西番莲不同种质资源抗寒性测定
2022-06-06郭玉琳吴凤禅李安定蔡国俊
郭玉琳 吴凤禅 李安定 蔡国俊




摘 要:为筛选贵州抗寒性强的西番莲品种,设定5个低温处理梯度,测定不同低温胁迫下6个品种西番莲枝条膜脂特性、可溶性物质、细胞保护酶,分析其变化趋势,并通过隶属函数法综合评价各品种抗寒能力。研究结果表明,所有品种枝条相对电导率均随处理温度降低而增加,‘蓝香西番莲’()、‘玛格丽特西番莲’(‘’)、‘贵寒1号’()半致死温度较其他低,各品种MDA变化趋势差异较大,总体上–20℃下MDA含量高于5℃处理,‘玛格丽特西番莲’‘贵寒1号’‘台农1号’‘巨无霸黄金果’枝条可溶性糖随处理温度降低呈下降趋势,各西番莲品种枝条可溶性蛋白质含量随处理温度降低呈先增加后减少趋势,除‘巨无霸黄金果’外,其他品种枝条CAT酶活性均随处理温度下降呈先上升后下降趋势,枝条POD酶活性随处理温度下降呈先上升后下降趋势,各品种枝条SOD酶活性随温度变化趋势差异较大。根据隶属函数法判定6个西番莲品种均为Ⅱ级,中抗寒性品种,得出各品种抗寒性强弱顺序为:‘贵寒1号’>‘蓝香西番莲’>‘玛格丽特西番莲’>‘台农1号’>‘紫花西番莲’()>‘巨无霸黄金果’。
关键词:西番莲;抗寒性;生理特性;隶属函数法中图分类号:S65 文献标识码:A
is an important fruit to raise peasants’ income in rough country in subtropical China. Cold resistance of is crucial to keep survival rates, quality and yield. In order to screen strong cold resistance varieties in Guizhou, we set five low temperature gradients, 5℃, 0℃, ?5℃, ‒10℃, ‒20℃, for 6 varieties from Pingtang, Guizhou, China, and determined the membrane lipid characteristics, soluble substances, and cell protective enzymes, and then analyzed the variation trend of above indictors under different temperature treatment. Finally we used the membership function method to evaluate the cold resistance capacity of comprehensively. The results showed the relative electric conductivity increased with decreasing temperature (mean relative electric conductivity of the varieties 35.19% in 5℃ treatment and 89.71% in 20℃ treatment), the semilethal temperature of ,‘’ and was lower than that of other varieties (‒1.426‒0.98℃). We also found the variation trends of intracellular malondialdehyde in plants (MDA) were different among varieties, and the MDA content under ‒20℃ treatment was generally higher than that under 5℃ treatment. When temperature decreased, the soluble sugar of majority varieties declined, the descent range of ‘QueenJuwuba’ was the highest (59.24%) while that of ‘Tainong’was the lowest (31.26%). Soluble protein content in the branches of all varieties increased first and then declined with decreasing treatment temperature. Catalase (CAT) activity was processed with temperature drop of all varieties except ‘QueenJuwuba’. Variation trend of peroxidase (POD) enzyme activity was the same as CAT. However, the variation of superoxide dismutase (SOD) enzyme activity with temperature was different among varieties. Through the membership function method, we identified the cold resistance of the varieties was in II level, its ranking with membership was: ‘GuiHanNo.1’ (0.543) > (0.542)>‘’ (0.521)> ‘Tainong’(0.501)> (0.489)> ‘QueenJuwuba’(0.459). The results indicated the cold-resistant of ‘GuiHanNo.1’ and was the strongest among the six varieties, while the cold-resistant of ‘QueenJuwuba’ was the poorest. The cold resistance should be improved for all six varieties.
