油菜素内酯对稻梨孢菌的抑菌活性
2018-01-27王梦姣
王梦姣
(陕西理工大学 生物科学与工程学院/陕西省资源生物重点实验室/陕西省食药用菌工程技术研究中心,陕西 汉中 723000)
稻瘟病是一种真菌性水稻(Oryzasativa)病害[1],我国南、北方稻区均有可能发生,一般造成减产10%~20%,严重时达到40%~50%,有的地方甚至颗粒无收[2-4]。病原菌为稻梨孢菌(Pyriculariaoryzae),其还能感染其他重要农作物,比如感染大麦(Hordeumvulgare)和小麦(Triticumaestivum)后导致其严重减产[5]。其传播途径具有循环性,并且病原真菌具有不易完全杀灭的特性,因此,水稻等作物的整个生育期都存在被感染的可能性[5]。目前主要依赖的防治措施,如种植抗性品种、减少菌源、药剂防治等[6],均存在一定局限性和不足。尤其是施用的药剂复杂,成分多源自化学合成,一般要进行多次药剂处理,且药剂作用效果不显著,极易造成稻田的二次污染[7-8]。因此,研究如何有效抑制稻瘟病菌的传播、降低其对农作物尤其是对水稻的侵害至关重要。
油菜素内酯(brassinosteroid,BR)为植物重要的生长调节剂之一,是植物生长发育必需的甾醇类激素[9-12]。研究表明,油菜素内酯不仅能够参与植物营养生长和生殖生长,还能够增强植物抵抗病虫害的能力[13]。Kutschera等[14]认为,油菜素内酯作为激活剂,能够激活植物内源杀菌剂,进而帮助植物抵抗外界病原微生物。但他并没有说明,油菜素内酯激活的是哪种植物内源杀菌剂,该杀菌剂如何在植物体内起作用。Belkhadir等[15]在试验中发现,油菜素内酯通过微生物介导的分子模式调节植物免疫应答的方法,增强植物对病虫害的抵抗能力。通过外源油菜素内酯处理,Nakashita等[16]发现油菜素内酯通过诱导抗性基因这一途径,达到提高植物抵抗病虫害能力的目的。在采用遗传学方法改变了黄瓜体内的油菜素内酯含量后,Xia等[17]发现,烟酰胺腺嘌呤二核苷酸磷酸(NADPH)氧化酶活性的增强会使黄瓜体内过氧化氢含量升高,进而帮助油菜素内酯提高植株对病虫害的抵抗能力。
目前,国内关于油菜素内酯对稻梨孢菌抑制作用的研究尚处空白阶段,为此,采用油菜素内酯及与其具有类似或相反功能的其他4种植物生长调节剂处理稻梨孢菌,探索其对稻梨孢菌的抑制作用,以期为采用植物源性药剂防治稻瘟病提供科学依据及研究思路。
1 材料和方法
1.1 稻梨孢菌、植物生长调节剂的来源
稻梨孢菌购自中国农业微生物菌种保藏管理中心(ACCC),编号分别为37631、37661。
选用油菜素内酯及其他4种与油菜素内酯功能相似或相反的植物生长调节剂对稻梨孢菌进行处理,各生长调节剂相关信息见表1。

表1 5种植物生长调节剂相关信息
1.2 植物生长调节剂对菌丝生长抑制活性测定
