生长环境与株龄对凤丹当年生果枝的生物量分配的影响
2018-04-10马菡泽汪成忠范国祯胡永红宋志平
马菡泽 汪成忠,2, 李 鼎 范国祯 胡永红 宋志平*
(1.生物多样性与生态工程教育部重点实验室,复旦大学生物多样性研究所,上海 200438; 2.苏州农业职业技术学院,苏州 215008; 3.上海辰山植物园,中国科学院上海辰山植物科学研究中心,上海市资源植物功能基因组学重点实验室,上海 201602)
植物在生长发育过程中,需要在不同器官间对内在资源(生物量)进行有效分配,以保障各生活史功能,即资源分配(resource allocation,也称为生物量分配)[1]。资源分配研究不仅对了解植物的生活史对策和生态适应性具有重要意义,也是解析作物产量形成机制的基础。资源分配受物种、环境、年龄及个体发育阶段等多种因素的影响[1~2]。比如,乔木、灌木和草本的资源分配式样差异明显,且受环境影响时采取分配策略也不一样[3~5]。大量研究表明,环境因子显著影响资源分配[1,6],当限制因子来自地上时,植物会增加茎叶的生物量,而当限制因子来自地下时,植物会增加根的生物量。近来有研究表明,看似是环境导致生物量分配变化,实际可能是因为个体大小变化所致[2],植物在个体较小时将同化生物量较多地分配到营养生长当中,当个体大小超过一定“阈值”时则将较多生物量用于繁殖生长[7],即异速生长模式[8~10]。个体大小与年龄相关。许多研究都表明多年生木本植物随株龄增大其地上生物量呈增加的趋势[11~14],这些研究多聚焦于多年生乔木物种,较少关注灌木。
无论乔木或灌木,多年生习性使得其资源分配规律复杂。这是因为,这些植物在资源利用上除了要保障当年的生长、繁殖,还必须满足来年的成活需求,其生物量分配在储藏(生存)、生长、繁殖器官之间存在权衡(trade-off),表现为源—库(source-sink)动态变化[2]。对于多年生植物,对整株取样来测定各组分的生物量在操作上也比较困难,替代办法是以当年生小枝(twig)为代表。小枝是多年生植物的构件中最活跃部分,叶片和果实大都长在其上,与老的枝条和茎相比,当年生小枝因次生维管组织相对较少,对营养元素、水和碳水化合物的分配更灵活[15],因而与较老枝条、甚至整株植物的生物量分配相比,当年生枝条的生物量分配最能反映植物的生活史特征[9,16]。
一些传统木本油料作物的生物量分配与产量关系研究相对较多,比如油棕[17]、油橄榄[18]、核桃[19~22]、油茶[23~24]等。这些研究总体上显示,生物量分配,特别是与产量密切相关的繁殖分配与作物明显受生长环境中的水分、营养和光照条件的影响。比如,核桃的座果数、平均果重和果枝率是核桃的主要产量构成要素,也是核桃丰产性的主要指标[19~20],采取控制水、肥和进行修剪等措施可提高核桃的结果量、坐果率及果实的干鲜重,增加繁殖分配,从而实现增产[21~22]。然而,株龄对木本油料作物的繁殖分配的效应较少关注。
油用牡丹是新型木本油料作物,近几年种植面积已快速扩大到6.67万公顷以上,但当前缺少高产品种和高产栽培措施,以至于油用牡丹籽粒单产一直不高[25],其原因在于,我们对油用牡丹的产量构成要素及产量形成机制缺乏了解。目前仅有少量研究探讨了油用牡丹的产量构成问题。比如,陈慧玲等[26]比较不同海拔种植的紫斑牡丹(Paeoniarockii)的单株果荚数、种子产量、百粒重、种子含油率等产量性状的表现,认为1 200~2 600 m是紫斑牡丹的最适海拔。赵玉如等[27]分析了不同花瓣类型牡丹的籽粒数量,发现5~7瓣类型对产量贡献最大,建议栽培管理中应重点关注5~7瓣的牡丹。凤丹是以杨山牡丹(Paeoniaostii)为原种形成的栽培类群,主要分布于我国黄河和长江中下游地区[28],具有结实率高、耐湿热、适应性强、病虫害少等特点,在安徽、河南、山东、陕西、四川等地有大面积种植,是油用牡丹主要栽培类型之一,在油用牡丹产业中具有举足轻重的地位。凤丹是多年生小灌木,主要以种子繁殖,从种子萌发到开花结实需要3~5年。凤丹也可以通过根状茎和根出条的方式进行无性繁殖。因而,凤丹整株用于繁殖和营养生长的生物量较难界定,但当年生果枝的营养与繁殖部分界限清晰,利于资源分配分析。而且,凤丹单株产量主要由当年生果枝数量和果枝的籽粒产量决定,果枝的繁殖分配与产量直接相关。此外,凤丹果枝具有“枯枝退稍”现象,即当年生果枝生长量的2/3会木质化,待到其年生长周期结束后会逐渐枯死,但不完全凋落[29],这种现象很可能与果枝的资源分配有关。因此,本研究以凤丹当年生果枝(小枝)为检测对象,通过比较不同年龄的种群以及不同产地的同龄的种群的小枝各组分的生物量分配差异,明确果枝资源分配规律,分析株龄及栽培环境对其的影响,为探讨凤丹产量形成机制奠定基础。
1 材料与方法
1.1 材料来源