; cold resistance; physiology properties; membership function
: 10.3969/j.issn.1000-2561.2022.05.010
植物的生长发育除了依靠自身的生理调节外,还受外部环境影响,其中低溫胁迫是重要的外部影响因素之一。低温分为冷害和冻害,冷害指0~10℃对植物造成的伤害,冻害指0℃以下温度对植物造成的伤害,低温不仅是影响植物自然地理分布的主要环境条件,也是制约农林生产活动的重要因素,当外界环境温度变低时,植物体内的生理调节也会随之变化以减轻低温对植物造成的伤害,植物细胞膜系统是最先受低温胁迫影响的部位,低温会破坏植物细胞膜系统,导致溶质外渗,引起细胞电导率发生改变,因此,植物抗寒性强弱与电导率变化密切相关,除此之外,植物细胞内丙二醛(MDA)含量与植物细胞质膜损伤程度呈负相关,可据此判定植物抗寒性强弱。
低温胁迫下,细胞内可溶性糖可增强细胞保水能力,保护细胞器和生物膜,为新陈代谢提供能量。可溶性蛋白也具增强细胞保水能力、调控抗寒基因等作用,通过研究低温胁迫下植物可溶性糖和蛋白的变化分析植物抗寒性在葡萄等多种农作物上已得到广泛应用。
酶作为植物体内抗氧化防御系统的重要物质,低温胁迫下可保持清除活性氧自由基的作用,调节细胞膜透性,防止膜损伤,超氧化物歧化酶(SOD),过氧化物酶(POD),过氧化氢酶(CAT)是研究得最多的保护酶系统,抗寒性强的植物在低温下仍能维持较高的酶活性水平。目前,以上植物生理指标在多种经济作物抗寒品种(如水稻、小麦等)筛选中均有较充分的研究,但关于西番莲生理指标对低温胁迫的响应及抗寒性判定研究较少。
西番莲()原产于热带地区,目前主要栽培于我国广西、云南等地。贵州属于喀斯特地貌,石漠化问题严重,西番莲作为常绿攀岩型植物,具有良好的植被覆盖能力及经济效益,是防治喀斯特地区石漠化防的理想栽培作物。西番莲种植平均气温为18℃以上,而贵州省常年温度为16℃左右,且冬季贵州大部分地区温度在0℃左右,导致西番莲不能顺利越冬,因此,选取抗寒性较强的西番莲品种极为迫切。本研究主要通过设定不同低温处理,测定‘贵寒1号’等6个品种的西番莲枝条膜脂特性、可溶性物质、细胞保护酶类对低温处理的响应,旨在选出抗寒性较强的西番莲品种,通过本研究,可以为喀斯特地区西番莲引种栽培和培育提供理论依据。
材料与方法
材料
试验材料为西番莲一年成熟标准株枝条,材料全部采自贵州省平塘种植基地,分别有‘贵寒1号’()、‘巨无霸黄金果’、‘玛格丽特西番莲’(‘’)、‘台农1号’、‘蓝香西番莲’()、‘紫花西番莲’()6个品种。
方法
1.2.1 半致死温度测定 对各株系西番莲一年生枝条进行室内人工低温试验。将每个品种的西番莲枝条剪成数段,分成6份,将枝条冲洗干净,并用吸水纸吸干枝叶表面水分,然后用自封袋保存并放入人工智能培养箱(温度控制精度为±0.1℃,用水银温度计进行校准)进行不同梯度温度处理,低温分别设置为5、0、–5、–10、–20℃,待温度稳定后维持16 h后在室温中解冻,放入50 mL的三角瓶中,加人40 mL重蒸馏水,用真空干燥器抽20~30 min,摇匀后用电导仪测出电导率(),代表低温处理的电解质渗出量。再将其放入蒸锅中(用保鲜膜封瓶口)蒸30 min后,测得不同植物部位被杀死后的电导率(),代表处理材料的原生质膜全部破坏后所渗出电解质的总量。以上实验3次重复,以浸泡液的导电率为对照()。电解质渗出率()=‒‒将测得的电导率曲线进行非线性回归分析,采用Logistic回归方程拟和,Logistic回归方程的表达式为:=/(1+)。=电解质渗出率,=处理温度,=细胞伤害率和饱和容量,为方程参数。对和进行回归分析,得出的数值,由Logistic求取二级导数得出=ln,此时值即半致死温度(LT)。
