大豆生物解离过程中形成的乳状液结构特征
2018-03-20江连洲王立敏隋晓楠扈莹莹
江连洲,王立敏,隋晓楠,毕 爽,丁 俭,张 亮,高 宇,扈莹莹,李 杨*
(东北农业大学食品学院,黑龙江 哈尔滨 150030)
生物解离乳状液是生物解离提油过程中由于亲水、亲油的两性物质(磷脂、蛋白质)存在而形成的一种复杂稳定体系[1],也是一种易受外界条件变化而影响其稳定性的典型弹性体系[2]。生物解离是近几年来新兴的一种提油方法,该技术经济、安全、绿色环保,且由于该技术处理条件温和,提取的油脂品质较高、色泽较好[3]。但是,生物解离提油中乳状液保留了部分油脂,限制了其油脂提取率,很大程度限制了该技术的工业化应用。因此,探究限制乳状液形成或者破除的方法,从而达到油脂最大提取率是普及该技术的重要途径之一。但乳状液的形成与其组成、界面性质、各组分分子的空间构象及组分间的分子交互作用等因素息息相关。仅研究乳状液的破除技术并不足以从本质上解决现存问题。而现今文献研究大多数集中对乳状液的破除技术、组分分析及界面性质,Chabrand等[4]研究了酶法破乳,获得了95%的提油率且对乳状液中界面蛋白进行分析,得出界面蛋白浓度为(11.40±0.35)mg/m2。Lamsal等[5]对乳状液进行多方面研究,其中包括乳状液的组分分析,发现乳状液中含有约57%油脂、35%水分、5%蛋白质、1.3%多糖和0.8%磷脂。de Moura等[6]分析了乳状液中的氨基酸组成,发现界面蛋白的疏水性氨基酸残基与乳状液的稳定性有密切关系;Chabrand等[7]发现在乳状液的油-水界面上存在液晶相,使得乳状液相当稳定;王瑛瑶等[8]研究了乳状液中肽的分子质量分布,发现界面吸附分子质量大部分位于3~10 kDa,具有较强的乳化稳定性。相关研究尚未涉及乳状液结构特征领域,但透彻解析乳状液结构特征是揭示乳状液稳定机制的重要途径。
本研究对大豆不同酶解时间(1、2、3 h)生物解离乳状液的结构特征进行解析,从流体动力学半径及其分布、油脂与蛋白空间分布进行分析和表征;通过乳状液与对应酶解时间下大豆分离蛋白(soy protein isolate,SPI)之间的对比,研究乳状液中二硫键、发色氨基酸及油脂-蛋白之间的相互作用,明确不同酶解时间乳状液荧光强度变化机制。通过对乳状液结构特征解析,以期为探究大豆生物解离乳状液的稳定性机制、寻求适当的破乳方法提供理论依据与指导。
1 材料与方法
1.1 材料与试剂
大豆分离蛋白 东北农业大学大豆实验室自制;挤压膨化大豆粉(蛋白40%、脂肪19.6%、纤维25.6%,以大豆粉质量为基数计) 山东高唐蓝山股份有限公司;Protex6L碱性蛋白酶(8 900 U/mL) 诺维信生物技术有限公司;正己烷 美国Sigma公司;盐酸、氢氧化钠等试剂均为分析纯。
1.2 仪器与设备
磁力搅拌器 广州仪科实验仪器有限公司;水浴锅英国IKA公司;BRAVO手持式拉曼光谱仪 德国布鲁克仪器公司;AIR MP激光扫描共聚焦显微镜 日本岛津公司;Allegra64R台式高速冷冻离心机 美国贝克曼公司。
1.3 方法
1.3.1 生物解离乳状液的制备
参考李杨等[9]的方法,称取一定量的挤压膨化大豆粉,加入2%(以大豆粉质量为基数计)Protex6L酶制剂,按照料液比1∶6(m/V)加入水,用玻璃棒搅拌均匀,在55 ℃水浴锅中保持恒温,用1 mol/L NaOH溶液调节水溶液的pH值保持在9.0,边搅拌边酶解,酶解1、2、3 h后,用2 mol/L HCl溶液调节水溶液的pH值至7.0,之后在100 ℃沸水中灭酶5 min。酶解结束后进行离心操作(4 500 r/min、20 min),离心后除去残渣,将游离油、乳状液和水解液倒入分液漏斗中,分离得到乳状液。
