不同浓度CaCl2对宜昌润楠幼苗的抗寒性研究
2018-01-30况红玲喻峰杜丹张欣王昆贺心茹费永俊
况红玲+喻峰+杜丹+张欣+王昆+贺心茹+费永俊

摘要:本文以多年生宜昌润楠(Machilusichangensis)为材料,采取室内控温系统模拟低温胁迫条件,研究不同浓度CaCl2处理对低温胁迫下宜昌润楠幼苗叶绿素含量、丙二醛含量、可溶性蛋白含量、可溶性糖含量、超氧化物歧化酶活性、过氧化物酶活性以及激素ABA含量的影响,初步探究CaCl2对润楠幼苗抗寒性的调节机理。结果表明:(1)宜昌润楠幼苗在-5℃条件下喷施5mmol/LCaCl2后叶绿素含量最高。(2)宜昌润楠幼苗在5℃条件下喷施25mmol/LCaCl2后丙二醛(MDA)含量最低。(3)宜昌润楠幼苗在低温胁迫条件下,通过喷施CaCl2能影响可溶性蛋白的含量。(4)宜昌润楠幼苗在-5℃条件下喷施25mmol/LCaCl2后可溶性糖含量最高。(5)宜昌润楠幼苗在低温胁迫条件下,通过喷施CaCl2能提高SOD和POD的活性,其中在0℃条件下,宜昌润楠幼苗经5mmol/LCaCl2喷施后的SOD活性达到最高,在-5℃条件下宜昌润楠幼苗经5mmol/LCaCl2喷施后的POD活性达到最高。(6)宜昌润楠幼苗在低温胁迫条件下,通过喷施CaCl2能提高激素ABA含量,在-5℃条件下喷施5mmol/LCaCl2后ABA含量最高。
关键词:宜昌润楠;幼苗;CaCl2;抗寒性
中图分类号:S79227文献标识码:A文章编号:1004-3020(2017)06-0014-06DifferentConcentrationsofCaCl2onColdResistanceofYichangRunanSeedlingKuangHongling(1)YuFeng(1)DuDan(1)ZhangXin(1)WangKun(1)HeXinru(2)FeiYongjun(2)
(1.FayeEcologicalEnvironmentGroupco.,LTD.Wuhan430206;2.YangtzeUniversityJingzhou434100)
Abstract:Inthispaper,theeffectsofdifferentconcentrationsofCaCl2onmalondialdehyde(MDA)content,solubleproteincontentandsolublesugarcontentofYunnanseedlinginYichangwerestudiedbyusingthetemperaturecontrolsystemundertheconditionoflowtemperaturestress.Theactivityofchlorophyll,thecontentofchlorophyllandtheABAcontentofthehormonewerestudied.ThemechanismofCaCl2onthecoldresistanceofRunonseedlingwasexplored.Theresultsshowedthat:(1)Thecontentofchlorophyllwasthehighestwhenspraying5mmol/LCaCl2at-5℃.(2)Thecontentofmalondialdehyde(MDA)wasthehighestwhenspraying25mmol/LCaCl2at5℃undertheconditionof5℃.(3)ThecontentofsolubleproteinwasincreasedbysprayingCaCl2undertheconditionoflowtemperaturestress.(4)Thecontentofsolublesugarwasthehighestwhenspraying25mmol/LCaCl2at-5℃.(5)TheactivityofSODandPODcouldbeincreasedbysprayingCaCl2undertheconditionoflowtemperaturestress,andtheactivityofSODwasthehighestaftersprayingwith5mmol/LCaCl2at0℃.TheactivityofPODwasthehighestafterspraying5mmol/LCaCl2at-5℃.(6)ThecontentofABAwasincreasedbysprayingCaCl2undertheconditionoflowtemperaturestress,andthecontentofABAwasthehighestat5℃for5mmol/LCaCl2.