为了了解株龄对资源分配的影响,我们对来自安徽铜陵和上海的4龄、6龄、8龄凤丹种群进行了取样调查。铜陵种群生长在安徽省铜陵市凤凰山牡丹园,该园位于安徽省南部,是丹皮的道地产区,园内种植有株龄不同(2~9龄)的凤丹种群。上海种群由来自凤凰山的2龄、4龄、6龄的凤丹材料移栽到复旦大学江湾实验基地,经过两年生长形成的4龄、6龄、8龄种群。为了比较不同栽培环境下凤丹资源分配的差异,我们同时调查了8地区的6龄种群,包括:安徽省铜陵市(TL)和亳州市(BZ),重庆市(CQ),湖南省邵阳市(SY),山东省菏泽市(HZ),河南省洛阳市(LY),陕西省咸阳市(XY)和上海市(SH)(表1)。
表1采样位点地理位置及环境信息
Table1Theenvironmentalinformationofsamplingpopulations
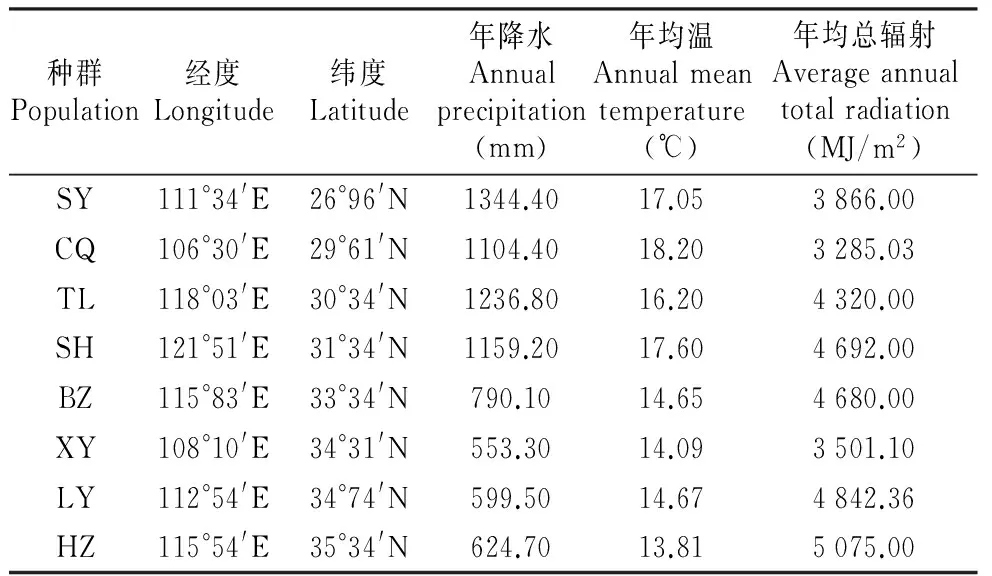
种群Population经度Longitude纬度Latitude年降水Annualprecipitation(mm)年均温Annualmeantemperature(℃)年均总辐射Averageannualtotalradiation(MJ/m2)SY111°34'E26°96'N1344.4017.053866.00CQ106°30'E29°61'N1104.4018.203285.03TL118°03'E30°34'N1236.8016.204320.00SH121°51'E31°34'N1159.2017.604692.00BZ115°83'E33°34'N790.1014.654680.00XY108°10'E34°31'N553.3014.093501.10LY112°54'E34°74'N599.5014.674842.36HZ115°54'E35°34'N624.7013.815075.00
1.2 样品采集及指标测定
在凤丹的果实成熟期(2016年7月),我们分别在各种群中随机选取大小与长势相近的凤丹15~30株作为样株,先统计样株的果枝数,然后从每个样株上选取1个果枝,从基部(与老茎连接处)截取,将果枝带回实验室检测。在室内先测定每个果枝的基茎和枝长,统计种子数,然后将果荚、种子、叶片、茎(包括叶柄)分开,置于80℃处理至恒重后称重。通过检测和计算,我们获得以下三类指标,第一类小枝形态指标:分枝数(twig number,TN);小枝基茎(basic diameter,BD);小枝长(twig height,TH);第二类营养生物量及其分配:茎生物量(stem mass,SM);叶生物量(leaf mass,LM);营养生物量(vegetative mass,VM)=SM+LM;总生物量(total mass,TM)=SM+LM+FM;叶生物量比例(leaf mass ratio,LMR)=LM/TM*100%;茎生物量比例(stem mass ratio,SMR)=SM/TM*100%;第三类繁殖生物量及其分配:种子量(seed number,SN);种子重(seed weight,SW);果实生物量(fruit mass,FM)=果荚重+SW;百粒重(100seed weight,100seed)=SW/SN*100;果实生物量比例(fruit mass ratio,RMRf)=FM/TM*100%;种子生物量比例(seed mass ratio,SM/TM*100%,RMRs)=SW/TM*100%。
1.3 数据分析
数据分析之前,我们对不符合正态分布数据进行log或squrt转换。用单因素方差分析(one-way ANOVA)对来自不同地区的6龄种群进行分析,以检验环境效应。采用双因素方差分析(two-way ANOVA)对铜陵和上海两地4龄、6龄、8龄种群的检测指标进行了分析,以检验株龄、环境效应。如果方差检验显示存在效应,进一步采用多重比较来检验两两之间的差异。我们还采用相关分析方法分析了检测指标之间的相关性。为了讨论环境因子对小枝生物量分配的影响,我们对年均总辐射、降水、年均温和纬度这四个环境因子和测量指标也进行了回归分析。这些分析都由软件spss 16.0完成。
此外,我们用异速生长方程来描述6龄种群小枝内不同功能器官之间生物量分配关系。
Y=λXβ
(1)
对方程进行对数转换后构成Y和X的线性关系:
(2)
式中,X和Y分别代表所考察的不同功能器官生物量;λ代表属性关系的截距(Slope);β为斜率,代表了相对生长的指数。
当β=1时为等速生长,β大于或小于1时为异速生长[30]。异速生长方程的参数估计采用标准化主轴估计(Standardized major axis estimation,SMA)方法[31],由软件SMATR(Version 2.0)来计算完成。
2 结果
2.1 不同地点6龄凤丹种群的小枝性状表现