1.3.2 SPI及酶解物的制备
参考Wolf等[10]的方法。大豆磨粉后与正己烷以1∶6(m/V)的比例混合,在常温下搅拌脱脂3 次,每次2 h。将脱脂豆粉按1∶10(m/V)的比例与水混合,然后用2 mol/L NaOH溶液调节pH值至8.5,45 ℃搅拌2 h后,将其悬浮液在4 ℃、10 000×g条件下离心20 min,取上清液再用2 mol/L HCl溶液调节pH值至4.5。静置后在4 ℃、6 000×g条件下离心20 min,所得蛋白沉淀水洗2 次,最后用2 mol/L NaOH溶液调节蛋白质pH值至7.0。将此蛋白溶液冷冻干燥后研磨即得粉末状SPI。将得到的SPI在与制备乳状液相同的条件下酶解1、2、3 h得到酶解物。
1.3.3 拉曼光谱分析
实验所用的拉曼光谱仪为BRAVO手持式拉曼光谱仪。扫描波长785 nm,功率为300 mW,扫描范围400~2 000 cm-1,扫描时间每次60 s,积分10 次,4 次扫描进行累加。以苯丙氨酸((1 003±1)cm-1)作为归一化因子,得到乳状液、SPI及其酶解物的拉曼光谱图。谱图基线校正、谱峰归属查找采用ACD Labs V12软件进行。
1.3.4 激光扫描共聚焦显微镜
参考李国平等[11]的方法将新鲜制备的乳状液样品滴在显微镜的载玻片上,启动LSC软件,选择光路参数(Bean):扫描模式为xyz;扫描形式为512×512或1 024×1 024;扫描速率为400 Hz;Bean Exp为6;Pinhole为1;扫描方向为Uni。伪彩色均采用绿色,调整P-T、Zoom及Position大小,直至获取高质量的图像。通过单次扫描和连续扫描方式进行图像采集。由于乳状液中蛋白荧光强度较强,因此无需染色,在对乳状液分层扫描后得到的“光学切片”经过三维重建后,得到不同酶解时间下乳状液的空间分布。
1.3.5 流体动力学半径及其分布
参考Tang等[12]的测定方法。采用动态光散射技术,利用Mastersize 2000激光粒度分析仪测定乳状液的粒度分布,对新鲜的乳状液样品进行测定。参数设置为:颗粒折射率1.520,颗粒吸收率0.001,以水为分散剂,折射率为1.330。实验采用D4,3,即体积平均直径表征液滴粒度的大小,乳状液样品制备出后立即测定,每个样品重复3 次。
1.4 数据分析
每个实验重复3 次,采用Origin8.0软件作图。采用SPSS V17.0软件进行方差分析(analysis of variance,ANOVA)及相关性分析,P<0.05为显著性差异。
2 结果与分析
2.1 不同酶解时间对乳液流体动力学半径及分布的影响
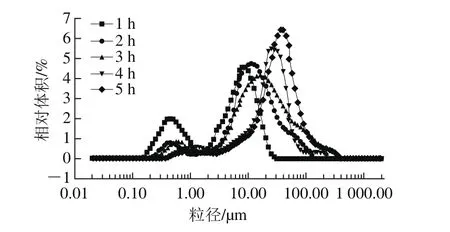
图1 不同酶解时间下乳状液的粒径分布Fig.1 Particle size distribution of emulsions at different hydrolysis times
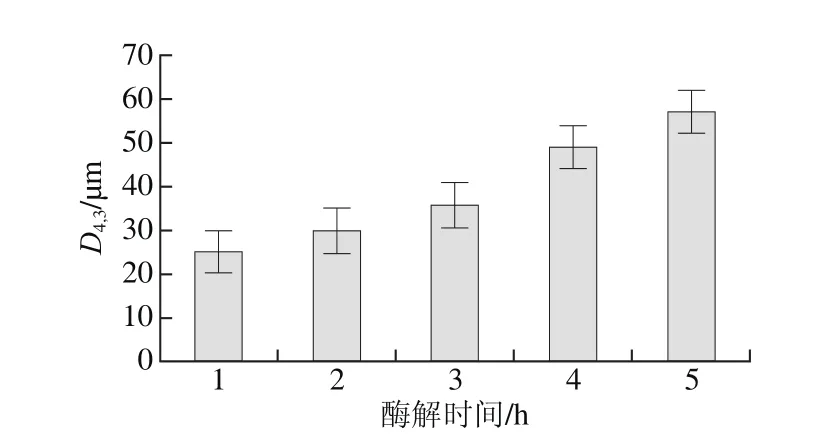
图2 不同酶解时间下乳状液体积平均粒径D4,3Fig.2 Average particle diameters (D4,3) of emulsions at different hydrolysis times