1.2.2 枝条MDA、可溶性糖、可溶性蛋白、SOD、POD、CAT测定 采用硫代巴比妥酸法测定丙二醛(malondialdehyde, MDA)含量,蒽酮法测定可溶性糖(soluble sugar, SS)含量,考马斯亮蓝G-250法测定可溶性蛋白(soluble protein, SP)含量,氮蓝四唑(NBT)法测氧化物歧化酶(superoxide dismutase, SOD)活性,愈創木酚法测过氧化物酶(peroxidase, POD)活性,过氧化氢还原法测过氧化氢酶(catalase, CAT)活性,每个指标重复测3次,结果取平均值。
数据处理
数据计算、制图等采用Excel软件处理,SPASS软件进行相关系数分析。
结果与分析
西番莲枝条膜脂特性对低温胁迫的响应
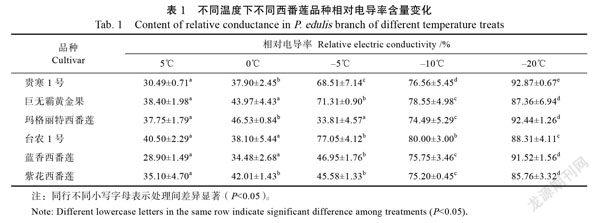
由表1可知,随着温度的降低,所有品种的西番莲相对电导率逐渐上升,5℃时,‘贵寒1号’电导率最低,‘台农1号’电导率最高,当温度下降至0℃时,所有品种的相对电导率变化趋势不大,当温度下降至‒5℃时,‘贵寒1号’‘巨无霸黄金果’‘台农1号’的电导率显著上升,电导率在70%左右,说明枝条受到了伤害,其中‘台农1号’电导率最高,‘贵寒1号’电导率最低;其余品种变化电导率均在50%以下,说明枝条受到的伤害要小于上述5个品种,当温度下降至‒10℃时,‘玛格丽特西番莲’‘蓝香西番莲’‘紫花西番莲’的电导率显著上升,说明在‒5~‒10℃温度区间内,对该品种的枝条伤害较大;在‒10~‒20℃时,6个西番莲品种的电导率持续上升,相对电导率均在85%~93%之间,说明该温度区间下6个品种的西番莲枝条已经受到最大伤害。
根据在不同温度条件下测定的6个西番莲品种枝条的相对电导率可求得其Logistic方程、低温半致死温度及拟合度,求取结果见表2。6个西番莲品种的拟合度在0.903~0.977之间,拟合度较高,说明数据可信度高,从Logistic方程统计结果可以看出,6个西番莲品种的半致死温度范围为0.980~‒3.359之间,半致死温度最低为‘蓝香西番莲’,最高为‘台农1号’,通过半致死温度数值对这6个品种的抗寒性进行排序,结果为:‘蓝香西番莲’>‘紫花西番莲’>‘玛格丽特西番莲’>‘贵寒1号’>‘巨无霸黄金果’>‘台农1号’。
一般认为,MDA含量积累越多,机体中清除自由基的能力越差,组织自我保护能力越弱。由图1可知,5℃处理下各品种MDA含量最低,相对5℃处理,0℃处理后5个品种枝条MDA含量上升幅度显著,‘贵寒1号’‘紫花西番莲’‘台农1号’‘蓝香西番莲’MDA含量均在0℃处理时达到最大,说明在0℃处理下‘贵寒1号’‘紫花西番莲’‘台农1号’‘蓝香西番莲’‘巨无霸黄金果’枝条细胞质膜已严重受损,‘玛格丽特西番莲’枝条MDA含量随处理温度降低缓
慢上升,在‒10℃处理下达到最高。
西番莲枝条可溶性物质对低温胁迫的响应
随处理温度降低,‘玛格丽特西番莲’‘贵寒1号’‘台农1号’‘巨无霸黄金果’枝条可溶性糖含量均呈下降趋势(图2)。‘蓝香西番莲’与‘紫花西番莲’枝条可溶性糖含量变化趋势相似,在5~‒5℃处理下降低,‒10℃处理下略有回升,所有品种枝条可溶性糖含量最高值均出现在5℃处理时,最低值出现在‒20℃处理,且最高值与最低值差异极显著,‘巨无