One-way ANOVA对8个来自不同地点的6龄凤丹种群的小枝性状表现的分析结果显示,除了指标百粒重(100 seed)和基径(BD)在不同种群间没有显著差异外(100 seed的平均值为30.41±7.24 g,基径均值为8.54±1.45),其他指标都明显表现出地点效应(表2)。总的来说,洛阳(LY)种群单株果枝数最多而果枝最短,铜陵(TL)种群果枝数最少,重庆(CQ)和邵阳(SY)果枝最长;上海(SH)种群的小枝较粗,而其他种群的小枝基径没有明显差异;小枝茎生物量最大的为重庆和邵阳种群,最小的为洛阳种群;邵阳种群小枝的叶生物远大于其他种群,而其他种群之间的小枝叶生物没有明显差异;小枝果实生物量最大的为洛阳种群,最小的则为上海种群;种子数和重量最多为LY种群,最少为SH种群;小枝总重较大的为CQ和SY种群,而HZ、SH、TL种群的小枝总重较小(表3)。在生物量分配方面,SY种群小枝的营养分配(SMR、LMR)较多而繁殖分配(RMRf、RMRs)最少,LY种群繁殖分配最多而营养分配最少(表3)。
2.2 不同株龄凤丹种群的小枝性状表现
双因素方差分析(two-way ANOVA)分析结果表明,株龄仅在百粒重和分枝数(TN)两个指标上有明显效应;除基径、茎生物量(SM)、总生物量(TM)和百粒重等指标上没有明显地点效应外,其它指标都受地点的影响;叶生物量(LM)、百粒重、叶生物量比例(LMR)、茎生物量比例(SMR)、果实生物量比例(RMRf)和种子生物量比例(RMRs)等指标还受株龄与地点之间交互作用的影响(表2,图1)。
表2凤丹小枝性状表现的地点、株龄效应方差分析的F值
Table2F-valuesofANOVAfortwigtraitperformanceofP.ostii
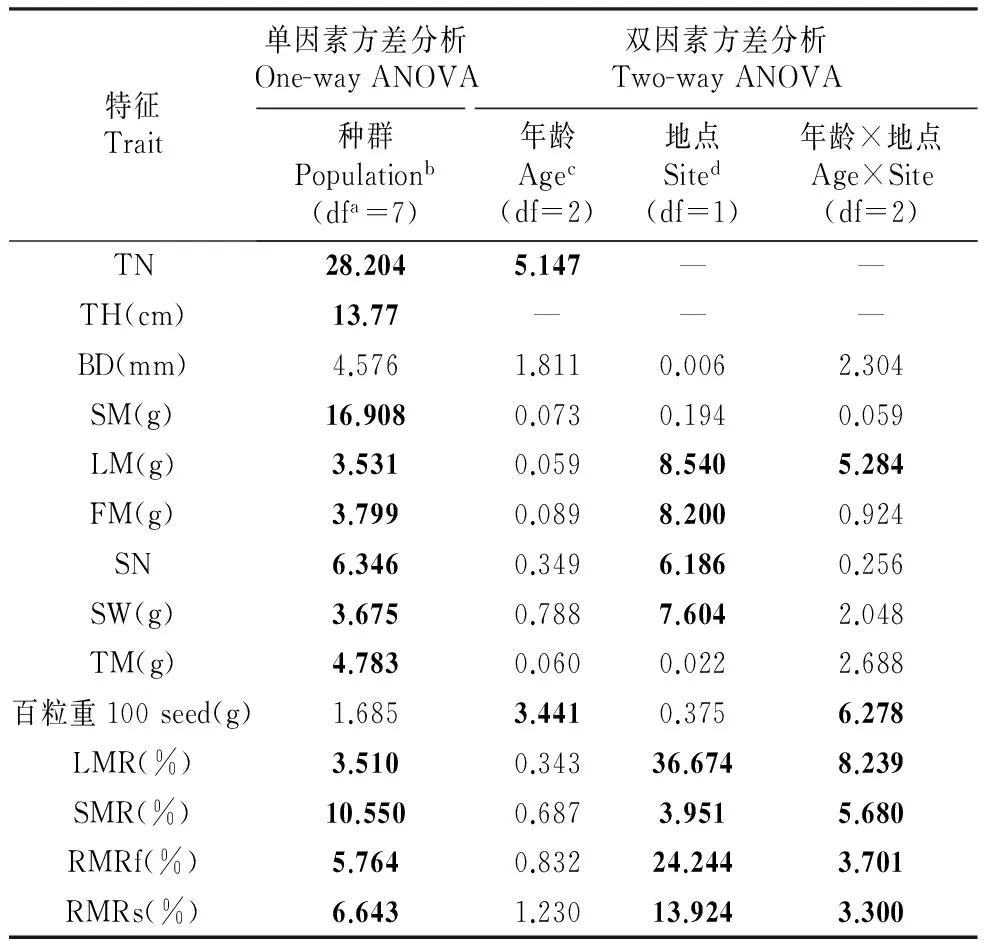
特征Trait单因素方差分析One-wayANOVA双因素方差分析Two-wayANOVA种群Populationb(dfa=7)年龄Agec(df=2)地点Sited(df=1)年龄×地点Age×Site(df=2)TN28.2045.147——TH(cm)13.77———BD(mm)4.5761.8110.0062.304SM(g)16.9080.0730.1940.059LM(g)3.5310.0598.5405.284FM(g)3.7990.0898.2000.924SN6.3460.3496.1860.256SW(g)3.6750.7887.6042.048TM(g)4.7830.0600.0222.688百粒重100seed(g)1.6853.4410.3756.278LMR(%)3.5100.34336.6748.239SMR(%)10.5500.6873.9515.680RMRf(%)5.7640.83224.2443.701RMRs(%)6.6431.23013.9243.300
注:a.自由度;b.来自上海(SH)、铜陵(TL)、亳州(BZ)、菏泽(HZ)、洛阳(LY)、咸阳(XY)、重庆(CQ)、邵阳(SY)的8个6龄种群;c. 4龄,6龄,8龄三个龄级;d.包括了上海和铜陵两个地区P<0.05加黑标出
Note: a. Freedom degree; b. Includes the 6-year old population from SH,TL,BZ,HZ,LY,XY,CQ and SY; c. Includes the 4,6 and 8-year old populations; d. Includes Shanghai and TonglingP<0.05 is shown with bold.

表3 不同地区6龄凤丹种群小枝生物量指标平均值
注:角标不同代表不同种群间差异显著,两两比较检验方法为S-N-K,差异显著检验水平P=0.05。
Note:Different lowercase letters denote significant differences among populations according to the Student-Newman-Keuls test atP=0.05.
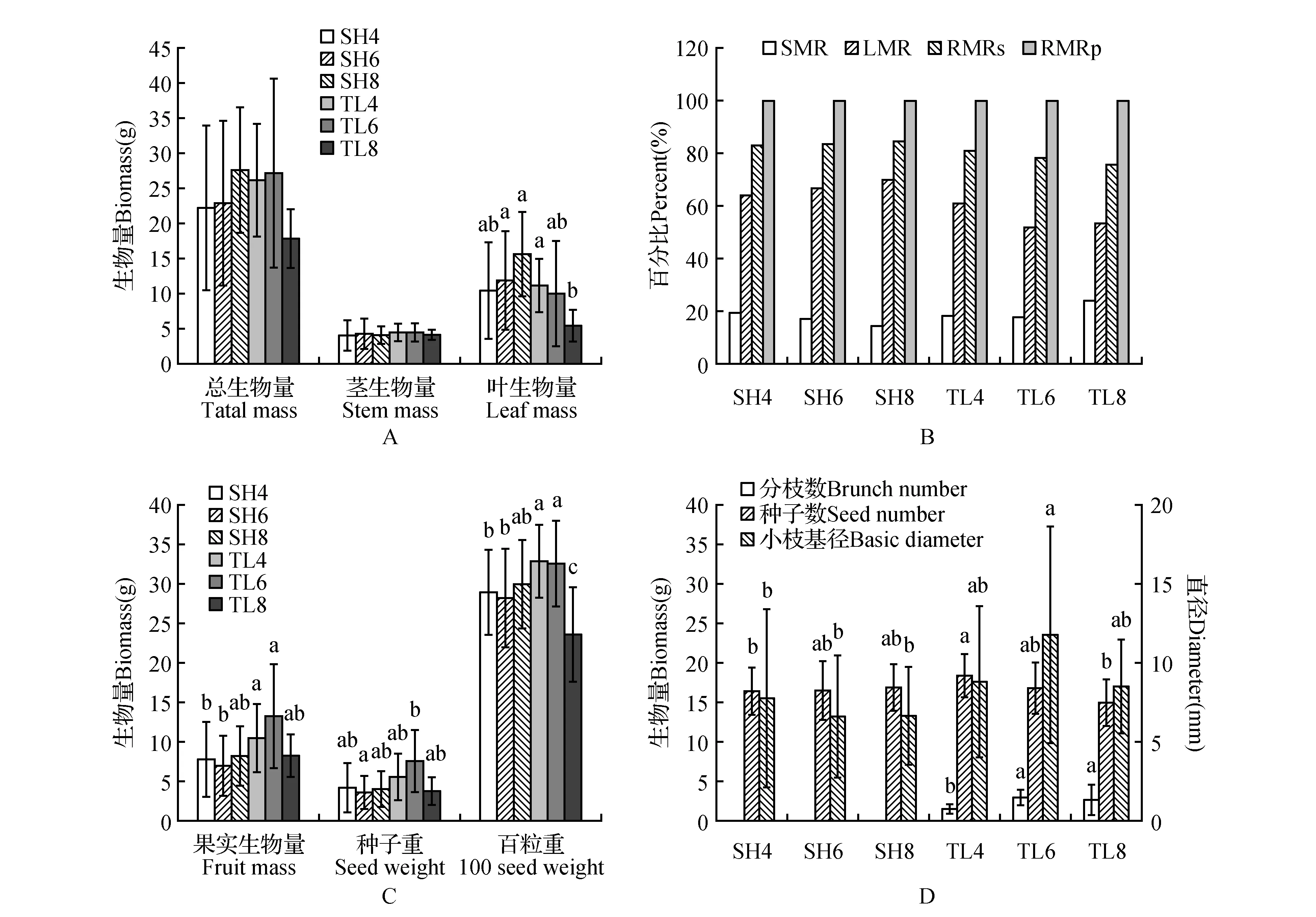
图1 不同株龄当年生小枝的性状表现 SH4、SH6、SH8,上海4龄、6龄、8龄种群;TL4、TL6、TL8,铜陵4龄、6龄、8龄种群Fig.1 Twig trait performance in different age populations of P.ostii SH4,SH6,SH8,4,6,8 year old populations from Shanghai; TL4,TL6,TL8,4,6,8 year populations from Tongling of Anhui Province
既不表现为株龄效应也不表现为地点效应指标基径、茎生物量、总生物量的变化范围分别为4.73~12.82 mm、0.71~9.50 g、4.20~55.5 g。分枝数随着株龄增大而增加(1.43~2.60,图1A)。8龄种群的小枝100seed明显低于6龄和4龄种群(图1A)。上海种群的叶生物量总体上大于铜陵种群,其中铜陵8龄种群(TL8)的叶生物量最小(图1B);而果实重指标总体上铜陵种群大于上海种群,其中TL6果实重最大(13.23±6.64 g),种子重指标表现与此类似(7.46±3.99 g)(图1C);相应地,铜陵种群的繁殖分配比例(RMRf和RMRs)高于上海种群,TL6种群最高(图1D)。