将固体CPDA培养基(配方为:每1 L培养基中加200 g土豆、5 g蛋白胨、20 g葡萄糖、3.0 g MgSO4、5.0 g KH2PO4、维生素B1和B2各1片、琼脂8~15 g)高温高压灭菌,待温度降至55 ℃后,加入不同浓度的植物生长调节剂(表2),混匀、倒平板。待平板表面完全干透后,将培养好的稻梨孢菌从原始平板上切下0.5 cm×0.5 cm方形菌丝块,接种至新的培养基中部。每种菌每个生长调节剂浓度处理做5个重复。28 ℃恒温培养,每日测量菌体直径。待处理浓度为0 μmol/L(对照)的植物生长调节剂平板上菌落长至满皿时,计算菌丝延伸抑制率[18],并对各处理菌落形态进行观察、拍照。菌丝延伸抑制率计算公式为:菌丝延伸抑制率=(对照菌丝延伸长度-处理菌丝延伸长度)/对照菌丝延伸长度×100%。
采用回归分析法对5 种植物生长调节剂各处理浓度的对数值(x) 和菌丝延伸抑制率的概率值(y) 进行分析,计算出5 种植物生长调节剂对2种供试菌株的毒力回归方程、相关系数(r)、半数效应浓度(EC50)。

表2 5种植物生长调节剂处理浓度
1.3 油菜素内酯对菌丝体干质量的影响试验
将液体CPDA培养基分装至三角瓶中,高温高压灭菌,待温度降至55 ℃后,加入0(对照)、0.5、2 μmol/L epi-BL,混匀,接种0.5 cm×0.5 cm方形菌丝块,200 r/min、28 ℃摇菌培养。培养第4天(经过前期试验,第4天时epi-BL对菌丝延伸抑制现象开始出现;同时,到第4天时菌丝生长状态良好,烘干称质量时可以忽略初始接种菌块对其质量的影响)开始,每天取出三角瓶,将菌丝抽滤、洗涤、离心、烘干,称恒质量。至菌丝长满三角瓶(第8天)时停止试验。每个处理重复3次。菌丝干质量变化率计算公式为:M=(Mc-M^)/Mc×100%。M:菌丝干质量变化率;Mc:对照菌丝干质量;M^:试验组菌丝干质量。
1.4 油菜素内酯对孢子萌发的抑制试验
将0.5 cm×0.5 cm方形菌丝块分多点接种到米糠培养基(配方为:选无霉稻草切成3 cm小段,用清水浸泡12 h,充分吸水后倒掉水,加米糠和水,水量以手握时指间有水珠为宜;干稻草最终比例为75%,米糠为25%)上,置于恒温培养箱(28 ℃)中培养。待菌丝长满,将少许无菌水倒入培养皿中,用毛笔刷下菌丝后,置于365 nm 黑光灯下培养,48 h后,培养皿中即产生大量孢子。刷洗霉层,过滤、镜检,调配成孢子悬浮液,向其中加入不同浓度的epi-BL(表2),每个浓度处理设3个重复。将孢子悬浮液振荡培养24 h 后,镜检孢子萌发情况,计算孢子萌发抑制率,孢子萌发抑制率= (对照萌发率-处理萌发率) /对照萌发率×100%[19]。利用回归分析方法,计算回归方程、r和EC50值。
2 结果与分析
2.1 5种植物生长调节剂对稻梨孢菌菌丝体延伸的抑制作用
由表3可知,epi-BL对稻梨孢菌菌丝延伸有较强的抑制作用,随着处理浓度的升高,稻梨孢菌菌落直径递减、抑制率升高,抑制率与生长调节剂浓度呈正相关。当epi-BL浓度达到5 μmol/L时,其对37631和37661两种供试菌株菌丝延伸的抑制率均达到50%以上(分别为58.18%和53.37%)。图1中A和B分别是37631和37661两种供试菌在不同浓度epi-BL处理后平皿上的菌落形态图。从图1可以直观地观察到,epi-BL对稻梨孢菌菌丝延伸有较强的抑制作用。同时还能发现,与对照平皿相比,在不同浓度生长调节剂处理情况下,菌落颜色、表面质地、边缘形状均发生改变。37631内部菌落颜色随epi-BL浓度升高而逐渐加深,菌落边缘有明显白圈,菌落半径逐渐缩小,至5 μmol/L处理时菌丝几乎不延伸,只能观察到菌落边缘白圈;37661菌落颜色随epi-BL浓度升高也逐渐加深,菌落半径逐渐缩小,至5 μmol/L处理时菌落表面质地已变为水状。这说明,epi-BL除了能够强烈抑制菌丝延伸,还能改变菌落形态。