流体动力学半径分布是评价乳液稳定性的重要指标[13],同时可以直观地展示乳液中油滴的分散状态。本研究利用动态光散射技术探究不同酶解时间下乳液中油滴的分布情况及体积平均粒径(D4,3)的大小。由图1可以看出,本实验得出不同酶解时间下乳液粒径分布相似,且与Wu等[1]的研究结果相似。由图2可以看出,酶解时间越长,乳状液D4,3越大,说明酶解过程中乳状液油滴不断聚集,这是由于Protex蛋白酶改变了油滴表面的蛋白,油滴表面的静电作用发生变化,同时酶解时间延长增加了分子碰撞的机率,因此油滴分子趋于聚集[14]。同时发现,酶解4 h和5 h时D4,3明显增大,10~100 μm粒径区间相对体积增大,而0.1~1 μm粒径区间相对体积减小,这说明这时乳状液油滴开始大量聚集,较不稳定;Jung等[15]研究指出酶解3 h时酶解程度已经达到最大、提油率最高,无需再继续酶解,因此后续研究以酶解1、2、3 h的乳状液为研究对象。
2.2 不同酶解时间对乳状液三维空间结构的影响
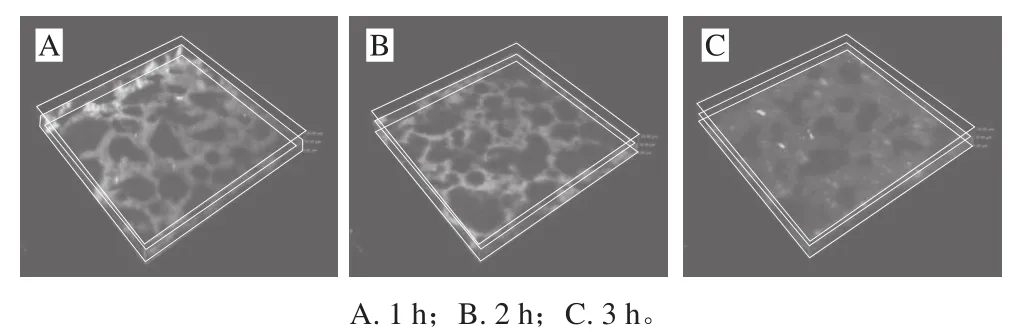
图3 不同酶解时间下乳状液中蛋白质的空间分布Fig.3 Spatial distribution of proteins in emulsions at different hydrolysis times
激光扫描共聚焦显微镜,可以实现对样品进行断层扫描、成像、无损伤观察,更清晰、直观地观察乳状液中油脂与蛋白的空间分布[16]。图3为不同酶解时间下乳状液中蛋白质的空间分布情况。根据乳状液中油脂不发荧光,蛋白具有较强荧光的特点,可知绿色荧光即乳状液中蛋白分布,黑色为油脂分布。通过观察可知,酶解1 h时,乳状液中蛋白相和油脂相分离,油滴分布密集,油滴小而多,油滴外面包裹一层蛋白膜起到稳定作用,这与之前研究相符合[17];酶解3 h得到的乳状液蛋白与油脂相混乱,油滴分散、变大,呈现许多不规则的形状。这是由于在酶解过程中,油滴界面吸附蛋白被酶解成小分子肽,从油滴表明脱落进入游离相中,致使相邻的小油滴发生聚集合并,呈现出不规则的状态[18]。
此外,还可以看出荧光强度逐渐降低。荧光强度是表征蛋白质结构的重要指标,通过荧光强度的变化,可以得知蛋白质的几种重要的氨基酸残基所处的微环境变化,它与蛋白质疏水性基团有关[19]。王瑞等[20]研究发现,大豆蛋白中产生荧光的氨基酸残基多分布于蛋白质分子内部,酶解处理可以使蛋白球形结构变得松散伸展,深埋在球状结构内部的疏水性基团暴露增多,荧光强度增加。然而在本研究中,由图3观察可知,酶解时间越长,乳状液荧光强度下降明显。主要是由于酶解降低了乳液中蛋白质的质量浓度及分子质量,导致乳状液荧光强度下降;同时,Yang等[21]研究也指出,蛋白质的荧光猝灭与分子内芳香族氨基酸残基、二硫键构象及蛋白分子疏水相互作用密切相关。
2.3 不同酶解时间对乳状液蛋白质结构的影响
拉曼光谱是研究分子相互作用的一种工具,因此常采用拉曼光谱法分析乳状液蛋白氨基酸及二硫键构象的变化。乳状液及SPI拉曼光谱图中对应的谱峰及谱峰指认见表1。

表1 乳状液及SPI拉曼光谱的特征峰指认Table1 Raman band assignment of SPI and emulsion
在拉曼光谱测定中,谱线强度与散射中心(化学键和基团)数目为正比例关系[22]。因此,可以根据样品的拉曼谱线强度来判断乳状液中某些化学键或基团的改变程度,谱线强度变小意味着乳状液中蛋白或油脂对应的基团或化学键受到损伤,谱线的偏移说明它所对应的化学键或基团在不同的环境中发生了变化[19]。