Keywords:YichangRunan;seedling;CaCl2;coldresistance
潤楠属的植物在全球大约有100个种,主要散布在东部的热带、亚洲东南部和亚热带区域。我国大约有68种3个变种,分布于西南部和南部地区[1]。润楠属植物包括宜昌润楠(M.ichangensis)、黄枝润楠(M.versicolora)、滇润楠(M.yun-nanensis)、梨润楠(M.pomifera)、华润楠(M.chinensis)、绒毛润楠(M.velutina)、刨花润楠(M.pauhoi)、黄绒润楠(M.grijsii)和粉叶润楠(M.glaucifolia)等。
宜昌润楠为樟科(Lauraceae)润楠属(Machilus),是常绿阔叶树种,分布于陕西南部、湖北、四川等地,是我国的特有树种,属于国家二级保护植物。其生长海拔为560~1400m山坡,高7~15m,树冠卵形。花期4月,果期8月。果黑色,近球形,直径约为10mm。小枝细短、无毛。顶芽近球形,芽磷呈圆形,边缘有浓密的缘毛。叶柄8~20(25)mm;叶常集生在当年生枝上,呈长圆状披针形至长圆状倒披针形,长10~24cm,宽2~6cm。叶正面无毛,中脉背面突起,正面凹面,侧脉纤细[1]。宜昌润楠树姿优美,四季常青,材质俱佳,兼有驱虫、杀菌、减噪、净化空气等生态功能,同时,可用作城市行道树,是具有潜在开发性的园林树种[2]。在温度比较低的地区是否能种植此树种,要取决于其幼苗是否可以忍耐低温的危害[3]。endprint
低温胁迫,是植物生长和发育过程中的一种典型环境胁迫因子。植物从种子萌发、开发、结果以及休眠等各个生长阶段均受到环境温度的制约,低温胁迫下会干扰植物的正常生理,产生冻害和冷害两种现象[4]。植物体具有抗寒特性,而不同的植物种类、品系中,甚至同种植物不同组织中抗寒能力存在着差异性[5]。因此,探明植物的抗寒机理是开展植物育种、合理引种、防寒栽培的基础工作,是植物学研究的重要内容之一,对于减少自然灾害具有重要的经济价值。本研究从生理水平对宜昌润楠幼苗抗寒性进行研究,为润楠属的抗寒性及地方引种栽培提供依据。
1材料与方法
1.1试验材料
试验取材于荆州市长江大学西校区盆景园的一年生宜昌润楠的实生苗。荆州市位于东经111°15′~114°05′,北纬29°26′~31°37′,地处湖北省中南部,江汉平原腹地,属亚热带过渡性季风气候,年平均气温为159~166℃,无霜期242~263d,年均日照时数1800~2000h,年降水量1100~1300mm。
湖北林业科技第46卷第6期况红玲,等:不同浓度CaCl2对宜昌润楠幼苗的抗寒性研究1.2试验设计
试验于2016年3月~2016年10月进行。2016年4月初,取盆景园内长势一致且无病虫害的宜昌润楠1a生苗,移栽入营养钵内,放入温室常规管理三个月后,每隔2d对材料喷施CaCl2,3次重复,4个处理:处理1为清水,处理2为5mmol/LCaCl2,处理3为15mmol/LCaCl2,处理4为25mmol/LCaCl2,每处理5株苗木,清水作对照,最后一次试剂处理48h后,将材料分别放入5,0,-5℃人工智能培养箱内培养48h后测定抗寒性指标。
1.3测定方法
叶绿素含量测定参考王学奎方法[6];丙二醛含量的测定采用王学奎方法[6];蛋白质含量的测定采用王学奎的考马斯亮蓝G-250法[6];植物组织可溶性糖测定参考王学奎的蒽酮比色法[6];超氧化物歧化酶活力的测定参考王学奎的氮蓝四唑法[6];过氧化物酶活性的测定采用王学奎的愈创木酚法[6];激素ABA含量的测定参考彭运生方法[7]。
1.4数据处理
实验数据使用SPSS18.0进行统计分析,试验数据均用平均值±标准误表示(mean±SEM)。采用单因素方差分析和Tukey多重检验,显著差异设置为P>005。
2结果與分析
2.1不同浓度CaCl2对宜昌润楠幼苗叶绿素的影响