表3 5种植物生长调节剂对稻梨孢菌菌丝延伸的抑制率 %

图1 不同浓度油菜素内酯处理下稻梨孢菌株37631(A)和37661(B)的菌落生长情况
从表3还可以看出,不同浓度的IAA和ABA对37631的菌丝延伸具有抑制作用,但抑制作用较小,抑制率均在5%以下;6-BA和GA对37631的菌丝延伸具有促进作用,菌丝抑制率为负数。IAA和GA对37661菌丝延伸具有抑制作用,但抑制作用不明显;6-BA对37661具有一定促进作用,100 μmol/L ABA对37661的菌丝延伸具有一定抑制作用,抑制率为19.70%。
对以上5种植物生长调节剂处理结果进行直线回归分析,结果见表4。由表4可以看出, epi-BL、IAA和ABA处理浓度与菌丝延伸抑制率呈正相关,GA和6-BA呈负相关。在正相关的3种生长调节剂中,epi-BL对37631和37661两株供试菌的EC50值较小,其毒力回归方程分别为y=4.432+0.993x和y=3.868+1.576x,EC50值分别为3.7 μmol/L和5.2 μmol/L。
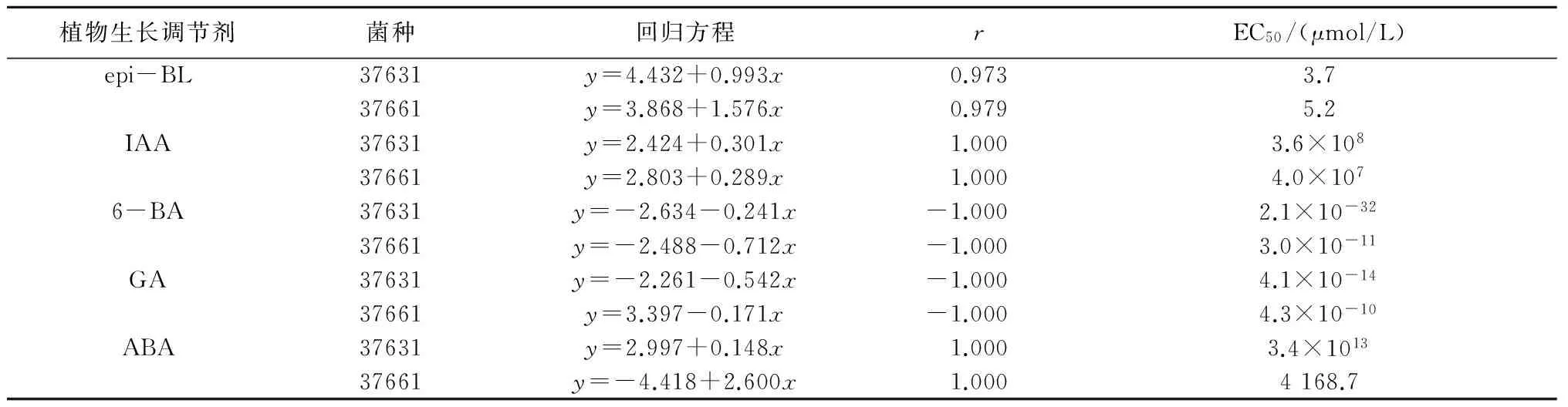
表4 5种植物生长调节剂对稻梨孢菌菌丝延伸的毒力回归分析
2.2 油菜素内酯对稻梨孢菌菌丝体生长量的影响
通过图2可以看出,油菜素内酯能够影响稻梨孢菌菌丝体的正常生长。随着培养时间的延长和处理浓度的增大,epi-BL对菌丝干质量的影响程度逐渐增大。培养至第8天时,与对照组相比,在0.5 μmol/L处理情况下,稻梨孢37631和37661菌丝干质量分别减少了29.86%和27.18%;在2 μmol/L处理情况下,稻梨孢37631和37661菌丝干质量分别减少了52.60%和48.44%。
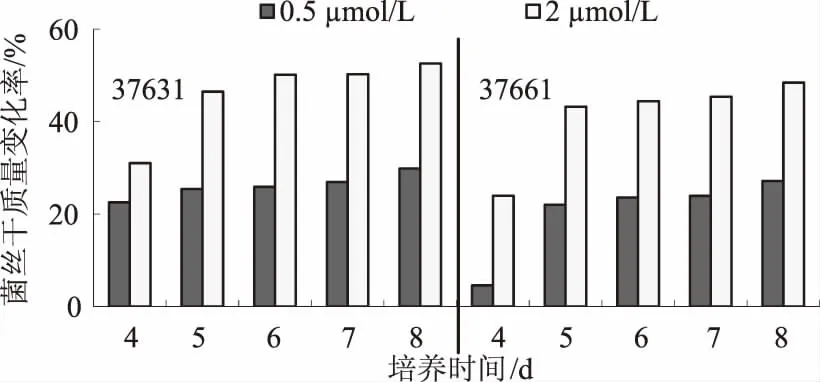
图2 不同浓度油菜素内酯处理下稻梨孢菌菌丝体干质量的变化
2.3 油菜素内酯对稻梨孢菌孢子萌发的抑制作用
从表5 可以看出,epi-BL对稻梨孢菌孢子萌发具有抑制作用,随着浓度升高,抑制率逐渐上升。当epi-BL浓度为2 μmol/L时,其对37631和37661两个供试菌株的孢子萌发抑制率分别为36.89%和23.98%;当浓度为5 μmol/L 时,抑制率分别达到53.25%和49.34%。这与epi-BL对稻梨孢菌菌丝延伸抑制的分析结果一致。
对上述结果进行回归分析发现,epi-BL的浓度对数与孢子萌发抑制率概率值均呈线性关系(表6)。epi-BL对37631和37661两株菌的毒力回归方程分别为y=4.446+0.873x和y=4.135+1.071x,EC50值分别为4.31 μmol/L和6.42 μmol/L。

表5 油菜素内酯对稻梨孢菌孢子萌发的抑制率 %
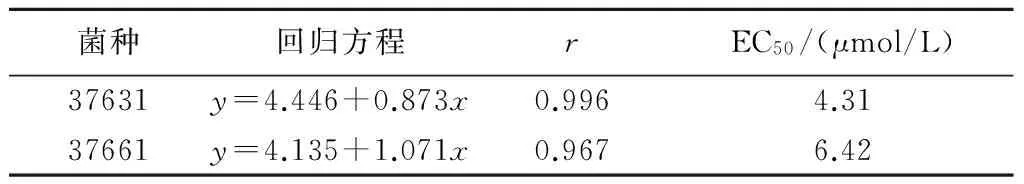