2.3 性状指标之间的相关性及繁殖分配
Pearson相关分析结果显示,除了种子数(SN)外,几乎所有指标之间都存在一定的相关性(表4),其中,基径与所有生物量指标以及叶生物量比例之间都成正相关关系,与繁殖分配呈负相关;而枝长(TH)与这些指标之间相关性明显下降。进一步的回归分析基径大小也可以较好地用来表征小枝的大小(图2)。表4还显示,小枝的总生物量与茎、叶、果实、种子生物量都显著正相关,果实与种子生物量高度正相关;繁殖分配与基径、枝长、茎生物量、叶生物量、总生物量、茎分配、叶分配等指标都显著负相关,而与种子数、种子重(SW)、果实重(FW)正相关。对各个种群分别进行相关性分析则显示,上述相关关系在不同种群之间存在明显差异(数据未提供,作者保持备查),例如:亳州(BZ)、重庆(CQ)、邵阳(SY)种群的果实生物量(FM)与总生物量(TM)相关性不显著,而其他种群都显著。
繁殖生物量(FM、SW)与总生物量(TM)、以及与营养生物量(VM=TM-FM或TM-SW)之间的异速生长模拟分析显示,邵阳种群的繁殖与营养生长之间(FM、SW vs. TM或VM),以及重庆(CQ)、菏泽(HZ)、洛阳(LY)、亳州(BZ)种群的果实生物量(FM)与营养生物量(VM)之间的关系不符合异速模型(P>0.05),其他指标之间的关系都可以用异速模型描述(表5)。表5显示,SH种群的营养生长(VM)与种子生物量(SW)之间回归线的slop>1,表明存在异速关系;其他种群的繁殖(FB、SW)与总生物量(TB)以及与营养(TM)之间为等速生长(slop=1),但回归方程的截距在种群间变化较大(-1.305 5~0.038)。
表型指标与环境因子的回归分析显示,随着纬度增加、光辐射增加,小枝总生物量、茎生物量与茎分配下降,而小枝叶生物量随降水增多而上升(图3),其他指标与环境因子之间没有显著相关性。

表4 凤丹当年生小枝形态与生物量指标间的Pearson相关系数(r)
*P<0.05,**P<0.001
表56龄凤丹种群的小枝繁殖与营养生物量的异速关系
Table5Allometricrelationshipsbetweenreproductiveandvegetativebiomassofthetwigof6-yearoldFengdanpopulation
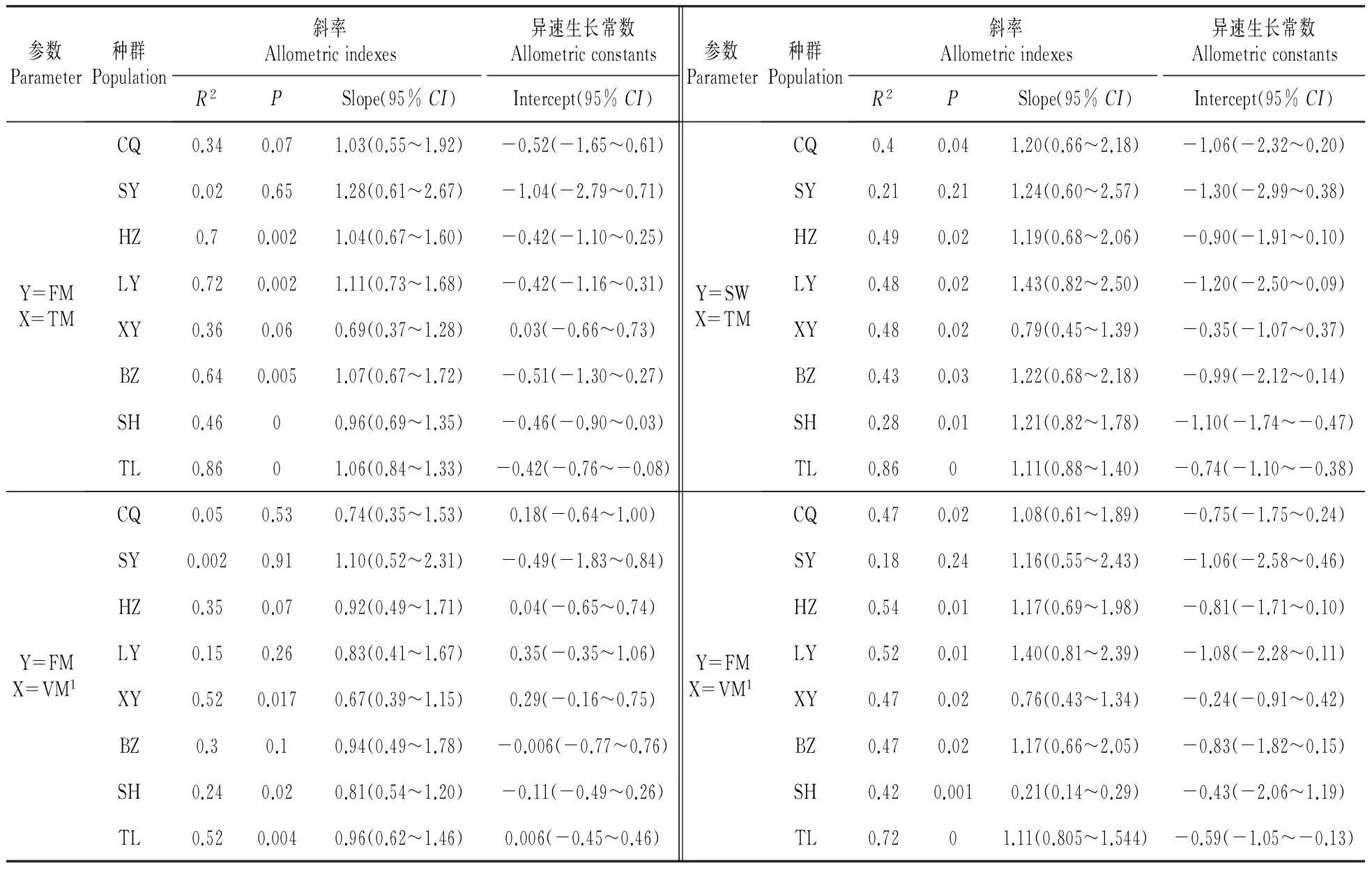
参数Parameter种群Population斜率Allometricindexes异速生长常数AllometricconstantsR2PSlope(95%CI)Intercept(95%CI)参数Parameter种群Population斜率Allometricindexes异速生长常数AllometricconstantsR2PSlope(95%CI)Intercept(95%CI)Y=FMX=TMCQ0.340.071.03(0.55~1.92)-0.52(-1.65~0.61)SY0.020.651.28(0.61~2.67)-1.04(-2.79~0.71)HZ0.70.0021.04(0.67~1.60)-0.42(-1.10~0.25)LY0.720.0021.11(0.73~1.68)-0.42(-1.16~0.31)XY0.360.060.69(0.37~1.28)0.03(-0.66~0.73)BZ0.640.0051.07(0.67~1.72)-0.51(-1.30~0.27)SH0.4600.96(0.69~1.35)-0.46(-0.90~0.03)TL0.8601.06(0.84~1.33)-0.42(-0.76~-0.08)Y=SWX=TMCQ0.40.041.20(0.66~2.18)-1.06(-2.32~0.20)SY0.210.211.24(0.60~2.57)-1.30(-2.99~0.38)HZ0.490.021.19(0.68~2.06)-0.90(-1.91~0.10)LY0.480.021.43(0.82~2.50)-1.20(-2.50~0.09)XY0.480.020.79(0.45~1.39)-0.35(-1.07~0.37)BZ0.430.031.22(0.68~2.18)-0.99(-2.12~0.14)SH0.280.011.21(0.82~1.78)-1.10(-1.74~-0.47)TL0.8601.11(0.88~1.40)-0.74(-1.10~-0.38)Y=FMX=VM1CQ0.050.530.74(0.35~1.53)0.18(-0.64~1.00)SY0.0020.911.10(0.52~2.31)-0.49(-1.83~