北京植物园引进植物物候变化随纬度变化的环境解释
2018-03-26阿巴斯吐尔逊高杰刘艳红
阿巴斯·吐尔逊, 高杰, 刘艳红
北京植物园引进植物物候变化随纬度变化的环境解释
阿巴斯·吐尔逊, 高杰, 刘艳红*
北京林业大学森林培育与保护省部共建教育部重点实验室,北京 100083
探讨了北京植物园主要引进树种的物候与原产地物候差异在纬度梯度上的变化机制。通过多元回归的方法在环境梯度上建立了最佳能量模型, 降水模型和海拔模型, 以此来解释引进的24种植物物候随纬度梯度在叶期(叶芽开放, 展叶期, 盛叶期)和花期(始花期, 盛花期, 花末期)上的变化规律。结果表明: (1)物候因子变化与纬度变化呈现极显著相关(p < 0.001), 纬度梯度变化越大, 物候变化也越加剧烈。(2)海拔模型在解释物候(叶期物候, 花期物候)变化中贡献相对较小,能量模型和降水模型能够很好的解释物候(叶期物候, 花期物候)沿着纬度梯度变化的规律。(3)方差分解结果显示三个模型对花期物候和叶期物候变化的解释程度具有一定的相似性。三个模型的交互效应较高(33.7%,31.1%), 三个模型可以很好的解释物候(花期物候, 叶期物候)沿着纬度变化的规律, 可为农林业引种驯化提供科学依据。
物候变化; 引进树种; 纬度; 方差分解
1 前言
全球气候变化已引起了国内外社会的广泛关注[1]。根据政府间气候变化专门委员会的报告和预测, 1880—2012年期间全球平均温度增加了0.85℃, 到2100年全球地表温度将会上升1.1—6.4℃[2]。近百年来, 全球气候变暖背景下我国平均气温也上升了0.5—0.8℃[3]。在全球气候变化的影响下植物物候也发生了不同程度的变化[4-6], 同时植物对气候变暖也产生了相对的响应生存策略[7-9]。物候是指受非生物因子影响而出现的以年为准周期的自然季节现象[10]。物候学(Phenology)是研究由环境因子驱动的植物发育期的一门学科, 它重点研究动植物和环境因子之间的相互关系[11]。植物物候的变化研究在树种引种、选种、植树造林、森林防火, 放牧, 养蚕, 病虫害预防, 历史气候变化重建, 保护生态环境、预测气候变化趋势、环境污染监测、植物生长发育模型建立, 准确的评估植被生产力以及全球碳收支等方面都有参考作用和理论意义[12-17]。植物物候变化与气温、光照、降水等因素息息相关, 然而在不同生态系统中这些驱动因素的差异会比较大[18]。物候变化在研究植物如何应对气候变化方面已被公认为是最易于观测的, 最敏感的感应器[19]。植物物候对气候变化的响应受地理位置的影响很大[20]。地理位置的差异一定程度上反映了光照、温度、水分的差异。能量、降水以及地理生境都是物候变化的重要影响因子[21-22]。目前国内外的有关物候的研究主要集中在植物物候时空变化规律及气候变化和物候变化之间的关系[23], 然而, 物候随着时空变化的解释却很少有定量的研究。因此我们通过建立不同模型来探讨北京植物园引进树种的物候与其对应原产地的物候在纬度尺度上的差异。通过建立最佳能量模型、降水模型和生境异质性模型, 对24种引进物种的物候随着纬度梯度变化进行解释, 为今后的物候研究及林业引种提供参考方法和科学依据。