表6 油菜素内酯对稻梨孢菌孢子萌发的毒力回归分析
3 结论与讨论
某些植物生长调节剂会直接参与植物抵御外界病原微生物侵染的过程[20-21],本试验以2株稻瘟病菌为供试菌株,探究了5种植物生长调节剂对稻瘟病菌生长的抑制作用。结果显示,油菜素内酯对稻瘟病菌菌丝延伸具有明显的抑制作用,当浓度达到5 μmol/L时,其对37631和37661两个菌株的菌丝延伸抑制率分别为58.18%和53.37%,EC50值分别为3.7 μmol/L和5.2 μmol/L。其他4种植物生长调节剂对稻瘟病菌菌丝延伸没有明显的抑制作用。菌丝干质量测定结果显示,油菜素内酯能明显抑制稻瘟病菌的生长,在2 μmol/L epi-BL处理情况下,培养至第8天时37631和37661两株稻瘟病菌菌丝干质量与对照相比分别减少了52.60%和48.44%。此外,油菜素内酯对稻瘟病菌孢子萌发也具有明显的抑制作用,当浓度达到5 μmol/L时,其对37631和37661两个菌株的孢子萌发抑制率分别为53.25%和49.34%,EC50值分别为4.31 μmol/L和6.42 μmol/L。
虽然部分植物激素在促进植物生长发育这一功能上具有一定共性,但其在参与植物抵抗病原微生物方面则具有较大差异。例如,油菜素内酯和生长素均具有促进植物生长的作用,但在细条病菌和稻梨孢菌侵染水稻过程中,生长素起毒力因子的作用[22],然而,根据本试验结果,油菜素内酯能够直接抑制稻梨孢菌生长。赤霉素和油菜素内酯都可以促进植物叶和芽的生长,但李双胜[23]的研究表明,赤霉素在提高植物抵抗核盘菌的过程中起正作用因子的作用,但油菜素内酯为负作用因子。本试验结果表明,油菜素内酯对稻梨孢菌具有显著的抑制作用,为后续利用油菜素内酯协助水稻抵抗稻瘟病菌提供了可靠的试验基础。
同时,值得注意的是,稻梨孢不仅菌丝延伸速度受到了油菜素内酯的强烈影响,其菌落形态也产生了一定改变, 37661菌株在5 μmol/L epi-BL处理条件下,菌落形态改变最为明显。这说明,油菜素内酯不仅参与了稻梨孢菌丝体的分裂,还可能涉及其生理生化代谢。其他植物生长调节剂虽然没有明显影响稻梨孢菌丝体的延伸速度,但是处理后2个菌株的菌落表面形态与对照相比都发生了一定变化(未列出),这说明,其他植物生长调节剂可能也参与了菌体的生长代谢。
[1] Law J W,Ser H L,Khan T M,etal.The potential ofStreptomycesas biocontrol agents against the rice blast fungus,Magnaportheoryzae(Pyriculariaoryzae)[J].Front Microbiol,2017,8(3):45-66.
[2] Zhong X H,Yang J X,Shi Y L,etal.The DnaJ protein OsDjA6 negatively regulates rice innate immunity to the blast fungusMagnaportheoryzae[J].Mol Plant Pathol,2017,2:12546-12561.