0.84)HZ0.350.070.92(0.49~1.71)0.04(-0.65~0.74)LY0.150.260.83(0.41~1.67)0.35(-0.35~1.06)XY0.520.0170.67(0.39~1.15)0.29(-0.16~0.75)BZ0.30.10.94(0.49~1.78)-0.006(-0.77~0.76)SH0.240.020.81(0.54~1.20)-0.11(-0.49~0.26)TL0.520.0040.96(0.62~1.46)0.006(-0.45~0.46)Y=FMX=VM1CQ0.470.021.08(0.61~1.89)-0.75(-1.75~0.24)SY0.180.241.16(0.55~2.43)-1.06(-2.58~0.46)HZ0.540.011.17(0.69~1.98)-0.81(-1.71~0.10)LY0.520.011.40(0.81~2.39)-1.08(-2.28~0.11)XY0.470.020.76(0.43~1.34)-0.24(-0.91~0.42)BZ0.470.021.17(0.66~2.05)-0.83(-1.82~0.15)SH0.420.0010.21(0.14~0.29)-0.43(-2.06~1.19)TL0.7201.11(0.805~1.544)-0.59(-1.05~-0.13)
VM1=TM-FM; VM2=TM-SW
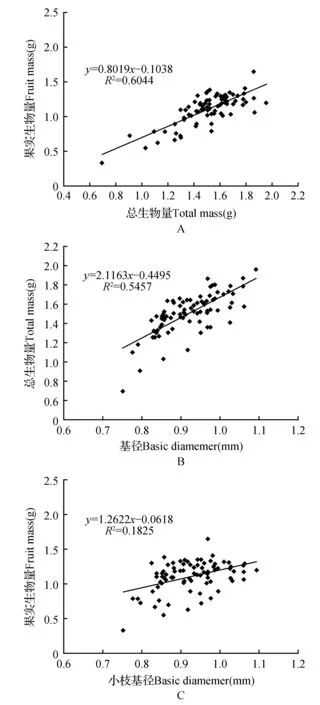
图2 小枝大小(基径BD或总生物量TM)与繁殖生物量(FM)的线性回归关系Fig.2 Log10-log10 linear regression plot of twig size(BD or TM) and reproductive biomass(FM)

图3 环境因子与生物量特征线性回归关系 a.茎生物量及分配(SM、SMR)与年均光辐射;b.叶生物量分配(LMR)与年均降水量;c.叶生物量(LM)及营养生物量(VM)与纬度Fig.3 Linear regression plot of environment variables and biomass characteristics a. SMR and SM vs average optical radiation; b. LMR vs average annual precipitation; c. VM and LM vs latitude
3 讨论
3.1 凤丹小枝的资源分配的影响因素
3.1.1株龄效应
通常认为,年龄(株龄)对多年生木本植物的生物量分配有显著效应,随着株龄增大,地上生物量比例增大[14,32~34],繁殖生物量绝对量增加但相对比例下降[35~37],即所谓地上与地下部分、营养与繁殖结构的异速生长关系。本研究显示,株龄对凤丹当年生果枝的营养与繁殖分配没有明显效应,不同株龄凤丹的小枝的大小、各器官的生物量比例没有显著差异(表2,图1)。出现这种现象的原因可能是,当年生小枝除了在生长发育初期(萌芽)需要消耗其他结构储藏的资源外,其他阶段基本上依赖自身生长过程中叶片积累的同化物,因而不同株龄凤丹的当年生小枝的大小没有差异(基径、总生物量都没有差异(图1D))。同时,基于营养(资源)就近分配原则,小枝繁殖结构的生长发育主要依赖自身同化物,几乎不需要其它构件提供资源,因此当年生小枝的营养与繁殖分配也与年龄无关。此外,理论与实验研究表明,植物的繁殖与个体大小相关,只有达到一定大小的个体才能从营养生长转向繁殖生长,即繁殖的个体大小阈值,而这种阈值通常为物种属性,与年龄无关[7,38]。由此推测,只有达到一定大小的凤丹小枝(实际上由芽的大小决定)才能开花结实,不同株龄果枝大小没有差异,相应地营养与繁殖分配也没有差异。凤丹果枝有“枯枝退稍”现象,而营养枝则没有。可能正是因为当年生果枝的繁殖生长完全依赖小枝自身积累的同化物,导致小枝资源枯竭,使得果枝上大部分的结构无法越冬存活,才出现“枯枝退稍”现象。
尽管本研究没有检测到株龄对当年生果枝生物量分配有明显效应,但我们发现不同株龄凤丹的当年果枝数量差异明显,4龄植株的果枝数(1.43±0.64)显著低于6龄和8龄植株(6龄=2.85±1.04和8龄=2.60±1.95),而不同株龄单个果枝的繁殖量(种子重)没有差异,因而从整株水平来看,株龄效应明显。此外,我们采取破坏性取样分析不同株龄凤丹整株的生物量分配结果也显示,不同株龄凤丹植株的大小及各部分生物量分配(包括繁殖分配)比例差异显著(马菡泽等未发表数据)。因此,本研究并不否定年龄对多年生木本植物的生物量分配有显著效应的观点[12~14]。