2 材料与方法
2.1 数据采集与整理
结合2010—2014年的物候数据, 我们于2015年3月初至8月底又对北京植物园100种引进树种的春季物候进行实地观测, 为了科学的比较北京植物园引进物种物候与原产地物候的变化规律, 我们从100种树种里面选出符合条件的24个树种(如表1)作为研究对象。物候观测采用《中国物候观测方法》标准, 从每个树种中选择3到5株具有代表性, 达到开花结实的, 生长发育良好且无病虫害的健壮植株作为物候观测的标准株, 并在它们向南的方向进行观测[24]。主要观测物候指标包括花期(始花期, 盛花期, 花末期)和展叶期(叶芽开放期, 展叶始期, 展叶盛期)。观测间隔为1—2 d测一次, 每天下午2点以后采用目测与望远镜结合进行观测。物候观测数据均采用Julian日换算方法, 将逐年物候现象出现日期转换成距1月1日的实际天数, 得到各物候期的时间序列。原产地物候数据则通过查阅大量有关文献获得。文献选取标准为: 已经正式发表且为几年的平均物候观测数据。
树种原产地和北京植物园的纬度、年均降水量、年均日照时数、最冷月均温(1月)、最干月降雨量通过查阅相关文献及中国气象网站获得。由于3个花期物候指标, 3个展叶期物候指标变化具有高度的相似性, 因此, 我们分别做均值作为平均花期物候指标和平均叶期物候指标。本文选取年均温、光照时数和最冷月均温作为能量模型因子; 年均降雨量、最干月降雨量选作降水模型因子、海拔(平均海拔)选作海拔模型因子。本文中, 所有因子指标都是引进物种北京与原产地气候因子之间的差值。
2.2 分析方法
首先对影响物候变化的环境变量进行正态检验, 对结果不理想的数据(平均海拔变化)取自然对数, 用Pearson相关性系数检验多个影响因子间的线性相关性; 运用单因素相关分析物候变化因子与纬度变化之间的线性回归和二次回归关系, 利用RDA分析环境因子变化矩阵对物候变化矩阵的解释程度以及变量因子之间的相互关联。依据最优线性AICc指数(Snakelike information criterion)分别筛选出能够解释24种植物花期物候与叶期物候沿着纬度梯度变化的最优的能量模型、降水模型和海拔模型。AICc指数可以在一系列备选线性模型中筛出最优模型, 即AICc值最低的模型。
然后, 采用方差分解(variation partitioning)的方法, 把平均花期物候、平均叶期物候变化能够被全部解释的因变量方差分解成能量模型、降水模型和海拔模型的独立效应以及各个模型之间的彼此交互效应; 独立作用的大小表明某一模型在解释某个物候指标变化的相对重要性。