图1多重比较结果显示,宜昌润楠幼苗叶绿素总含量在5℃时处理1、处理2极显著高于处理4,较显著高于处理3;在0℃时处理1极显著高于处理3,较显著高于处理2、处理4,而处理2、处理4互相不显著;在-5℃时处理2极显著高于处理3、处理4,较显著高于处理1,而处理3和处理4互相不显著。综上所述,不同浓度CaCl2对宜昌润楠幼苗叶绿素总含量影响中-5℃条件下处理2效果最显著。
2.2不同浓度CaCl2对宜昌润楠幼苗丙二醛(MDA)的影响图2多重比较结果显示,宜昌润楠幼苗丙二醛(MDA)含量在5℃时处理1较显著高于处理2、处理3、处理4,而处理2、3、4之间两两差异不显著;在0℃时处理1极显著高于处理4,显著高于处理3、较显著高于处理2;在-5℃时处理1、3、4极显著高于处理2,而处理1、3、4之间两两差异不显著。综上所述,不同浓度CaCl2对宜昌润楠幼苗丙二醛含量影响中-5℃条件下处理1最显著。
2.3不同浓度CaCl2对宜昌润楠幼苗可溶性蛋白质含量的影响图3多重比较结果表明,宜昌润楠幼苗可溶性蛋白含量在5℃时处理1极显著高于处理2、3、4,而处理2、3、4之间两两差异不显著;在0℃时处理4极显著高于处理3,较显著高于处理1、处理2,而处理1、处理2互相不显著;在-5℃时处理4极显著高于处理2,较显著高于处理1、3,而处理1和处理3互相不显著。综上所述,不同浓度CaCl2对宜昌润楠幼苗可溶性蛋白含量影响中5℃条件下处理1效果最显著。
2.4不同浓度CaCl2对宜昌润楠幼苗可溶性糖含量的影响图4多重比较结果表明,宜昌润楠幼苗可溶性糖含量在5℃时处理1极显著高于处理4,显著高于处理3、较显著高于处理2;在0℃时处理1、2极显著高于处理4,较显著高于处理3,而处理1、处理2互相不显著;在-5℃时处理4极显著高于处理2,较显著高于处理1、3,而处理1和处理3互相不显著。综上所述,不同浓度CaCl2对宜昌润楠幼苗可溶性糖含量影响中-5℃条件下处理4效果最显著。
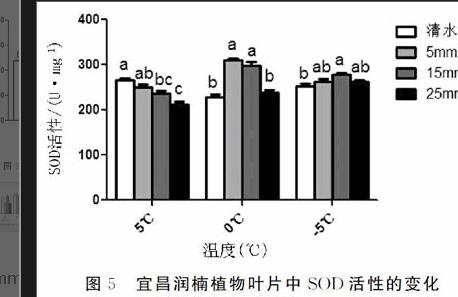
2.5不同浓度CaCl2对宜昌润楠幼苗超氧化物歧化酶(SOD)活性的影响图5多重比较结果显示,宜昌润楠幼苗超氧化物歧化酶SOD活性在5℃时处理1极显著高于处理4,显著高于处理3、较显著高于处理2;在0℃时处理2、3极显著高于处理1、2,而处理1、4互相不显著,处理2、3互相不显著;在-5℃时处理3极显著高于处理1,较显著高于处理2、4,而处理2和处理4互相不显著。综上所述,不同浓度CaCl2对宜昌润楠幼苗超氧化物歧化酶SOD活性影响中0℃条件下处理2效果最显著。
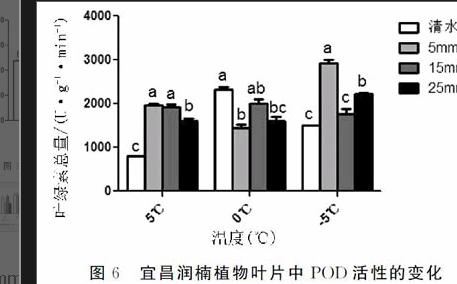
2.6不同浓度CaCl2对宜昌润楠幼苗超过氧化物酶(POD)活性的影响图6多重比较结果显示,宜昌润楠幼苗过氧化物酶POD活性在5℃时处理2、3极显著高于处理1,较显著高于处理4,而处理2、3互相不显著;在0℃时处理1极显著高于处理2,显著高于处理4,较显著高于处理3;在-5℃时处理2极显著高于处理1、3,较显著高于处理4,而处理1和处理3互相不显著。综上所述,不同浓度CaCl2对宜昌润楠幼苗过氧化物酶POD活性活性影响中-5℃条件下处理2效果最显著。
2.7不同浓度CaCl2对宜昌润楠幼苗激素ABA含量的影响图7多重比较结果表明,宜昌润楠幼苗ABA含量在5℃时处理4极显著高于处理1、2、3,而处理1、2、3两两差异不显著;在0℃时处理3极显著高于处理2、4,较显著高于处理1,而处理2和处理4互相不显著;在-5℃时处理1、2极显著高于处理4,较显著高于处理3,而处理1、2互相不显著。综上所述,不同浓度CaCl2对宜昌润楠幼苗ABA含量影响中-5℃条件下处理2效果最显著。endprint
3结论与讨论
3.1外源CaCl2对叶绿素含量的影响