另外,我们还发现在百粒重指标上存在年龄效应(表2),8龄种群的百粒重明显小于6龄和4龄种群(图1)。一般认为,单个果实内种子数量与百粒重之间呈现负相关关系,即种子数越多,单个种子大小越小,即对有限资源利用上的权衡[39]。然而,本研究并未发现这种趋势。其原因可能在于,本研究百粒重是基于由单个果实种子数和单个果实种子重量计算而来,8龄植株有少数果枝上结两个果实,而4龄、6龄果枝上通常只有一个果实,这导致8龄小枝的果荚资源消耗比例增加,使得在总的繁殖投入相对稳定、种子量变化不大的情况下,单粒种子质量下降,仍然可以看成是一种权衡。如果这种式样是真实的,在实际生产中,我们可以通过去掉小枝上的次花来减少繁殖资源的消耗,间接提高经济产量(籽粒越大,出油率越高)。
3.1.2环境效应
本研究显示,不同地理种群的果枝大小及其内在生物量分配存在明显差异。种群之间的表型差异既可以由环境差异所致,也有可能是遗传分化的反映,或者是两者共同作用的结果[2]。现有的遗传学研究表明,我国各地栽培的凤丹(即杨山牡丹)种群遗传上没有明显分化[28]。特别是,本研究中上海种群与铜陵种群具有相同来源,遗传背景相同,但它们之间也表现出明显差异(表2),强烈暗示我们检测到种群之间的资源分配差异是环境效应的结果,支持环境对资源分配有显著影响的观点[1]。
在所调查的8个6龄种群中,部分种群(如重庆、邵阳、上海)小枝将较多的生物量分配到了营养结构中,而另一些种群(如洛阳、铜陵等)则将较多的生物量分配到了繁殖结构中。这种变化反映了不同地区环境条件的差异对营养与繁殖生长的影响和植物对生长环境变化的响应。当环境条件有利时,如水肥条件优越时,植物倾向于营养生长,对于农作物即所谓“疯长”现象;当环境条件下降时,植物提高繁殖投入,以保证繁殖成功。进一步分析显示种群果枝生物量分配明显当地气候变量有关,例如,茎分配随光辐射增加而下降,但随降水增加而增加;而叶及营养结构生物量与纬度负相关(图3)。Poorter等(2012)在对850个物种生物量分配随环境因子改变的Meta分析中指出,环境因子会显著影响植物资源分配模式,当限制资源为光时植物会增加茎和叶生物量分配这与我们的结果一致。我们的结果表明,年均光辐射较强的地区,如洛阳,其果实生物量远高于其他地区,因此凤丹在种植时因首先考虑开阔的环境而非郁闭的环境,这一点在凤丹的近缘种滇牡丹(Paeoniadelavayi)中也得到了证实[40]。
除了上述环境因子外,土壤肥力和栽培密度也是影响植物繁殖分配地上生物量总量的重要因素。比如,玉米和大豆的繁殖生物量会随着密度的增加呈现先升后降的趋势[7];黄帚橐吾(Ligulariavirgaurea)则随着密度增加呈现下降趋势[41]。钝裂银莲花(Anemoneobtusiloba)随着土壤氮浓度的增加而繁殖生物量上升[42],苘麻(Abutilontheophrasti)等植物随着N浓度上升而地上生物量上升[6]。因此,我们推测各地凤丹种群的繁殖生物量分配差异与当地的土壤肥力和栽培密度不同有关。是否如此,需要设计试验来深入探讨。
3.2 凤丹小枝的资源分配式样
通常认为,植物的繁殖与营养结构存在异速生长关系,而且这种关系为物种属性,同一物种不同种群表现相似[43]。近来也有研究显示,同一物种不同种群会因生长环境不同而发生异速关系变化,比如异速生长方程的斜率(slope)变化,>1或<1[44]。异速方程斜率反映了随着个体大小增加,营养生长转向繁殖的效率,即相对生长率[45]。本研究显示,虽然总体上凤丹种群(邵阳、洛阳种群例外)小枝的繁殖与营养生长的关系可以用异速生长方程来描述,但斜率都没有明显偏离1(SH种群的SW vs. VM例外)(表5),提示繁殖与营养结构为等速生长。小枝不同结构之间等速生长关系在杜鹃属(Rhododendron)中也发现过[10]。
营养与繁殖异速生长方程的斜率和截距在种群间的变化反映了种群对生长环境响应。例如,在个体大小相同情况下,马先蒿(Pedicularis)高海拔种群的营养与繁殖异速生长方程的截距较大,反映出高海种群的繁殖效率更高[44],这实际上是该类群在繁殖策略上的适应性表现[46]。本研究发现营养与繁殖异速生长方程的斜率在凤丹种群之间没有差异,但截距变化较大(表5),这可能反映了生长环境的影响。表形性状与环境因子的相关性分析结果(图3)也支持这个结论。
作物的产量通常有单株产量和单位面积的植株数量决定。本研究显示不同株龄凤丹的单个果枝的生物量及分配没有差异,但果枝数量差异明显。这表明,在栽培密度不变条件下,株龄越高,单株果枝数越大,单株产量就越大,总产量也就越高。因而,在油用牡丹的生产上应采取措施提高单株果枝数,如采用高龄植株,或通过株型修整促进分枝等。此外,本研究显示生长环境对凤丹的果枝的总生物量及分配有显著影响,提示优化生长环境,如光照和水肥条件等,可有效提高凤丹的籽粒产量。
1.Poorter H,Niklas K J,Reich P B,et al.Biomass allocation to leaves,stems and roots:meta-analyses of interspecific variation and environmental control[J].New Phytologist,2011,193(1):30-50.