表1 所观测树种及原产地(自然分布地)
本文所用分析与绘图软件: Sigmaplot 12.5、R 3.1.3、SAM 4.0。
3 结果分析
3.1 物候变化与纬度梯度变化关联
从图1可以看出, 纬度变化与物候变化指标间存在极显著正相关(p<0.001)。原产地与引入地(北京)的纬度相差越大, 物候变化就越剧烈。另外, 不同的花期和叶期指标随着纬度变化具有高度相似的物候变化。
3.2 物候变化与环境因子变化关联
如表2所示, 各物候指标与各潜在影响因子之间普遍存在显著关联(< 0.05), 光照时数对花期物候变化的影响程度显著高于对叶期物候变化的影响程度。年均温、最冷月均温和最干月降雨量的变化对物候变化的影响程度要高于光照时数、年均降雨量和海拔变化。
3.3 物候变化的环境模型解释
根据AICc最小值原则分别挑选各个最佳模型的解释因子, 如表3所示, 其中能量模型、降水模型、海拔模型分别能够解释花期物候变化方差的71.4%、52.5%、33.2%, 能够解释叶期物候变化的69.9%、79.2%、38.9%。海拔模型对物候变化的解释程度最低, 能量模型、降水模型对物候变化的解释程度相似, 在解释花期物候、叶期物候方差上解释程度略有差异。
方差分解结果如图2所示, 对平均花期物候而言, 能量模型A(20.1%)的独立作用要大于降水模型B(1.3%)和海拔模型C(0.8%)的独立作用。能量模型和降水模型的交互效应最大(18.2%), 三个模型的交互效应较高, 达到33.7%。三个模型总共可以解释花期物候方差的72.8%。对平均叶期物候而言, 降水模型(7.7%)的独立作用大于能量模型(4.5%)和海拔模型(0.3%)的独立作用, 能量模型和降水模型的交互效应最大(33.6%), 三个模型的交互效应较高, 达到31.1%。海拔模型与能量模型、降水模型的单独交互效应较低。在物候变化的不同时期, 能量模型和降水模型的独立作用有较大差异。
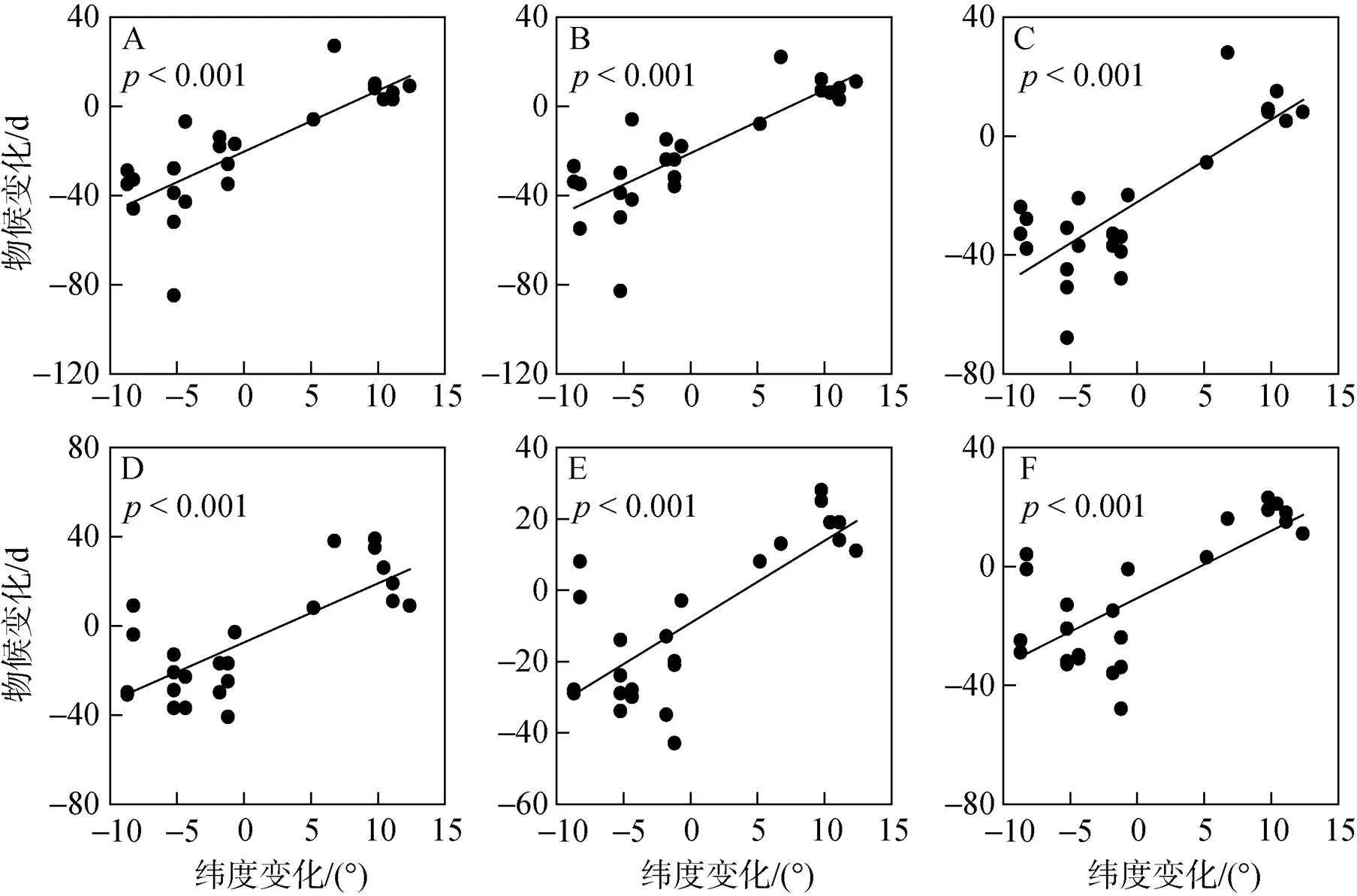
A. 始花; B. 盛花; C. 花末; D. 叶芽开放; E. 展叶始; F. 展叶盛

表2 物候变化与各解释模型下环境因子变化关联
注: *代表<0.05, **代表<0.01, ***代表<0.001, ****代表<0.0001

表3 花期、叶期物候变化的最佳解释能量模型、降水模型和海拔模型
4 讨论
