2013~2015年黑龙江省马铃薯晚疫病菌群体结构研究
2018-01-25吕文河吕典秋闵凡祥白雅梅王晓丹
吕文河,敖 翔,吕典秋,闵凡祥,白雅梅,王晓丹
(1.东北农业大学农学院,哈尔滨 150030;2.黑龙江省农业科学院植物脱毒苗木研究所,哈尔滨 150086;3.东北农业大学资源与环境学院,哈尔滨 150030)
马铃薯是我国重要粮食作物和经济作物之一,具有适应性强、产量高、营养丰富等特性和一定药用价值[1]。马铃薯生产过程易受病害影响,其中晚疫病尤为严重。马铃薯晚疫病(Late blight)为致病疫霉(Phytophthora infestans)引起的毁灭性病害。晚疫病可发生于马铃薯整株,严重时导致马铃薯茎叶死亡和块茎腐烂。一般可造成马铃薯减产20%~30%,甚至50%以上至绝收。
致病疫霉属于异宗配合、半活体营养型卵菌,存在A1、A2两种交配型。两种交配型同时存在,可通过有性生殖发生基因重组,产生卵孢子。Niederhauser首次发现大量卵孢子,证实晚疫病菌存在A2交配型[2]。20世纪80年代后,国内外学者相继报道A2交配型,“新”群体扩散,晚疫病菌群体结构组成逐渐复杂[3]。张志铭等测定国内多省菌株,在内蒙古和山西菌株中发现3株A2交配型[4]。随后其他省市发现A2交配型[5-7]。朱杰华测定2000年采集黑龙江省不同地点45株晚疫病菌株,仅克山发现1株A2交配型菌株[8]。但金光辉等后续报道中未发现A2交配型[9-11],近年王腾等再次证实黑龙江省内存在A2交配型菌株[12-14]。生理小种为晚疫病菌重要表现型之一,其组成与变异直接关系马铃薯晚疫病发生与流行[15]。国内各地晚疫病菌生理小种组成逐渐复杂,发现“超级毒力小种”[11,16-17]。目前,马铃薯晚疫病防治手段主要为化学防治,其中苯基酰胺类杀菌剂甲霜灵(metalaxyl)对晚疫病防效较好,具有保护兼治疗作用,但极易产生抗药性。国内外已相继发现抗性菌株[7,18]。朱杰华等测定晚疫病菌对甲霜灵敏感性,发现黑龙江省内大多数马铃薯主产区均已存在抗性菌株,并占据主导地位[8,11-12,19]。A2交配型影响黑龙江省晚疫病菌群体结构,不同年份、地点甲霜灵类杀菌剂使用存在差异,因此对晚疫病菌株群体结构开展持续性研究十分必要。
本研究通过测定2013~2015年黑龙江省马铃薯晚疫病菌株交配型、甲霜灵敏感性并作生理小种鉴定,探究黑龙江省晚疫病菌群体遗传结构变化,以期为晚疫病防治提供科学依据。
1 材料与方法
1.1 材料
试验待测菌株为2013~2015年采集自黑龙江省齐齐哈尔、绥化、哈尔滨和佳木斯,经分离、纯化后保存(菌株由黑龙江省农业科学院植物脱毒苗木研究所提供)。
交配型A1、A2标准菌株均由比利时瓦隆农业研究中心提供。