2.Weiner J.Allocation,plasticity and allometry in plants[J].Perspectives in Plant Ecology,Evolution and Systematics,2004,6(4):207-215.
3.高杰,郭子健,刘艳红.北京松山不同龄级天然油松林生物量分配格局及其影响因子[J].生态学杂志,2016,35(6):1475-1480.
Gao J,Guo Z J,Liu Y H.Biomass allocation pattern and its influencing factors across natural Chinese pine forests of different ages in Songshan,Beijing[J].Chinese Journal of Ecology,2016,35(6):1475-1480.
4.姚红,谭敦炎.胡卢巴属4种短命植物的生长特点及繁殖分配动态[J].新疆农业大学学报,2005,28(3):26-29.
Yao H,Tan D Y.The growth characteristics and the reproductive allocation dynamics in four ephemeral species ofTrigonella[J].Journal of Xinjiang Agricultural University,2005,28(3):26-29.
5.刘贵周,蔡传涛,罗媛,等.两种萝芙木生物量分配与估测模型研究[J].云南植物研究,2009,31(1):63-66.
Liu G Z,Cai C T,Zhou Y,et al.Biomass allocation and biomass estimation models ofRauvolfiavomitoriaandRauvolfiaverticillata(Apocynaceae)[J].Acta Botanica Yunnanica,2009,31(1):63-66.
6.Mcconnaughay K D M,Coleman J S.Biomass allocation in plants:ontogeny or optimality? A test along three resource gradients[J].Ecology,1999,80(8):2581-2593.
7.Weiner J,Lovett D J,Lovett D L.The influence of competition on plant reproduction[M].//Lovett Doust J,Lovett Doust L.Plant Reproductive Ecology:Patterns and Strategies.New York:Oxford University Press,1988:228-245.
8.祝介东,孟婷婷,倪健,等.不同气候带间成熟林植物叶性状间异速生长关系随功能型的变异[J].植物生态学报2011,35(7):687-698.
Zhu J D,Meng T T,NI J,et al.Within-leaf allometric relationships of mature forests in different bioclimatic zones vary with plant functional types[J].Chinese Journal of Plant Ecology,2011,35(7):687-698.
9.杨冬梅,毛林灿,彭国全.常绿和落叶阔叶木本植物小枝内生物量分配关系研究:异速生长分析[J].植物研究,2011,31(4):472-477.
Yang D M,Mao L C,Peng G Q.Within-twig biomass allocation in evergreen and deciduous broad-leaved species:allometric scaling analyses[J].Bulletin of Botanical Research,2011,31(4):472-477.
10.李亚男,杨冬梅,孙书存,等.杜鹃花属植物小枝大小对小枝生物量分配及叶面积支持效率的影响:异速生长分析[J].植物生态学报,2008,32(5):1175-1183.
Li Y N,Yang D M,Sun S C,et al.Effects of twig size on biomass allocation within twigs and on lamina area supporting efficiency inRholdlden:allometric scaling analyses[J].Journal of Plant Ecology,2008,32(5):1175-1183.
11.Gu D,Yang D R,Compton S G,et al.Age at pollination modifies relative male and female reproductive success in a monoecious fig tree[J].Symbiosis,2012,57(2):73-81.
12.López F,Fungairio S,De Las Heras P,et al.Age changes in the vegetative vs.reproductive allocation by module demographic strategies in a perennial plant[J].Plant Ecology,2001,157(1):13-21.
13.Carbone M S,Czimczik C I,Keenan T F,et al.Age,allocation and availability of nonstructural carbon in mature red maple trees[J].New Phytologist,2013,200(4):1145-1155.
14.Enright N J.Age,reproduction and biomass allocation inRhopalostylissapida(Nikau Palm)[J].Austral Journal of Ecology,2010,10(4):461-467.
15.Xiang S A,Wu N,Sun S C.Within-twig biomass allocation in subtropical evergreen broad-leaved species along an altitudinal gradient:allometric scaling analysis[J].Trees,2009,23(3):637-647.
16.Bonser S P,Aarssen L W.Meristem allocation:A new classification theory for adaptive strategies in herbaceous plants[J].Oikos,1996,77(2):347-352.
17.Pallas B,Mialet-Serra I,Rouan L,et al.Effect of source/sink ratios on yield components,growth dynamics and structural characteristics of oil palm(Elaeisguineensis) bunches[J].Tree Physiology,2013,33(4):409-424.
18.Cherbiy-Hoffmanna S U,Hallb A J,Rousseauxa M C.Fruit,yield,and vegetative growth responses to photosynthetically active radiation during oil synthesis in olive trees[J].Scientia Horticulturae,2013,150:110-116.
19.赵宝军,宫永红.早实核桃优良单株产量主要影响因子的相关分析及通径分析[J].中国果树,2013,(3):27-29.
Zhao B J,Gong Y H.The correlative analysis and pathway analysis of high productive early-fruiting walnut[J].China Fruit,2013,(3):27-29.
20.陈春芳,吴云华,余崇彪,等.早实核桃产量影响因子研究初报[J].湖北林业科技,2008(4):21-23.
Chen C F,Wu Y H,Yu C B,et al.Initial report on study of impact factor of early-fruit bearing production forJuglansregiaL.[J].Hubei Forestry Science and Technology,2008(4):21-23.
21.Martens S N,Ustin S L,Norman J M.Measurement of tree canopy architecture[J].International Journal of Remote Sensing,1991,12(7):1525-1545.
22.陈虹,董玉芝,朱小虎,等.新疆早实核桃品种测土配方施肥肥效试验初报[J].新疆农业科学,2010,47(8):1584-1589.