龚高法等人研究发现, 我国植物春季物候的地理分布存在明显的规律性[20], 已经有不少研究表明, 全球气候变化背景下全球范围内植物的春季物候都出现提前的趋势[25–27]。在纬度梯度上, 每向北增加1个纬度, 物候推迟2.8 d, 海拔高度每升100 m推迟1.1 d[28], 维度变化对植物的花期物候有显著性的影响[29], 我国植物春季物候随着纬度向北1度会推迟3.3 d, 海拔每升100 m推迟1.6 d[30]。从图1可以看出, 树种春季物候变化和纬度变化具有显著的正相关性(p<0.001), 与郑景云等[28], Camille[29]和韩超等[30]的研究结果一致。植物物候随着纬度发生变化的主要原因是随着纬度的变化, 光照、降水、温度等环境因子都会发生相应的变化, 这些环境因子在生物完成生长发育过程中直接或间接地影响植物体内的生化反应来导致植物物候发生变化。
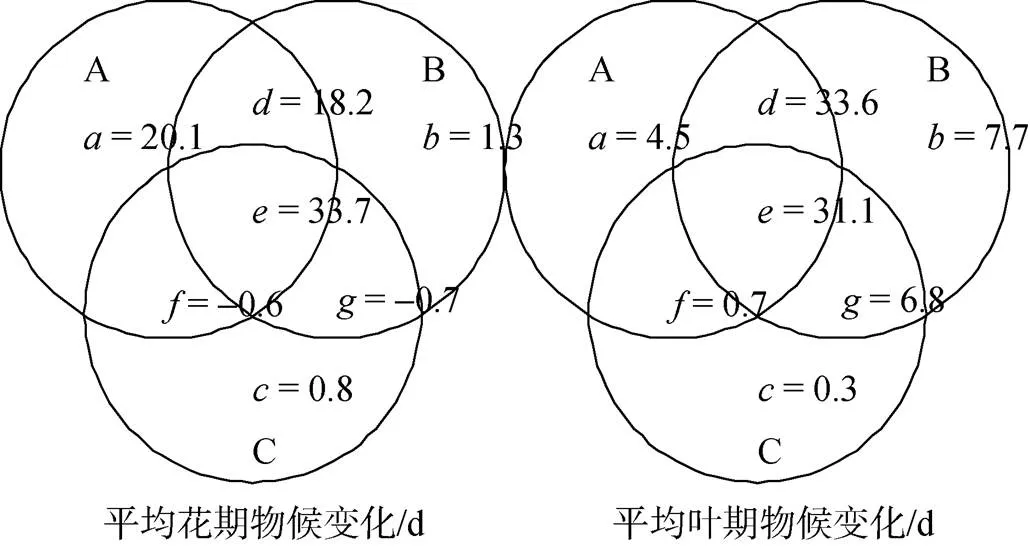
图2 能量模型(A)、降水模型(B)、海拔模型(C)所能解释平均花期物候变化、叶期物候变化方差
通过这三个模型中各因素对开花期和展叶期的独立作用和交互作用可以看出, 不同气候因子在树种不同发育阶段所起的作用有所差异。植物发育生长是一个复杂的过程, 植物完成整个生长季实际上是植物受到外界环境因素的驱动下, 通过植物体内的生化反应来实现。植物物候受多种因素的影响, 但这些因素对植物物候的影响程度是不同的[31]。温度和水分是植物萌芽物候发生的两大重要因素, 因为植物体内生化反应的发生都需要一定量的积温和水分。植物萌芽阶段水分和温度是两大重要因素, 但随着植物发芽, 展叶至能进行光合, 对外界环境进行能量交流以后, 植物对光照的要求增加, 而水分和温度只满足植物体内基本需求量的情况下绿色植物依然能生存。车少静等[32]分析石家庄春季自然物候对气候变化的响应, 发现气温, 日照和降水和物候变化的相关性程度不一样, 其中石家庄春季物候与气温的关系最好, 与降水表现出弱的相关性, 与日照关系不明显。神祥金等[33]研究长春木本植物春季物候和气候因子的关系, 发现温度对植物物候的影响最大, 而光照和降水的影响不明显。
以往的研究大多只注重某单一环境因子对植物物候变化的影响, 很少有考虑多个因子对树种物候变化的相对方差贡献。这样只考虑环境因子里面某一个因素的单独作用可能会得片面性的结论, 因为植物物候变化是外界环境因子和植物本身生理反应的综合响应结果。能量模型、降水模型、海拔模型能不同程度上解释叶期物候变化的69.9%、79.2%、38.9%, 花期物候变化方差的71.4%、52.5%、33.2%。能量模型和降水模型对树种春季物候变化的解释能力比海拔模型的解释能力强。同一树种在不同生境物候存在差异[34]。这一结果一定程度上解释影响植物物候的生态因子当中, 气候因子的影响程度比地形因子的影响更为明显。大多数研究表明, 温度是影响植物物候的主导因子, 温度的高低直接影响到植物体内活性酶的作用[35], 因而间接影响植物物候的变化。