰老期,子叶色为绿色的突变系比黄色的突变系叶片叶绿素含量下降幅度小甚至出现叶绿素含量上升的趋势,这可能与衰老期叶绿素降解受阻有关。辐射诱变对品质的改良起到一定作用[17-18],本研究通过蛋白质分析,结果表明,大豆滞绿突变体中,绿色种皮、绿色子叶的大豆蛋白质含量相对高于黄色种皮、黄色子叶的大豆突变系,且滞绿大豆的蛋白质含量相对较高,均值达到44%左右。这与薛树鹏等[19]对大豆科丰14研究得出的诱变后代蛋白质含量降低结果不一致,可能是由于突变系材料不一致导致的。今后可以在分子基因方面,深入研究种皮色、子叶色与品质方面的关系,以获得优质的突变系材料,为育种工作作出新的贡献。
参考文献:
[1]孙佩光,吴琼,徐碧玉,等.植物滞绿机理研究进展[J].基因组学与应用生物学,2015,34(2):438-448.
[2]任钧,王晓磊,高炯,等.国内大豆品种资源中滞绿(stay-green)性状的初步研究[J].植物生理学报,2014,50(9):1336-1346.
[3]RNNINGCM,BOUWKAMP J C,SOLOMOST.Observations on the senescence of a mutant non-yellowing genotype of Phaseolus vulgaris L.[J].J Exp Bot,1991,42:235-241.
[4] THOMAS H,SCHELLENBERG M,VICENTINI F,et al.Gregor mendel's green and yellowpea seed[J].Bot Acta,1996,109:3-4.
[5] FANG C,LI C,LI W,et al.Concerted evolution of D1 and D2 to regu-late chlorophyll degradation in soybean[J].Plant J,2014,77(5):700-712.
[6] NAKANO M,YAMADA T,MASUDA Y,et al.A green-cot-yledon/stay-green mutant exemplies the ancient whole-genome duplications in soybean[J].Plant Cell Physiol,2014,55(10):1763-1771.
[7] YANG K,JEONG N,MOON J K,et al.Genetic analysis of genes controllingnatural variation ofseed coat and flower colors in soybean[J].J Hered,2010,101(6):757-768.
[8]宋健,郭勇,邱丽娟.大豆种皮色相关基因定位[C]//第二届植物生物技术与现代农业论坛.武汉:武汉大学,2013.
[9]白志元,张瑞军,雷梦林,等.不同株系制种对大豆杂交种农艺性状的影响[J].山西农业科学,2016,44(8):1055-1058,1072.
[10]郭数进,杨凯敏,霍瑾,等.大豆不同性状与产量的相关性及主成分分析[J].山西农业科学,2015,43(5):505-508.
[11]王宏勇,郭数进,李贵全.晋旱125×(昔野×501)杂交后代产量与主要农艺性状的相关性及主成分分析[J].天津农业科学,2017,23(3):76-80.
[12]刘斌美,叶亚峰,章忠贵,等.一个籼稻脆性突变体的生物学特性及基因定位研究 [J].植物遗传资源学报,2011,12(5):259-264.
[13]杨凯敏,李贵全,郭数进,等.60Co诱变大豆后代表型性状的研究[J].天津农业科学,2014,20(10):5-9.
[14]杨万明,王敏,张谨华,等.不同百粒质量山西野生大豆生物学性状比较[J].山西农业科学,2016,44(7):927-931,937.
[15]周永航.大豆滞绿(Stay-green)突变体在辐射诱变育种中的应用研究[D].太谷:山西农业大学,2016.
[16]田风霞,惠振,王国坤,等.植物体内的叶绿素降解与滞绿突变体[J].植物生理学通讯,2010,46(5):505-511.
[17]于绍轩,韩粉霞,孙君明,等.空间环境对大豆主要农艺性状及蛋白品质的诱变效应[J].核农学报,2010,24(3):453-459.
[18] MANJAYA J G,SUSEELAN K N,GOPALAKRISHNA T,et al.Radiation induced variability of seed storage proteins in soybean(Glycine max (L.)Merrill)[J].Food Chemistry,2007,100(4):1324-1327.
[19]薛树鹏,郭蓓,杨柳,等.大豆‘科丰14’诱变系主要农艺性状和品质性状变异分析[J].农学学报,2014,4(3):1-6.