Chen H,Dong Y Z,Zhu X H,et al.Preliminary discussion on the effects of fertilizer experiment with “3414” design in precocious walnut[J].Xinjiang Agricultural Sciences,2010,47(8):1584-1589.
23.唐健,宋贤冲,曹继钊,等.不同产量油茶林树体结构及生物量分配差异[J].湖北农业科学,2013,52(12):2848-2850.
Tang J,Song X C,Cao J Z,et al.Different yield oil Camellia tree structure and differences of biomass allocation[J].Hubei Agricultural Sciences,2013,52(12):2848-2850.
24.康志雄,邹达明,王芷虔.用灰色系统理论分析油茶产量形成[J].浙江林业科技,1993,13(3):62-65.
Kang Z X,Zou D M,Wang Z J.Analysis on formation of teaoilCamelliaoutput by grey system theory[J].Zhejiang Forestry Science and Technology,1993,13(3):62-65.
25.李晓青,韩继刚,刘炤,等.不同地区凤丹经济性状及其籽油脂肪酸成分分析[J].粮食与油脂,2014,27(4):43-46.
Liu X Q,Han J G,Liu Z,et al.Economic characteristics investigation and seed oil fatty acid composition analysis ofPaeoniaostiiplants in different areas[J].Cereals & Oils,2014,27(4):43-46.
26.陈慧玲,李洪喜,张建华,等.‘保康紫斑’牡丹生长适应性及结籽性状[J].林业工程学报,2014,28(4):43-46.
Chen H L,Li H X,Zhang J H,et al.The growth adaptation and seed-bearing characteristics ofPaeoniarockii‘Baokang Ziban’ in Hubei Province[J].Journal of Forestry Engineering,2014,28(4):43-46.
27.赵玉如,于晓金,谷欣,等.油用牡丹产量构成初步观测及分析[J].山东林业科技,2014,(5):79-81.
Zhao Y R,Yu X J,Gu X,et al.The analysis and observation of yield components of oil peony[J].Journal of Shandong Forestry Science and Technology,2014(5):79-81.
28.王佳.杨山牡丹遗传多样性与江南牡丹品种资源研究[D].北京:北京林业大学,2009:72-129.
Wang J.Genetic diversity ofPaeoniaostiiand germplasm resources of tree peony cultivars form Chinese Jiangnan area[D].Beijing:Beijing Forestry University,2009:72-129.
29.李嘉珏.中国牡丹与芍药[M].北京:中国林业出版社,1999:120-157.
Li J Y.Chinese peony and Paeonia lactiflora[M].Beijing:China Forestry Publishing House,1999:120-157.
30.Harvey P H,Pagel M D.The comparative method in evolutionary biology[M].Oxford:Oxford University Press,1991:169-172.
31.Falster D S,Warton D I,Wright I J.User’s guide to SMATR:standardised major axis tests & routines version 2.0[Z].2006.
32.Samuelson L J,Stokes T A,Butnor J R,et al.Ecosystem carbon density and allocation across a chronosequence of longleaf pine forests[J].Ecological Applications,2017,27(1):244-259.
33.Yang X,Yang Z Y,Wang X Y,et al.Tree biomass allocation and it’s model additivity forCasuarinaequisetifoliaina tropical forest of Hainan Island,China[J].PLoS One,2016,11(3):e0151858.
34.Li H,Li C Y,Zha T S,et al.Patterns of biomass allocation in an age-sequence of secondaryPinusbungeanaforests in China[J].The Forestry Chronicle,2014,90(2):169-176.
35.赵方,杨永平.中华山蓼不同海拔居群的繁殖分配研究[J].植物分类学报,2008,46(6):830-835.
Zhao F,Yang Y P.Reproductive allocation in a dioecious perennialOxyriasinensis(Polygonaceae) along altitudinal gradients[J].Journal of Systematics and Evolution,2008,46(6):830-835.
36.赵志刚,杜国祯,任青吉.5种毛茛科植物个体大小依赖的繁殖分配和性分配[J].植物生态学报,2004,28(1):9-16.
Zhao Z G,Du G Z,Ren Q J.Size-dependent reproduction and sex allocation in five species of Ranunculaceae[J].Chinese Journal of Plant Ecology,2004,28(1):9-16.
37.王一峰,刘启茜,裴泽宇,等.青藏高原3种风毛菊属植物的繁殖分配与海拔高度的相关性[J].植物生态学报,2012,36(1):39-46.
Wang Y F,Liu Q Q,Pei Z Y,et al.Correlation between altitude and reproductive allocation in threeSaussureaspecies on China’s Qinghai-Tibetan Plateau[J].Chinese Journal of Plant Ecology,2012,36(1):39-46.
38.廖万金,张全国,张大勇.不同海拔藜芦种群繁殖特征的初步研究[J].植物生态学报,2003,27(2):240-248.
Liao W J,Zhang Q G,Zhang D Y.A preliminary study on the reproductive features ofVeratrumnigrumL. along an altitudinal gradient[J].Chinese Journal of Plant Ecology,2003,27(2):240-248.
39.Smith C C,Fretwell S D.The optimal balance between size and number of offspring[J].The American Naturalist,1974,108(962):499-506.
40.Li W,Pan W W,Tan R,et al.Loss of preferred habitat and pollen limitation threatens reproduction in a rare mountainPaeoniadelavayi[J].Journal of Mountain Science,2016,13(12):2147-2154.
41.刘左军,杜国祯,陈家宽.不同生境下黄帚橐吾(Ligulariavirgaurea)个体大小依赖的繁殖分配[J].植物生态学报,2002,26(1):44-50.
Liu Z J,Du G Z,Chen J K.Size-dependent reproductive allocation ofLigulariavirgaureain different habitats[J].Chinese Journal of Plant Ecology,2002,26(1):44-50.
42.牛克昌,赵志刚,罗燕江,等.施肥对高寒草甸植物群落组分种繁殖分配的影响[J].植物生态学报,2006,30(5):817-826.
Niu K C,Zhao Z G,Luo Y J,et al.Fertilization effects on species reproductive allocation in an alpine meadow plant community[J].Journal of Plant Ecology,2006,30(5):817-826.
43.Niklas K J.Plant allometry:is there a grand unifying theory?[J].Biological Reviews,2004,79(4):871-889.
44.Guo H,Weiner J,Mazer S J,et al.Reproductive allometry inPedicularisspecies changes with elevation[J].Journal of Ecology,2012,100(2):452-458.
45.Weiner J,Rosenmeier L,Massoni E S,et al.Is reproductive allocation inSeneciovulgarisplastic?[J].Botany,2009,87(5):475-481.
46.Weiner J,Campbell L G,Pino J,et al.The allometry of reproduction within plant populations[J].Journal of Ecology,2009,97(6):1220-1233.