2.3.1 二硫键分析
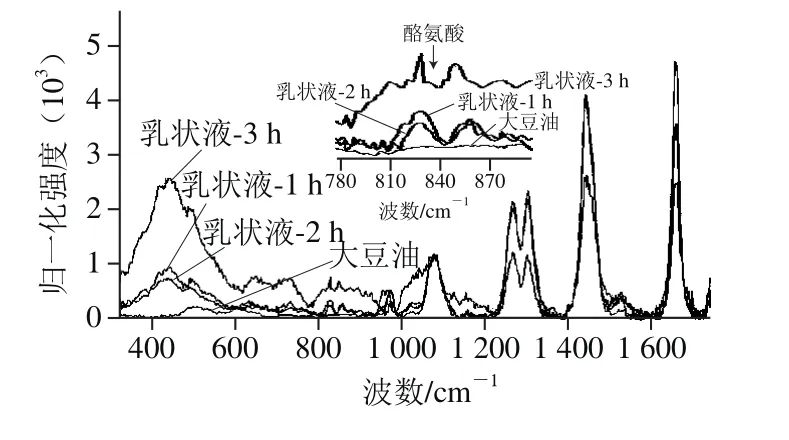
图4 酶解时间对乳状液结构影响的拉曼光谱分析图Fig.4 Raman spectroscopy analysis of emulsions at different hydrolysis times
由图4可知,随着酶解时间延长乳状液中二硫键含量增多,酶解3 h得到的乳状液中二硫键对应的峰值最高。Howell等[23]发现乳液体系中油脂的存在可以使蛋白的构象发生改变,推测酶解过程中由于油脂蛋白之间的相互作用导致二硫键含量增多,二硫键连接的聚集体增加,乳状液中荧光减弱。
在拉曼光谱中,二硫键的特征谱带为500~550 cm-1。在该区间中拉曼位移与振动模式的对应关系为:500~510 cm-1处为gauche-gauche-gauche(g-g-g)模式,515~525 cm-1处为gauche-gauche-trans(g-g-t)模式,535~545 cm-1处为trans-gauche-trans(t-g-t)模式。由图5可知,SPI为t-g-t构型,即分子间二硫键构型;乳状液为g-g-g构型,即分子内二硫键构型[22],说明由于油脂-蛋白疏水相互作用改变了乳状液中二硫键构象。Yang等[21]研究蛋白质的荧光猝灭与二硫键构象及二硫键连接的聚集体有关。
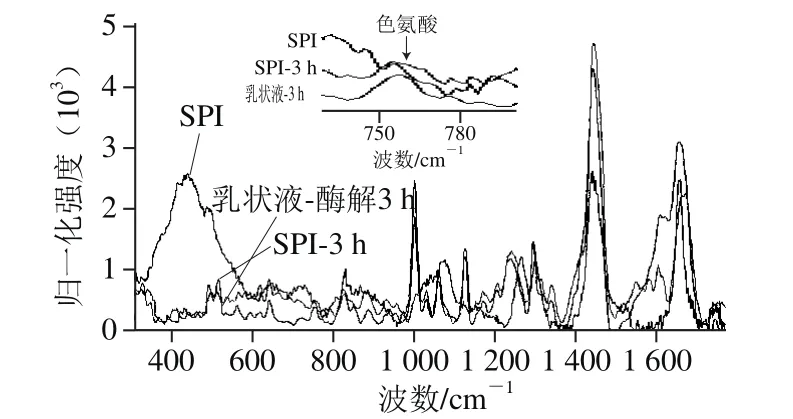
图5 酶解时间对SPI及乳状液结构影响的拉曼光谱分析图Fig.5 Raman spectroscopy analysis of emulsion and SPI hydrolysate
2.3.2 发色基团的分析
表2 酪氨酸费米共振线I850cm-1/I830 cm-1以及色氨酸I760cm-1/I1 003 cm-1Table2 Fermi resonance doublet ratio I850cm-1/I830 cm-1 for tyrosyl andfor tryptophan

表2 酪氨酸费米共振线I850cm-1/I830 cm-1以及色氨酸I760cm-1/I1 003 cm-1Table2 Fermi resonance doublet ratio I850cm-1/I830 cm-1 for tyrosyl andfor tryptophan
注:同列肩标字母不同表示不同酶解时间的I850cm-1/I830cm-1及酪氨酸残基差异显著(P<0.05)。**.标准强度为拉曼光谱各条带强度与苯丙氨酸(1 003 cm-1)相对强度的比值。
?
酪氨酸的费米共振会引起850 cm-1和830 cm-1附近的特征峰随侧链微环境改变,用这2 条谱线的强度比,可以推测蛋白质分子中的酪氨酸残基的暴露与埋藏。I850cm-1/I830cm-1的比值为1.25~1.40时,表明酪氨酸残基完全暴露于分子表面(“暴露态”);比值为0.3~0.5时,酪氨酸残基完全埋藏于分子内部(“包埋态”)[19]。由表2可知,酶解3 h后SPI相对于未酶解SPI的I850cm-1/I830cm-1比值增大,I850cm-1/I830cm-1=1.21时,即酪氨酸完全暴露于分子表面。乳状液中I850cm-1/I830cm-1随着酶解时间延长越来越小,酶解3 h时,乳状液I850cm-1/I830cm-1=0.46,酪氨酸由“暴露”变化为“包埋”态[24]。Herrero等[25]研究发现当蛋白质遇到油脂时,其疏水基团会倾向于油相内,造成酰基与蛋白质侧链之间的疏水基团接触,引起蛋白质氨基酸无序排列,使蛋白分子发生交联。因此可以推断乳状液中由于油脂与蛋白的相互作用使蛋白部分发生交联聚集,导致发色基团被包埋在分子内部;另外,乳状液中蛋白与油脂疏水相互作用也会屏蔽分子内部的疏水性基团,产生空间位阻,使发色团被掩盖,导致蛋白荧光减弱[26]。或许乳状液中小分子的作用也使蛋白结构发生了改变,这一假设需要以后进一步研究。