霸黄金果’降幅最大(59.24%),‘台农1号’降幅最小(31.26%)。
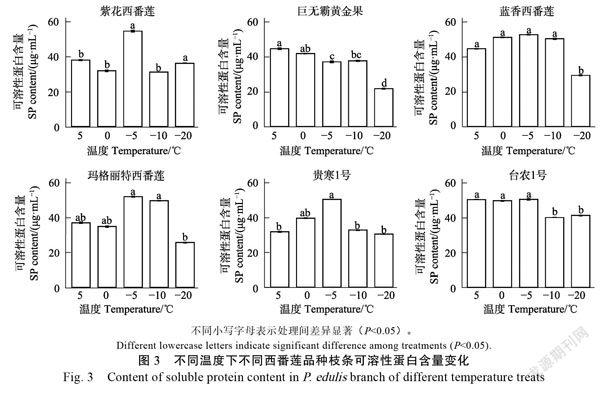
除‘巨无霸黄金果’外,各西番莲品种枝条可溶性蛋白质含量随处理温度降低呈先增加后减少趋势(图3),且‒5℃处理下,除‘巨无霸黄金果’外的其他品种枝条可溶性蛋白含量都处于较高水平(50.68~55.00 μg/mL),‘巨无霸黄金果’枝条可溶性蛋白质含量随处理温度降低呈下降趋势。
西番莲枝条细胞保护酶类对低温胁迫的响应
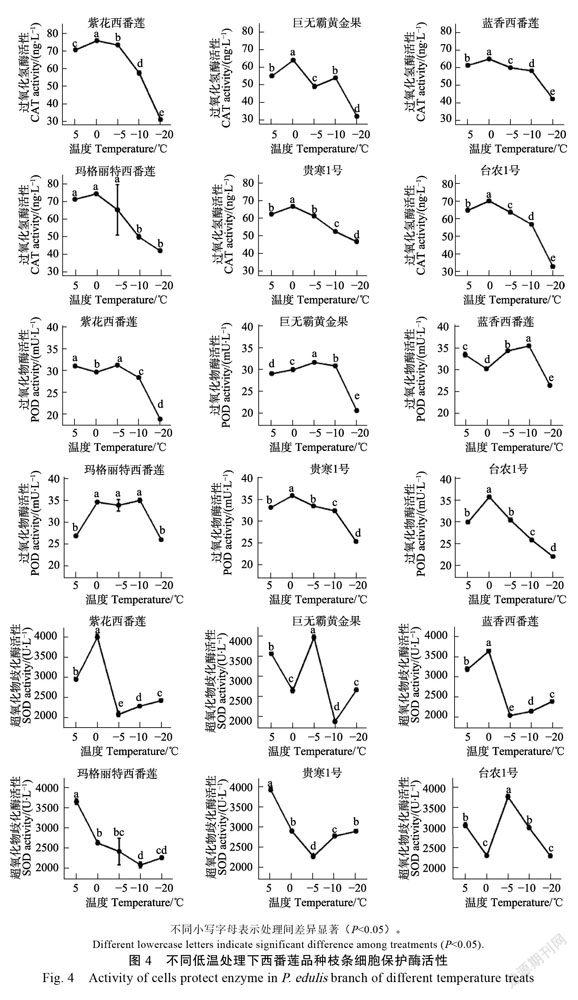
除‘巨无霸黄金果’外,其他品种枝条CAT酶活性均在5~0℃处理间上升,随后随处理温度下降呈下降趋势(图4),‘蓝香西番莲’‘玛格丽特西番莲’和‘贵寒1号’下降趋势较缓。
‘紫花西番莲’‘巨无霸黄金果’‘玛格丽特西番莲’‘蓝香西番莲’‘贵寒1号’枝条POD酶活性在‒10~‒20℃降幅最大(21.52%~33.77%),5~ ‒10℃间变化幅度较小(2.40%~8.58%),‘台农1号’枝条酶活性在0℃处理时达到最高值,随后随处理温度降低基本呈直线下降趋势。
枝条SOD酶活性变化与温度关系较复杂,‘紫花西番莲’和‘蓝香西番莲’枝条SOD酶活性随处理温度降低呈先上升后下降随后上升趋势,最低值出现在‒5℃处理下,‘贵寒1号’枝条SOD酶活性最低值也出现在‒5℃处理下,但随处理温度降低,其变化趋势为先降低后上升,‘台
农1号’和‘巨无霸黄金果’枝条SOD酶活性在‒5℃处理下达到最大值,‘玛格丽特西番莲’枝条SOD酶活性则随处理温度降低呈下降趋势,当温度小于‒10℃时,其值有回升,但与‒10℃下的值差异不显著。
隶属函数法对西番莲抗寒性综合评价
参照董万鹏等、姜丽娜等的隶属函数法及对抗寒性划分方法,对各指标进行抗寒性判定,6个品种均为Ⅱ级,中抗寒性品种,其平均隶属度由大到小为:‘贵寒1号’>‘蓝香西番莲’>‘玛格丽特西番莲’>‘台农1号’>‘紫花西番莲’>‘巨无霸黄金果’(表3)。
讨论
低温胁迫下,西番莲枝条膜脂特性、可溶性物质、细胞保护酶类均随不同温度处理产生不同变化趋势。枝条相对电导率均随处理温度降低而增加,低温易使细胞间隙中的水分结冰,产生压力,将细胞内的水分向间隙中挤压,导致原生质脱水,从而增大细胞膜透性,相对电导率增加。‘蓝香西番莲’‘紫花西番莲’‘玛格丽特西番莲’相对电导率在‒5℃处理下均处于较低水平,当处理温度小于‒10℃时,各品种电导率差异小,说明‘蓝香西番莲’‘紫花西番莲’‘玛格丽特西番莲’细胞膜透性抗性较其他品种强,当温度低于‒10℃时,西番莲枝条细胞膜受损严重。由相对电导率推算的各品种半致死温度结果显示,‘蓝香西番莲’‘玛格丽特西番莲’‘贵寒1号’半致死温度较低(分别为‒3.359℃,‒2.384℃,‒1.426℃),抗寒性强,其中‘贵寒1号’半致死温度较董万鹏等研究的‘贵寒1号’半致死温度高(‒2.857℃)。
细胞膜受到逆境伤害时,膜脂产生过氧化作用,生成MDA,因此MDA与相对电导率表征作用一致,低温下枝条MDA含量越大,细胞膜质过氧化程度越严重。相对其他品种来说,‘玛格丽特西番莲’枝条MDA含量随温度降低上升幅度较小,说明其抗寒性强。