叶绿素是植物光合作用必需色素。Allen曾报道叶绿素是光合作用的物质基础,同时也是光合作用的光敏化剂[8]。低温胁迫时植物叶片中叶绿素会发生变化,叶绿素的合成受到影响,同时加剧叶绿素的降解。这在黄瓜[9]、甜菜[10]以及卫矛科三种常绿植物[11]等研究中均见报道。
本文研究结果显示,5℃和-5℃条件下的宜昌润楠幼苗随着CaCl2浓度的增加,叶绿素总量呈先增后减的趋势,暗示在低温胁迫下,一方面CaCl2的处理具有时间依赖性,仅在其喷施初期具有保护性作用;另一方面可能由于CaCl2喷施不能有效的抵消低温对叶绿素的损伤。同时,在所有处理中-5℃条件下处理2叶绿素含量最高。
3.2外源CaCl2对丙二醛含量的影响
在正常情况下,植物细胞内活性氧的产生和清除之间保持着一个动态的平衡,能使细胞免受到活性氧的损伤。在低温胁迫下,植物体内的O-自由基清除能力下降,细胞中积累大量的活性氧,MDA的代谢平衡状态被破坏,MDA的大量积累进而造成细胞损伤,MDA可以抑制细胞保护酶的活性,同时能降低抗氧化物的含量,使细胞膜脂过氧化的发生加剧[12]。王华等研究显示,植物体内MDA的含量反映了抗寒性的强弱,且二者呈负相关性[13]。
本文对低温胁迫下宜昌润楠幼苗的MDA的含量进行测定,结果表明在5℃和0℃条件下,宜昌润楠幼苗中MDA含量随CaCl2浓度增加MDA的含量逐渐下降。在所有处理中5℃条件下处理4中MDA含量最低。产生以上现象有以下几种解释,第一种可能是过低低温胁迫导致润楠植物叶片细胞受损,即使通过CaCl2的处理不能改善低温胁迫对细胞的损伤,进而植物中MDA的含量呈现出上升趋势,这与李燕萍[14]在CaCl2调节油茶抗寒性的影响研究结果类似。第二种可能由于低温胁迫导致植物细胞膜的通透性发生改变,大量Ca2+离子进入,使磷脂酶A2的活性增加,进一步在酶促反应作用下形成羟基化合物最终分解为MDA,同时在MDA和羟基化合物的作用下,使得细胞膜的通透性增大,又使得大量Ca2+离子进入,发生恶性循环[15]。
3.3外源CaCl2对可溶性蛋白含量的影响
可溶性蛋白是细胞内渗透调节物质,可增加细胞渗透性,降低冰点,增加植物的耐寒力。当低温伤害较轻时,植株会通过自身的生理调控来抵御外界逆境胁迫,而持续的严重低温胁迫会带来不可逆的伤害,从而导致生理失调,植株表現出冷害症状乃至死亡。彭艳华等研究表明,低温胁迫下可溶性蛋白含量升高,有利于蛋白质的合成,提高对低温胁迫的抗性[16]。同时,低温胁迫能激活大量蛋白基因,进而产生大量新合成抗寒性蛋白[17]。
本文研究结果显示,0℃条件下,处理4中宜昌润楠幼苗可溶性蛋白质含量高于其对照组处理1。在-5℃条件下,处理4中可溶性蛋白质含量高于其对照组处理1。可能暗示了低温胁迫下通过诱导抗寒基因表达进而使蛋白上升。这与前人的报道在植物茄[18]和毛白杨[19]中的报道具有一致性,CaCl2处理能够促进抗寒过程中可溶性蛋白质的合成,使可溶性蛋白质的含量增加。
3.4外源CaCl2对可溶性糖含量的影响
低温胁迫条件下,可溶性糖积累与植物细胞液浓度密切相关,一方面液泡中糖的积累能增加组织中非结冰水,减缓细胞质脱水过度,使细胞不会遇冷凝固,进而使植物的抗寒性提高[20]。另一方面可溶性糖对线粒体及膜上偶联因子均有保护作用,大部分植物中可溶性糖含量与其自身抗寒性成正相关。大量研究表明随着温度胁迫的加强可溶性糖含量逐渐增加,暗示可溶性糖是植物抗寒性的一种主要保护性物质。
本文研究结果表明,在所有处理中-5℃条件下处理4中宜昌润楠幼苗可溶性糖含量最高。在5℃条件下,经不同浓度CaCl2处理后宜昌润楠幼苗各处理组间可溶性糖含量都低于其对照组,这与前人的研究相符[21],推测可能由于在该低温温度胁迫下植物的呼吸作用较强,致使可溶性糖含量下降,或者通过糖的代谢作用提高能量,产生保护性物质,维系植物细胞的正常生理功能。
3.5外源CaCl2对SOD和POD活性的影响