756 cm-1附近的拉曼谱带归属为色氨酸侧链,发色氨基酸中,色氨酸发色能力最强,已有研究表明,756 cm-1附近的色氨酸拉曼归属谱带强度越低,蛋白质的色氨酸趋于“暴露”态,反之则趋于“包埋”态[27]。由图5可知,酶解后的SPI色氨酸峰值与原SPI相比显著降低,这与上述酪氨酸变化机制相似。但是在乳状液中,色氨酸对应的峰值不太明显,这可能是因为乳状液中油脂与蛋白质的相互作用或蛋白质周围小分子的影响改变了蛋白质的分子结构并导致疏水基团暴露[23]。
3 结 论
本研究首先利用动态光散射技术、激光扫描共聚焦显微镜得到了不同酶解时间(1、2、3 h)下乳状液的粒径分布及蛋白质与油脂的空间分布,又通过SPI与乳状液对比,从分子间相互作用出发,利用拉曼光谱分析了乳状液中荧光强度变弱机理,主要结论如下:
共聚焦显微镜及粒径分布结果显示:酶解时间延长,乳状液粒径变大,油滴数量减少,并呈现出不规则的形状,由于在酶解过程中,乳状液中蛋白质被酶解成小分子肽,从油滴表面脱落,进入游离相中,致使相邻的小油滴发生聚集合并。同时,在乳状液空间结构中荧光强度呈现明显的减弱趋势。
拉曼光谱分析乳状液中荧光强度减弱机理结果显示:与酶解后的SPI相比,乳状液中二硫键发生了偏移,且随着酶解时间延长,乳状液中二硫键含量增多,表明乳状液中油脂与蛋白质之间的相互作用改变了二硫键构象,同时导致二硫键连接的聚集体含量增加;酶解SPI时,I850cm-1/I830cm-1>1.2,酪氨酸完全暴露,但在乳状液中I850cm-1/I830cm-1<0.5,酪氨酸由“暴露”变化为“包埋”态,且随着酶解时间延长,I850cm-1/I830cm-1越来越小,推测乳状液中由于油脂与蛋白质的相互作用使蛋白质部分交联聚集,导致发色基团被包埋在分子内部;另外,乳状液中蛋白质与油脂疏水相互作用屏蔽分子内部的疏水性基团,产生空间位阻,使发色团被掩盖,导致蛋白荧光减弱;酶解SPI时,色氨酸被暴露,但在乳状液中没有出现对应的峰,推测乳状液中油脂与蛋白质的相互作用或蛋白周围小分子的影响改变蛋白质的分子结构并导致疏水基团暴露的变化。或许乳液中小分子的作用也使乳液中蛋白结构发生改变,这一假设需要以后进一步研究。
上述结论对于系统地探讨乳状液复杂体系内分子间作用力、分子组成和空间构象提供了一定理论支持,同时,也为开发新型高效破乳技术提供理论参考。
[1] WU J, JOHNSON L A, JUNG S. Demulsification of oil-rich emulsion from enzyme-assisted aqueous extraction of extruded soybean flakes[J]. Bioresource Technology, 2009, 100(2): 527-533.DOI:10.1016/j.biortech.2008.05.057.
[2] 王瑛瑶, 王璋, 罗磊. 水酶法提花生油中乳状液性质及破乳方法[J].农业工程学报, 2008, 24(12): 259-263.