温度升高可以提前满足植物对积温的要求, 从而导致其物候的提前[36]。植物物候和气温状况相关, 特别是在植物整个生长发育阶段气温与植物各物候有显著的相关性[37]。降水变化对物候变化具有较高的解释程度。降水对植物物候变化的影响在不同地区有差异, 比如在热带和干旱地区更为明显。有研究表明, 植物开花会受到降水的影响[38]。方差分层分解的结果显示: 极端降水以及极端低温对于物候变化的解释程度较高, 物候受到极端天气的影响。然而, 对我们研究的树种来说, 这种极端天气出现在冬季, 这时间段适当的低温和降水量是植物顺利过冬进入冬芽物候至完成其他物候阶段的重要基础。有研究表明, 西双版纳经历1974年初的极端低温后, 81%的引种植物生长抽梢期提前, 35%的引种植物没有开花[39]。极端气候通过直接影响植物体内酶的活性来导致植物物候产生适应性的变化。
5 结论
植物的春季物候(花期和叶期物候)与纬度变化有显著的相关性, 随着纬度增加植物的春季物候出现推迟。不同的花期和叶期指标随着纬度变化具有高度相似的物候变化; 对展叶始期的影响比对展叶盛期影响更明显, 也即是纬度变化对植物同一个物候期不同阶段的影响不同。植物物候是植物受多种因素综合作用下产生的对外界环境变化的响应, 但这些影响因子在植物不同发育阶段的影响程度不同。极端降水和极端低温对植物春季物候影响很大。本文利用三个模型能很好的解释物候随着纬度变化的规律, 这有利于为农林业引种驯化提供良好的借鉴意义。
[1] 黄彦. 低碳经济时代下的森林碳汇问题研究[J]. 西北林学院学报, 2012, 27(3): 260–268.
[2] PARRY M L, OSVALDO F, CANZIANI C E,. Intergovernmental Panel on Climate Change Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change[M]. Cambridge, UK and New York, USA: Cambridge University Press, 2007.
[3] 周振伟, 张续, 郭大勇, 等. 黑龙江省应对气候变暖的对策[J]. 黑龙江水专学报, 2008, 35(2): 81–84.
[4] AMANO T, SMITHERS R J, SPARKS T H,. A 250-year index of first flowering dates and its responses to temperature changes[J]. Biological Science, 2010, 227: 2451–2457.
[5] 廖雪萍, 李耀先, 黄梅李, 等. 气候变化对桂林植物物候的影[J]. 广西植物, 2012, 23(4): 487–493.