可溶性糖增加利于提高细胞液浓度,增强植物抗寒性,有研究认为温度下降促进植物可溶性糖积累,抗寒性越强的物质细胞内可溶性糖含量增加幅度越大;本研究中‘蓝香西番莲’与‘紫花西番莲’枝条可溶性糖与其余品种相比含量在‒10℃处理下略有回升,说明‘蓝香西番莲’和‘紫花西番莲’抗寒性较其他品种强。可溶性蛋白促进细胞持水力增强,可减少因原生质结冰造成的病害,本研究中除‘巨无霸黄金果’外,各西番莲品种枝条可溶性蛋白质含量随处理温度降低呈先增加后减少趋势,且‒5℃处理下,除‘巨无霸黄金果’外的其他品种枝条可溶性蛋白含量都处于较高水平(50.68~55.00 μg/mL),说明‘巨无霸黄金果’相较其余品种抗寒性较差,董万鹏等的研究也发现随处理温度降低西番莲可溶性蛋白呈先上升后下降趋势,所得结果一致。
超氧歧化酶和过氧化物酶同工酶可作为参与植物抗逆反应的催化剂,调节膜透性和防止膜的损伤,当植物处于低温胁迫时,SOD、CAT、POD三种膜保护酶相互协调共同提高植物体抗寒的能力,一般认为,酶活性越强越利于植物抗寒。除‘巨无霸黄金果’外,其他品种枝条CAT酶活性均随处理温度下降呈先上升后下降趋势,‘蓝香西番莲’‘玛格丽特西番莲’和‘贵寒1号’下降趋势较缓。‘紫花西番莲’‘玛格丽特西番莲’‘蓝香西番莲’‘贵寒1号’枝条POD酶活性降幅较小(2.40%~8.58%),枝条SOD酶活性变化与温度关系较复杂。‘蓝香西番莲’‘玛格丽特西番莲’和‘贵寒1号’低温下有较强的酶活性,利于抗寒。
根据隶属函数法判定6个西番莲品种均为Ⅱ级,中抗寒性品种,其平均隶属度由大到小为:‘贵寒1号’>‘蓝香西番莲’>‘玛格丽特西番莲’>‘台农1号’>‘紫花西番莲’>‘巨无霸黄金果’,‘贵寒1号’‘蓝香西番莲’‘玛格丽特西番莲’与抗寒性呈正相关的生理指标(如CAT、POD)均表现为同一低温处理下较其他品种更高的值,或下降趋势较缓,与抗寒性呈负相关的生理指标(如MDA)则表现为同一低温处理下较其他品种更低的值,隶属度结果与各生理指标测定结果吻合。董万鹏等的研究也表明‘贵寒1号’较其他西番莲品种有更强的抗寒性。
结论
本研究结果表明,6个西番莲品种均为Ⅱ级,中抗寒性品种,得出各品种抗寒性强弱顺序为:‘贵寒1号’>‘蓝香西番莲’>‘玛格丽特西番莲’>‘台农1号’>‘紫花西番莲’>‘巨无霸黄金果’。
参考文献
[4] 王 芳, 王 淇, 赵曦阳, 低温胁迫下植物的表型及生理响应机制研究进展[J]. 分子植物育种, 2019, 17(15): 5144-5153.WANG F, WANG Q, ZHAO X Y. Research progress of phenotype and physiological response mechanism of plants under low temperature stress[J]. Molecular Plant Breeding, 2019, 17(15): 5144-5153. (in Chinese)
[5] 刘 冰, 王有科. 应用Logistic方程确定花椒枝条低温半致死温度[J]. 甘肃农业大学学报, 2015(4): 57-61.LIU B, WANG Y K. Applieaiton of logisitic quaiton on determination of lehtal dose-50 temperature of shoots[J]. Journal of Gansu Agricultural University, 2015(4): 57-61. (in Chinese)