学者Fridovich在1975年提出细胞保护酶系统这一概念,认为分布十分广泛的SOD和POD是细胞中最重要的抗氧化酶[22]。在低温胁迫条件下,植物细胞膜脂发生氧化和过氧化损伤。而SOD和POD能清除O2-和过氧化物,减少或阻止羟基自由基的发生,减轻膜脂的过氧化水平,对细胞膜起到重要的保护作用,进而降低低温胁迫对植物的损伤[23],提高植物的抗寒性能力。
本文研究结果显示,低温胁迫条件下,在5℃条件下,伴随CaCl2浓度的增加,宜昌润楠幼苗的SOD活性逐渐降低。在0℃条件下,宜昌润楠幼苗的SOD活性随着CaCl2浓度的增加出现上升的趋势,处理2活性达到最高。同样,在5℃条件下,经CaCl2处理后的宜昌润楠幼苗POD活性增加,并且在所有处理中-5℃条件下处理2中POD活性最高。暗示宜昌润楠幼苗在低温胁迫条件下,通过喷施CaCl2能提高SOD、POD的活性,清除低温胁迫产生的不利损伤。这与低温胁迫条件下Ca2+处理其他植物的研究具有一致性[24-30]。康国章等学者研究表明Ca2+能提高植物抗寒能力主要通过两个途径实现。一方面提高保护酶(SOD、POD)活性、细胞膜脂结构和细胞壁的稳定性来增加抗寒性;另一方面可能是在分子水平上,植物能感知低温并诱导其抗冻基因的mRNA的大量表达提高植物的抗寒能力[31]。
3.6外源CaCl2对激素ABA含量的影响
植物激素对控制植物生长发育具有重要的功能,在其生物过程中发挥关键作用[32],广泛参与逆境条件下各种生理的调节[33]。内源激素ABA在植物体内的变化与植物抗性之间存在正相关性[34],能特征植物在胁迫条件下的响应。前人研究报道在低温胁迫条件下,低温胁迫加强植物体内ABA大量积累,不同植物间ABA含量不同,抗寒性越强植物体内ABA含量越高[35]。本文研究结果显示:在5℃条件下,喷施不同浓度CaCl2后宜昌润楠幼苗的ABA含量总体有所增加,这与Iqbal等在小麦种的研究结果相符[36]。喷施CaCl2后能增加植物中ABA含量,加强了植物对低温胁迫的抗应激能力,而在所有处理中-5℃条件下处理2中宜昌润楠幼苗的ABA含量最高。endprint
综上所述,宜昌润楠经不同浓度CaCl2处理后,综合测定指标考虑,5mmol/LCaCl2对提高宜昌润楠抗寒性效果显著,此研究为润楠属的地方引种栽培奠定了基础。
参考文献
[1]中国植物志编委会.中国植物志(第31卷)[M].北京:科学出版社,1982:768.
[2]丁宝章,王遂义.河南植物志[M].郑州:河南科学技术出版社,1978:2021.
[3]孙守家.玫瑰花蕾采后衰老机理研究[D].山东泰安:山东农业大学,2003.
[4]LyonsJM.Chillinginjuryinplants.AnnRevPlantPhysiol[J].1973,24:445466.
[5]SteponkusPL,UemuraM,JosephRA,etal.ModeofactionoftheCOR15ageneonthefreezingtoleranceofArabidopsisthaliana[J].ProceedingsoftheNationalAcademyofSciences,1998,95(24):1457014575.
[6]王学奎.植物生理生化实验原理和技术(第2版)[M].北京:高等教育出版社,2006:3132.
[7]彭运生,凌祖铭.高效液相色谱测定水稻幼苗中内源激素(GA3,IAA和ABA)的方法研究[J].分析测试通报,1992,11(2):5255.
[8]AllenDJ,OrtDR.Impactsofchillingtemperaturesonphotosynthesisinwarmclimateplants[J].TrendsinPlantScience,2001,6(1):3642.
[9]马德华,卢育华,庞金安.低温对黄瓜幼苗膜脂过氧化的影响[J].园艺学报,1998,25(1):6164.
[10]和红云,薛琳,田丽萍,等.低温胁迫对甜瓜幼苗叶绿素含量及荧光参数的影响[J].北方园艺,2008(4):121127.