[3] JMLN D M, CAMPBELL K, DE ALMEIDA N M, et al. Protein extraction and membrane recovery in enzyme-assisted aqueous extraction processing of soybeans[J]. Journal of the American Oil Chemists’ Society, 2011, 88(6): 877-889. DOI:10.1007/s11746-010-1737-0.
[4] CHABRAND R M, GLATZ C E. Destabilization of the emulsion formed during the enzyme-assisted aqueous extraction of oil from soybean fl our[J]. Enzyme and Microbial Technology, 2009, 45(1): 28-35. DOI:10.1016/j.enzmictec.2009.03.008.
[5] LAMSAL B P, JOHNSON L A. Separating oil from aqueous extraction fractions of soybean[J]. Journal of the American Oil Chemists’ Society, 2007, 84(8): 785-792. DOI:10.1007/s11746-007-1090-0.
[6] DE MOURA J M L N, CAMPBELL K, MAHFUZ A, et al. Enzymeassisted aqueous extraction of oil and protein from soybeans and cream de-emulsif i cation[J]. Journal of the American Oil Chemists’ Society,2008, 85(10): 985-995. DOI:10.1007/s11746-008-1282-2.
[7] CHABRAND R M, HYUNJUNG K, CHENG Z, et al. Destabilization of the emulsion formed during aqueous extraction of soybean oil[J].Journal of the American Oil Chemists’ Society, 2008, 85(4): 383-390.DOI:10.1007/s11746-008-1199-9.
[8] 王瑛瑶, 王璋. 水酶法提油中乳状液的特性研究[J]. 食品科学, 2009,30(11): 112-115.
[9] 李杨, 江连洲, 魏东旭, 等. 水酶法制取大豆油和蛋白关键技术及机理研究[C]//中国食品科学技术学会第七届年会论文摘要集. 北京:中国食品科学技术学会, 2010: 2.
[10] WOLF W J. Soybean proteins. their functional, chemical, and physical properties[J]. Journal of Agricultural and Food Chemistry, 1970,18(6): 969-976. DOI:10.1021/jf60172a025.