[6] 柴弋霞, 曾 雯, 金晓玲, 蔡梦颖. 铁冬青雌雄株在低温胁迫下的响应及抗寒性差异[J]. 中南林业科技大学学报, 2017(9): 93-98.CAI Y X, ZENG W, JIN X L, CAI M Y. Physiological characteristics and cold hardiness between male and female plants of [J]. Journal of Central South University of Forestry & Technology, 2017(9): 93-98. (in Chinese)
[7] SUN M Y, FU L, CHEN M L, WANG X, SONG M Y, LI Z. Characterization of two winter wheat varieties’ responses to freezing in a frigid region of China[J]. Canadian Journal of Plant Science, 2016, 97(5): 8.
[8] BEZIRGANOALU, UYSAL P, YIIT O R. Cold stress resistance and the antioxidant enzyme system in [J]. Journal of Animal and Plant Sciences, 2018, 28(2): 561-567.
[9] LV C J, SUN L J, WEI X, GAO S H. Research advances in relationship between soluble sugar and cold resistance in grape[J]. Northern Fruits, 2018, 35(4): 25-27.
[10] SUN Y, HE Y, IRFAN A R, LIU X, YANG D. Exogenous brassinolide enhances the growth and cold resistance of maize (L) seedlings under chilling stress[J]. Agronomy, 2020, 10(4): 488.
[11] 胡 濤. 低温对水稻根系生理特性及其基因表达的影响[D]. 沈阳: 沈阳农业大学, 2019.FU T. Effects of low temperature on the physiological characteristics and gene expression of rice roots[D]. Shenyang: Shenyang Agricultural University, 2019. (in Chinese)
[12] BAO Y, YANG N, MENG J, WANG D, JING C. Research on the adaptability of winter wheat Dongnongdongmai1 (L) to overwintering in alpine regions[J]. Plant Biology, 2020, 23(3): 11-13.
[13] DOGRU A, HÜSNÜ AKRLAR. Is leaf age a predictor for cold tolerance in winter oilseed rape plants[J]. Functional Plant Biology, 2020, 47(3): 13-15.