98%甲霜灵原药购自先正达公司,用二甲基亚砜(DMSO)将原药配制成100 mg·mL-1浓度母液备用。
用于鉴定生理小种寄主共12份(r,R1-R11),均为瓶装组培苗(由国际马铃薯中心提供)。
1.2 试验方法
1.2.1 黑麦培养基制备
称取60 g黑麦(1 L培养基)于量杯中,加入蒸馏水浸泡24 h,将上清液倒出,保留备用。用破碎机将浸泡后黑麦打碎,加入适量蒸馏水于100℃水浴1 h,水量不超过1 L。水浴后将上清液与之混合,再用4层纱布过滤、弃去残渣。滤液中加琼脂15 g和蔗糖15 g,定容至1 L,搅拌均匀后加热至琼脂、蔗糖融化,然后分装至500 mL锥形瓶中,封口后放入121℃高压湿热灭菌锅中25 min,出锅后摇匀倒入平皿以备用。
1.2.2 晚疫病菌分离与纯化
选取易感病品种马铃薯块茎表面消毒,切成V字型薯片(5 mm厚),斜面切一深槽,将马铃薯单病斑叶片夹入深槽,置于18℃光周期培养箱高湿黑暗培养5~7 d。待长出菌丝后,挑取少量菌丝于黑麦培养基上18℃光周期培养箱高湿黑暗培养,7 d后在菌落边缘切取有单菌丝培养基置于黑麦培养基上纯化培养。菌落形成后即转移至黑麦斜面培养基保存备用[20]。
1.2.3 交配型测定
采用皿内对峙法测定交配型[9]:在黑麦培养基上,将直径为6 mm待测菌株菌饼分别与同等尺寸A1、A2标准菌株菌饼对峙培养,两菌饼间距1.5~2.0 cm。将接菌培养皿倒置于18℃暗培养箱中培养直至两菌落相交。显微镜观察两菌落相交处卵孢子产生情况和待测菌株单独培养时是否有卵孢子产生。如果待测菌株仅与A2对峙培养时产生卵孢子,则该菌株为A1交配型;反之,仅与A1对峙培养时产生卵孢子则为A2交配型;如果待测菌株单独培养时即产生卵孢子,则为自育型菌株。
1.2.4 甲霜灵敏感性测定
采用菌落直径法测定甲霜灵敏感性[21]:取直径为8 mm待测菌株菌饼接种到含甲霜灵0、5、100 μg·mL-1黑麦培养基,每个处理3次重复。置于18℃光周期培养箱高湿黑暗培养7 d(对照菌落直径大于50 mm),十字交叉法量取菌落直径。甲霜灵敏感性划分标准[22]如下:敏感菌株(S):含甲霜灵5、100 μg·mL-1平板菌落直径≤40%对照菌落直径;中抗菌株(MR):含甲霜灵5 μg·mL-1平板菌落直径≥40%对照菌落直径且100 μg·mL-1平板菌落直径≤40%对照菌落直径;高抗菌株(R):含甲霜灵 5、100 μg·mL-1平板菌落直径≥40%对照菌落直径。
1.2.5 生理小种鉴定