[11] 李国平, 黄群策, 秦广雍. 用激光扫描共聚焦显微镜观察雪松花粉和花粉管[J]. 激光生物学报, 2006, 15(1): 1-8.
[12] TANG C H, CHOI S M, MA C Y. Study of thermal properties and heat-induced denaturation and aggregation of soy proteins by modulated differential scanning calorimetry[J]. International Journal of Biological Macromolecules, 2007, 40(2): 96-104. DOI:10.1016/j.ijbiomac.2006.06.013.
[13] LIU F, TANG C H. Emulsifying properties of soy protein nanoparticles:inf l uence of the protein concentration and/or emulsif i cation process[J].Journal of Agricultural and Food Chemistry, 2014, 62(12): 2644-2654.
[14] MCCLEMENTS D J. Critical review of techniques and methodologies for characterization of emulsion stability[J]. Critical Reviews in Food Science and Nutrition, 2007, 47(7): 611-649.DOI:10.1080/10408390701289292.
[15] JUNG S, MAURER D, JOHNSON L A. Factors affecting emulsion stability and quality of oil recovered from enzyme-assisted aqueous extraction of soybeans[J]. Bioresource Technology, 2009, 100(21):5340-5347. DOI:10.1016/j.biortech.2009.03.087.
[16] TURILLAZZI E, KARICH S B, NERI M, et al. Confocal laser scanning microscopy. using new technology to answer old questions in forensic investigations[J]. International Journal of Legal Medicine,2008, 122(2): 173-177. DOI:10.1007/s00414-007-0208-0.
[17] MOSCHAKIS T, MURRAY B S, DICKINSON E. Particle tracking using confocal microscopy to probe the microrheology in a phaseseparating emulsion containing nonadsorbing polysaccharide[J].Langmuir the Acs Journal of Surfaces and Colloids, 2006, 22(10):4710-4719. DOI:10.1021/la0533258.
[18] 赵翔. 花生水剂法提油过程形成乳状液的酶法破乳研究[D]. 郑州:河南工业大学, 2012.
[19] YAN X N, LIU B S, CHONG B H, et al. Interaction of cefpiramide sodium with bovine hemoglobin and effect of the coexistent metalion on the protein-drug association[J]. Journal of Luminescence, 2013,142: 155-162. DOI:10.1016/j.jlumin.2013.04.009.
[20] 王瑞, 李杨, 王中江, 等. 体外模拟消化过程中大豆蛋白的荧光光谱分析及热处理的影响[J]. 食品工业科技, 2016, 37(6): 128-132.DOI:10.13386/j.issn1002-0306.2016.06.017.
[21] YANG E S, YANG J H, PARK J W. Inactivation of NADP+-dependent isocitrate dehydrogenase by lipid peroxidation products[J]. Free Radical Research, 2004, 38(3): 241-249.DOI:10.1080/10715760400022996.
[22] 谢凤英, 马岩, 王晓君, 等. 拉曼光谱分析荞麦多酚对米糠蛋白结构的影响[J]. 食品科学, 2017, 38(3): 32-36. DOI:10.7506/spkx1002-6630-201703006.
[23] HOWELL N K, HERMAN H, LI-CHAN E C. Elucidation of proteinlipid interactions in a lysozyme-corn oil system by Fourier transform Raman spectroscopy[J]. Journal of Agricultural and Food Chemistry,2001, 49(3): 1529-1533.
[24] WONG H W, CHOI S M, PHILLIPS D L, et al. Raman spectroscopic study of deamidated food proteins[J]. Food Chemistry, 2009, 113(2):363-370. DOI:10.1016/j.foodchem.2008.09.027.
[25] HERRERO A M, CARMONA P, PINTADO T, et al. Olive oil-inwater emulsions stabilized with caseinate: elucidation of protein-lipid interactions by infrared spectroscopy[J]. Food Hydrocolloids, 2011,25(1): 12-18. DOI:10.1016/j.foodhyd.2010.04.014.
[26] DEVLIN M T, LEVIN I W. Acyl chain packing properties of deuterated lipid bilayer dispersions: vibrational Raman spectral parameters[J]. Journal of Raman Spectroscopy, 1990, 21(7): 441-451.
[27] LI-CHAN E C Y. The applications of Raman spectroscopy in food science[J].Trends in Food Science and Technology, 1996, 7(11): 361-370.