[14] 中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京: 科学出版社, 1999.Editorial Committee of Chinese Flora of Chinese Academy of Sciences. Flora of China[M]. Beijing: Science Press, 1999. (in Chinese)
[15] 陈 媚, 刘迪发, 徐 丽, 姚碧娇, 李祥恩, 符小琴, 高 玲. 西番莲研究进展[J]. 中国南方果树, 2020, 49(6): 182-190.CHEN M, LIU D F, XU L, YAO B J, LI X E, FU X Q, GAO L. Research progress of [J]. South China Fruits, 2020, 49(6): 182-190. (in Chinese)
[16] 吕文强, 唐金刚, 罗时琴, 林 涛, 周传艳. 4种植被恢复模式对贵州石漠化地区表层土壤有机碳氮的影响研究[J]. 林业资源管理, 2016(5): 47-52.LYU W Q, TANG J G, LUO S Q, LIN T, ZHOU C Y. Effect of 4 kinds of vegetation restoration patterns on the topsoil organic carbon and nitrogen in rocky desertification region of Guizhou province[J]. Forestry Resource Management, 2016(5): 47-52. (in Chinese)
[17] 董万鹏, 龙秀琴, 代丽华, 孙立坤, 李培林, 杨雨嘉. 越冬期间西番莲低温半致死温度及越冬表现研究[J]. 核农学报, 2016(8): 1656-1663.DONG W P, LONG X Q, DAI L H, SUN L K, LI P L, YANG Y J. Study on the semi-lethal temperature and wintering performance of in overwintering[J]. Journal of Nuclear Agricultural Sciences, 2016(8): 1656-1663. (in Chinese)
[18] 李合生. 植物生理生化實验原理和技术[M]. 北京: 高等教育出版社, 2000.LI H S. Principles and techniques of plant physiology and biochemistry experiments[M]. Bejing: Higher Education Press, 2000. (in Chinese)
[19] 董万鹏, 罗 充, 龙秀琴, 胡 静, 李 燕. 低温胁迫对西番莲抗寒生理指标的影响[J]. 植物生理学报, 2015(5): 771-777.DONG W P, LUO C, LONG X Q, HU J, LI Y. Effects of low temperature stress on physiological indexes of cold resistance of [J]. Plant Physiology Journal, 2015(5): 771-777. (in Chinese)
[20] 姜丽娜, 张黛静, 宋 飞, 刘 佩, 樊婷婷, 余海波. 不同品种小麦叶片对拔节期低温的生理响应及抗寒性评价[J]. 生态学报, 2014, 34(15): 4251-4261.JANG L N, ZHANG D J, SONG F, LIU P, FAN T T, YU H B. Evaluation of cold resistance of different wheat varieties based on physiological responses of leaves to low temperature at the jointing stage[J]. Acta Ecologica Sinica, 2014, 34 (15): 4251-4261. (in Chinese)
[21] 何跃君, 薛 立, 任向荣, 曹 鹤, 梁丽丽, 徐 燕. 低温胁迫对六种苗木生理特性的影响[J]. 生态学杂志, 2008, 27(4): 524-531.HE Y J, XUE L, REN X R, CAO H, LIANG L L, XU Y. Effects of low temperature stress on physiological characteristics of six trees peciesseed-lings[J]. Chinese Journal of Ecology, 2008, 27(4): 524-531. (in Chinese)
[22] GASPAR T, FRANCK T, BISBIS B, KEVERC C, JOUVER L, HAUSMAN J F, DOMMED J. Concepts in plant stress physiolog application to plant tissue cultures[J]. Plant Growth Regulation, 2002, 37(3): 263-285.
[23] SONG G L, HOU W H, WANG Q H, WANG J L, JIN X C. Effect of low temperature on eutrophicated waterbody restoration by [J]. Bioresource Technology, 2006, 97(15): 1865-1869.
[24] 高 媛, 齐晓花, 杨景华, 张明方. 高等植物对低温胁迫的响应研究[J]. 北方园艺, 2007(10): 58-61.GAO Y, QI X H, YANG J H, ZHANG M F. The response mechanism of cold stress in higher plants[J]. Northern Horticulture, 2007(10): 58-61. (in Chinese)
[25] HUSSAIN S, LIU G, LIU D, AHMED M, HUSSAIN N, TENG Y. Study on the expression of dehydrin genes and activities of antioxidative enzymes in floral buds of two sand pear ( Nakai) cultivars requiring different chilling hours for bud break[J]. Journal of Interventional Radiology, 2015, 39(6): 10-11.