[6] PARMESAN C. Influences of species , latitudes and methodologies on estimates of phenological response to global warming[J]. Global Change Biology, 2007, 13(9): 1860–1872.
[7] POST E, STRENSETH N C. Climatic variability, plant phenology and northern[J]. Ecology, 1999, 80(4): 1322–1339.
[8] PENUELAS J, FILELA I, COMAS P. Changed plant and animal life cycle from 1952 to 2000 in the mediterr anean region[J]. Global Change Biology, 2002, 8(6): 531–544.
[9] WALTHER G R, POST E, CONVEY P,. Ecological responses to recent climate change[J]. Nature, 2002, 416(6879): 389–395.
[10] 竺可桢, 宛敏渭. 物候学[M]. 北京: 科学出版社, 1973.
[11] SPARKS T H, CAREY P D. The responses of species to climate over two centuries: an analysis of the Marsh-am phenological record, 1736-1947[J]. Journal of Ecology, 1995, 83(83): 321–329.
[12] SCHWARTZ M D. Green-wave phenology[J]. Nature, 1998, 394(6696): 839–840.
[13] 张荣霞. 物候资料在农业气象工作中的应用[J]. 山东气象, 1992, (4): 41–42.
[14] 栗锡龄, 王月梅, 王有根, 等. 浅淡物候指标在农业生产上的应用[J]. 山西气象, 2006, 4: 23–25.
[15] SPANO D, CESARACCIO C, DUCE P,. Phenological stages of natural species and their use as climate indicators[J]. International Journal of Biometeorology, 1999, 42(3): 124–133.
[16] 刘亚辰, 王焕炯, 戴君虎, 等. 物候学方法在历史气候变化重建中的应用[J]. 地理研究, 2014, 33(4): 604–613.
[17] 沈志. 物候学在生态监测中的应用[J]. 干旱环境检测, 1991, 5(1): 50–52.
[18] ARORA K V, BOER G J. A parametrization of leaf phenology for the terrestrial ecosystem component of climate models[J]. Global Change Biology, 2005, 11(1): 39–59.
[19] AASA A, JAAGUS J, AHAS R,. The influence of atmospheric circulation on plant phenological phase in Central and Eastern Europe[J]. International Journal of Climatology, 2004, 24(12): 1551–1564.
[20] 龚高法, 简慰民. 我国植物物候期的地理分布[J]. 地理学报, 1983, 38(1): 33–39.
[21] 杨萍, 谌利民, 权秋梅, 等. 不同生境下柔毛淫羊藿的开花物候[J]. 西华师范大学学报: 自然学科版, 2015 , 36(1): 17–23.
[22] 韩立群, 张彦妮, 王晓丽, 等. 野生东北百合种群的生境与物候学特征[J]. 经济林研究, 2011, 29(3): 65–68.
[23] 莫非, 赵鸿, 王建永, 等. 全球变化下植物物候研究的关键问题[J]. 生态学报, 2011, 31(9): 2593–2601.
[24] 夏林喜, 牛永波, 李爱萍, 等. 浅谈木本植物物候观测要求及各物候期观测标准[J]. 山西气象, 2006 (2): 47–48.
[25] 陈效逑, 张福春. 近50年北京春季物候的变化及其对气候变化的响应[J]. 中国农业气象, 2001, 22(1): 1–5.
[26] DEFILA C, CLOT B. Phytophenological trends in Switzerland[J]. International Journal of Biometeorology, 2001, 45(4): 203–207.
[27] SCHWARTZ M D, REITER B E. Change in North American spring[J]. International Journal of Climatology, 2000, 20(8): 929–932.
[28] 郑景云, 葛全胜, 赵慧霞. 近40年中国植物物候对气候变化的响应研究[J]. 中国农业气象, 2003, 24(1): 28–32.
[29] CAMILLE PARMESAN. Influences of species, latitudes and methodologies on estimates of phenological Response to global warming[J]. Global Change Biology, 2007, (13): 1860–1872.