[3] Liu H,Dong S,Gu F,etal.NBS-LRR protein Pik-H4 interacts with OsBIHD1 to balance rice blast resistance and growth by coordinating ethylene-brassinosteroid pathway[J].Front Plant Sci,2017,8:127-135.
[4] Schauer S,Kutschera U.A novel growth-promoting microbe,Methylobacteriumfunariaesp.nov.,isolated from the leaf surface of a common moss[J].Plant Signaling and Behavior,2011,6:510-515.
[5] Liao J H,Chen P Y,Yang Y L,etal.Clarification of the antagonistic effect of the lipopeptides produced byBacillusamyloliquefaciensBPD1 againstPyriculariaoryzaevia in situ MALDI-TOF IMS analysis[J].Molecules,2016,21(12):1670-1692.
[6] 温小红,谢明杰,姜健,等.水稻稻瘟病防治方法研究进展[J].中国农学通报,2013,29(3):190-195.
[7] Wang X,Chen Y F,Yan W,etal.Synthesis and biological evaluation of benzimidazole phenylhydrazone derivatives as antifungal agents against phytopathogenic fungi[J].Molecules,2016,21(11):1574-1589.
[8] Wang X,Dai Z C,Chen Y F,etal.Synthesis of 1,2,3-triazole hydrazide derivatives exhibiting anti-phytopathogenic activity[J].Eur J Med Chem,2017,126:171-182.
[9] 王梦姣,邓百万,杨国鹏.拟南芥油菜素内酯的合成、修饰、信号转导及其在农作物育种中的应用研究进展[J].河南农业科学,2015,44(2):1-6.
[10] Khripach V,Zhabinskii V,de Groot A.Twenty years of brassinosteroids:Steroidal plant hormones warrant better crops for the ⅩⅪ century[J].Annals of Botany,2000,86:441-447.
[11] Wang M J,Liu X Y,Wang R,etal.Overexpression of a putativeArabidopsisBAHD acyltransferase causes dwarfism that can be rescued by brassinosteroid[J].Journal of Experimental Botany,2012,63(16):5787-5801.
[12] Greco M,Chiappetta A,Bruno L,etal.InPosidoniaoceanicacadmium induces changes in DNA methylation and chromatin patterning[J].J Exp Bot,2012,63(2):695-709.
[13] Deb A,Grewal R K,Kundu S.Regulatory cross-talks and cascades in rice hormone biosynthesis pathways contribute to stress signaling[J].Front Plant Sci,2016,7:1303-1312.
[14] Kutschera U,Wang Z Y.Brassinosteroid action in flowering plants:A Darwinian perspective[J].Journal of Experimental Botany,2012,63(10):3511-3522.
[15] Belkhadir Y,Jaillais Y,Epple P,etal.Brassinosteroids modulate the efficiency of plant immune responses to microbe-associated molecular patterns[J].PNAS,2012,109(1):297-302.
[16] Nakashita H,Yasuda M,Nitta T,etal.Brassinosteroid functions in a broad range of disease resistance in tobacco and rice[J].The Plant Journal,2003,33:887-898.
[17] Xia X J,Wang Y J,Zhou Y H,etal.Elevated H2O2levels resulting from enhanced NADPH oxidase activity are involved in the BR-induced stress tolerance in cucumber[J].Plant Physiology,2009,150:801-814.
[18] 檀根甲,祝建平.杀菌剂生物测定计算方法及应用[J].安徽农学通报,1998,4(1):27-28.
[19] 张翠荣,李明,李荣玉.5种植物粗提物对稻瘟病菌的抑菌活性[J].江苏农业科学,2016,44(8):162-164.
[20] 张智慧,聂燕芳,何磊,等.外源茉莉酸甲酯诱导水稻抗瘟性相关防御酶和内源水杨酸的变化[J].植物病理学报,2010,40(4):395-403.
[21] 向妙莲,付永琦,何永明,等.茉莉酸甲酯浸种对水稻幼苗白叶枯病抗性及抗氧化酶活性的影响[J].中国水稻科学,2014,28(4):419-426.
[22] 傅晶.抑制病原菌诱导的生长素的积累赋予水稻广谱抗性[D].武汉:华中农业大学,2010.
[23] 李双胜.草酸在核盘菌(Sclerotiniasclerotiorum)与寄主植物互作中的生物学作用[D].杭州:浙江大学,2013.