[11]吴娜.卫矛科三种常绿阔叶植物抗寒性研究[D].保定:河北农业大学图书馆,2006.
[12]陈少裕.脂膜过氧化对植物细胞的伤害[J].植物生理学通讯,1991,27(2):8490.
[13]王华,王飞,陈登文,等.低温胁迫对杏花SOD活性和膜脂过氧化的影响[J].果树科学,2000,17(3):197201.
[14]李艳萍.油茶品种的耐寒性评价及CaCl2对油茶抗寒性的影响[D].南京:南京林业大学,2012.
[15]刘剑锋,张红艳,彭抒昂.梨采后果实质地与Ca2+,果胶,丙二醛,乙烯动态的关系[J].华中农业大学学报,2003,3:270273.
[16]彭艳华,刘成运,卢大炎,等.低温胁迫下凤眼莲叶片的适应[J].武汉植物学研究,1992,10(2):123127.
[17]刘祖祺.ABA/Gas调空特异蛋白质与柑橘的抗寒性[J].园艺学报,1993,20(4):335340.
[18]高洪波,陈贵林.钙调素拮抗剂和钙对茄子嫁接苗抗冷性的影响[J].园艺学报,2002,29(3):243246.
[19]LinSZ,ZhangZY.RoleofCaCl2incoldacclimationinducedfreezingresistanceofPopulustomentosaCuttings[J].ForestrystudiesinChina,2002,4(2):3842.
[20]张石成.植物的抗寒生理[M].北京:农业出版社,1990:8182.
[21]苗永美,王万洋,杨海林,等.外源Ca2+,SA和ABA缓解甜瓜低温胁迫伤害的生理作用[J].南京农业大学学报,2013,36(4):2529.
[22]Fridovich.TheBiologyofOxgenRedical[J].Seience,1975,201:875880.
[23]杨玲,袁月星,谢双琴.次适温下水杨酸浸种对水稻种子萌发的效应[J].植物生理学通讯,2001,37(4):288290.
[24]张文静,刘亮,黄正来,等.低温胁迫对稻茬小麦根系抗氧化酶活性及内源激素含量的影响[J].麦类作物学报,2016,36(4):501506.
[25]余土元,冯颖竹.低温对香蕉幼苗的伤害及外源H2O2,CaCl2对香蕉抗寒性的影响[J].中国农学通报,2011,27(25):219223.
[26]李君可,王静,王利鹃,等.预喷施CaCl2溶液对低温胁迫下烤烟幼苗生理生态的影响[J].2013,35(11):5461.
[27]李昳乐,吴军,尚忠海.低温胁迫及CaCl2处理对青竹复叶槭抗寒指标的影响[J].北京农业学院学报,2008,23(4):4749.
[28]彭向永,徐永茹,唐萍,等.外源钙对低温胁迫下甜瓜幼苗膜稳定性和渗透调节物质的影响[J].曲阜师范大学学报,2011,37(1):8184.
[29]张琳,王甲辰,左强,等.外源钙对低温胁迫下番茄幼苗生化指标及膜伤害的研究[J].北方园艺,2011(12):2426.
[30]严寒,冉茜,何勇,等.低溫胁迫下马蹄金幼苗对外源水杨酸的生理响应[J].江苏农业学报,2009,25(5):11881189.
[31]葛辛.高级植物分子生物学[M].北京:科学出版社,2004:123145.
[32]张雪峰.低温胁迫对玉米种子萌发过程中内源激素含量变化的影响[J].沈阳农业大学学报,2011,42(2):147151.
[33]罗立津,徐福乐,翁华钦,等.脱落酸对甜椒幼苗抗寒性的诱导效应及其机理研究[J].西北植物学报,2011,34(1):94100.
[34]LangLV,MantylaE,WelinB,etal.Alterationsinwaterstatus,endogenousabscisicacidcontent,andexpressionofrabl8geneduringthedevelopmentoffreezingtoleranceinArabidopsisthaliana[J].PlantPhysiol,1994,104:13411349.
[35]王兴,于晶,杨阳,等.低温条件下不同抗寒性冬小麦内源激素的变化[J].麦类作物学报,2009,29(5):827831.
[36]IqbalM,AshrafM,RehmanS,etal.Doespolyamineseedpretreatmentmodulategrowthandlevelsofsomeplantgrowthregulatorsinhexaploidwheat(TriticumaestivumL.)plantsundersaltstress[J].BotanicalStudies,2006,47:239250.(责任编辑:夏剑萍)endprint