[30] 韩超, 丁德平, 李讯. 中国物候季节时空分布规律研究[C]// 中国气象学会2008年会. 2008: 15–21.
[31] KöRNER C, BASLER D. Phenology under global warming [J]. Science, 2010, 327(5972): 1461–1462.
[32] 车少静, 赵世林. 石家庄春季自然物候对气候变化的响应研究[J]. 山东气象, 2009, 29(1): 1–5.
[33] 神祥金, 吴正方, 刘彩, 等. 长春市木本植物春季物候对气候变化的响应[J]. 中国农学通报, 2012, 28 (1): 112– 117.
[34] 康永祥, 陈绵, 康晋, 等. 毛梾的生物学特性研究[J]. 西北林学院学报, 2012, 27(3): 57–62.
[35] 李荣平, 周广胜, 张慧玲. 植物物候研究进展[J]. 应用生态学报, 2006, 17(3): 541–544.
[36] 肖登攀, 陶福禄. 过去30年气候变化对华北平原冬小麦物候的影响研究[J]. 中国生态农业学报, 2012, 20 (11): 1539–1545.
[37] 王连喜, 陈怀亮, 李琪, 等. 植物物候与气候研究进展[J]. 生态学报, 2010, 30(2): 0447–0454.
[38] AUSPURGER C K. Phenology, flowering synchrony and fruit set of 6 Neotropical Shrubs[J]. Bitropica, 1983: 15(4): 257–267.
[39] 赵俊斌, 张一平, 宋富强, 等. 引种保护植物对西双版纳极端冷年的春季物候响应[J]. 植物学报, 2010, 45(4): 435–443.
Environmental interpretation of the phenological change of introduced trees with latitudinal change in the Beijing Botanical Garden
Abasi Tuerxun, Gao Jie, Liu Yan-hong*
Key Laboratory for Silviculture and Conservation, Ministry of Education/Beijing Forestry University, Beijing 100083, China
The phenological variation mechanism of the main introduced tree species in Beijing Botanical Garden and their natural distribution areas along with latitudinal gradient were analyzed. By using multiple regression method to establish an optimum energy model and elevation model which are based on the environmental gradient, it explained the variation mechanism of flowering phenology (initial flowering stage, full booming stage, end of flowering) and leaf phenology (bud bursting, beginning of leaf expansion, leaf full expansion) indexes of 24 introduced tree species along with latitudinal gradient. Resultsare as follows. (1) The change of phenological index has a significant correlation with latitudinal change (<0.001), and thus the phenological change growth is intense with the increasement in latitude gradient.(2) The contribution of the elevation model to explain phenological (leafing, flowering) variance is relatively small compared with energy model and precipitation model which explain well the regulation of phenological (leafing, flowering) change with respect to latitudinal gradient.(3) All the three models have certain similarity while explaining the leaf and flowering phenology. The interaction effect of the three models reaches up to 33.7% and 31.1%, respectively, and can explain well the phenological change along with latitudinal gradient, which can provide a good basis for agro-forestry introduction and domestication.
Phenological change; introduced tree species; latitude; variance partitioning
S718.5
A
1008-8873(2018)01-071-07
10.14108/j.cnki.1008-8873.2018.01.010
2016-12-19;
2017-12-29
北京市重点学科项目“生态学”(20140801)
阿巴斯·吐尔逊(1987一), 男, 新疆拜城县人, 硕士, 主要从事生物多样性保护研究, E-mail: 2938196199@qq.com
刘艳红, 女, 博士, 教授, 主要从事生物多样性保护和森林生态学研究, E-mail: liuyh@bjfu.edu.cn
阿巴斯·吐尔逊, 高杰, 刘艳红. 北京植物园引进植物物候变化随纬度变化的环境解释[J]. 生态科学, 2018, 37(1): 71-77.
Abasi Tuerxun, Gao Jie, Liu Yan-hong. Environmental interpretation of the phenological change of introduced trees with latitudinal change in the Beijing Botanical Garden[J]. Ecological Science, 2018, 37(1): 71-77.