将待测菌株预培养10 d,在培养皿中加入适量灭菌水,用涂布棒将孢子囊充分洗出,过滤吸出液体即为孢子囊悬浮液,4℃环境下2~3 h待其释放游动孢子即为游动孢子悬浮液,利用血球计数板将浓度调至5×104个·mL-1备用。每个鉴别寄主3瓶苗(r=3),每瓶5株组培苗。采用活体接种[23],取游动孢子悬浮液10 μL,分别接种在瓶内鉴别寄主叶片上,每个鉴别寄主接种30片叶。接种后放置在20℃光周期培养箱高湿黑暗培养12 h,随后移至20℃光周期培养箱(16 h光照,8 h黑暗)。培养5 d后观察鉴别寄主是否发病,如鉴别寄主接种部位有菌丝和孢子囊产生,即确定为侵染;如无症状或出现免疫性坏死,即确定为不侵染。统计每个鉴别寄主发病情况,根据Black等[24]毒性基因与鉴别寄主基因型关系表确定待测菌株生理小种名称,并分析生理小种复杂性(Gleason(HG)多样性指数,Shannon(HS)多样性指数[25])和毒力基因复杂性(Ci和Cp指数[26])。
HG=(Np-1)/lnN;HS=-∑iPiln(Pi)。
Np:分离鉴定出生理小种个数,N:菌株数;Pi:第i个生理小种频率。
Ci=∑j(PjVj),j=1……Np;CP=(∑jVj)/Np。
Ci:每个菌株毒力基因平均值,Cp:每个小种毒力基因平均值,Pj:第j个生理小种频率,Vj:第j个生理小种毒力基因数目,Np:分离鉴定出生理小种个数。
2 结果与分析
2.1 交配型测定
测定待测菌株交配型,结果见表1。在132株晚疫病菌株中,A1交配型90株,占供试菌株68.18%;A2交配型42株,占供试菌株31.82%(见图1 A),未发现自育型菌株。由年际A2交配型分布情况看,A2交配型出现频率呈先升后降趋势,2014年达到最高(57.14%),超过A1交配型占比(见图1 B)。由地域交配型情况看,绥化地区未发现A2交配型;哈尔滨和齐齐哈尔地区检测 到A1、A2两种交配型分别占总菌株65.28%、57.50%,A1交配型占优势(见图1 C)。齐齐哈尔地区A2交配型出现频率波动较大,呈先升后降趋势;哈尔滨地区A2交配型频率变化明显,3年间呈下降趋势。
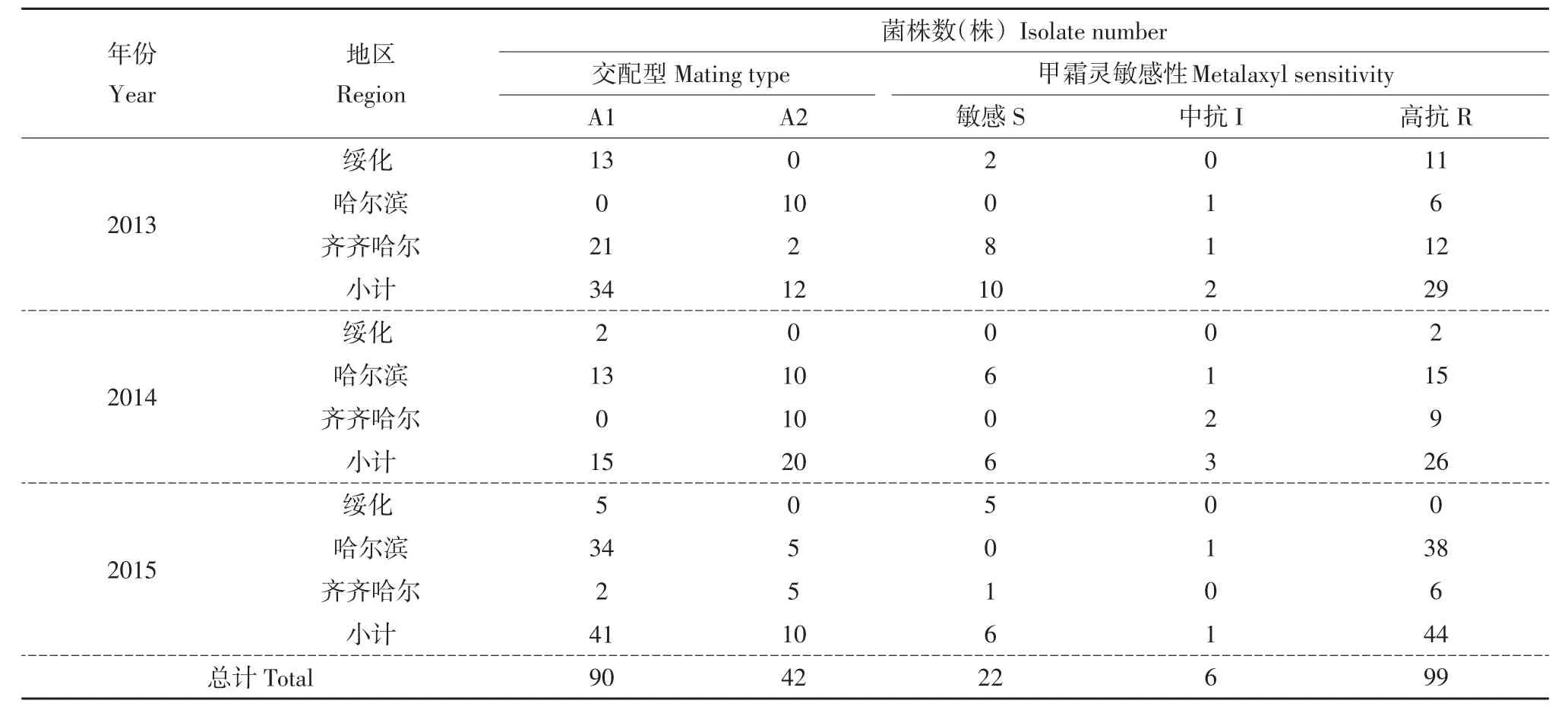
表1 2013~2015年黑龙江省马铃薯晚疫病菌交配型和甲霜灵敏感性Table1 Mating type and metalaxyl sensitivity of Phytophthora infestans isolates in Heilongjiang Province during 2013-2015
2.2 甲霜灵敏感性测定
测定待测菌株甲霜灵敏感性,结果见表1。在127株晚疫病菌株中,敏感菌株、中抗菌株、高抗菌株分别占17.32%、4.72%、77.95%(见图2 A)。在年际上,抗性菌株占比相对较高,并呈逐渐升高趋势;敏感菌株占比呈逐渐降低趋势,2015年为最低(11.76%)(见图2 B)。在地域上,绥化地区敏感菌株占比最高,达35.00%,哈尔滨地区最低,敏感菌株占总菌株8.82%(见图2 C)。
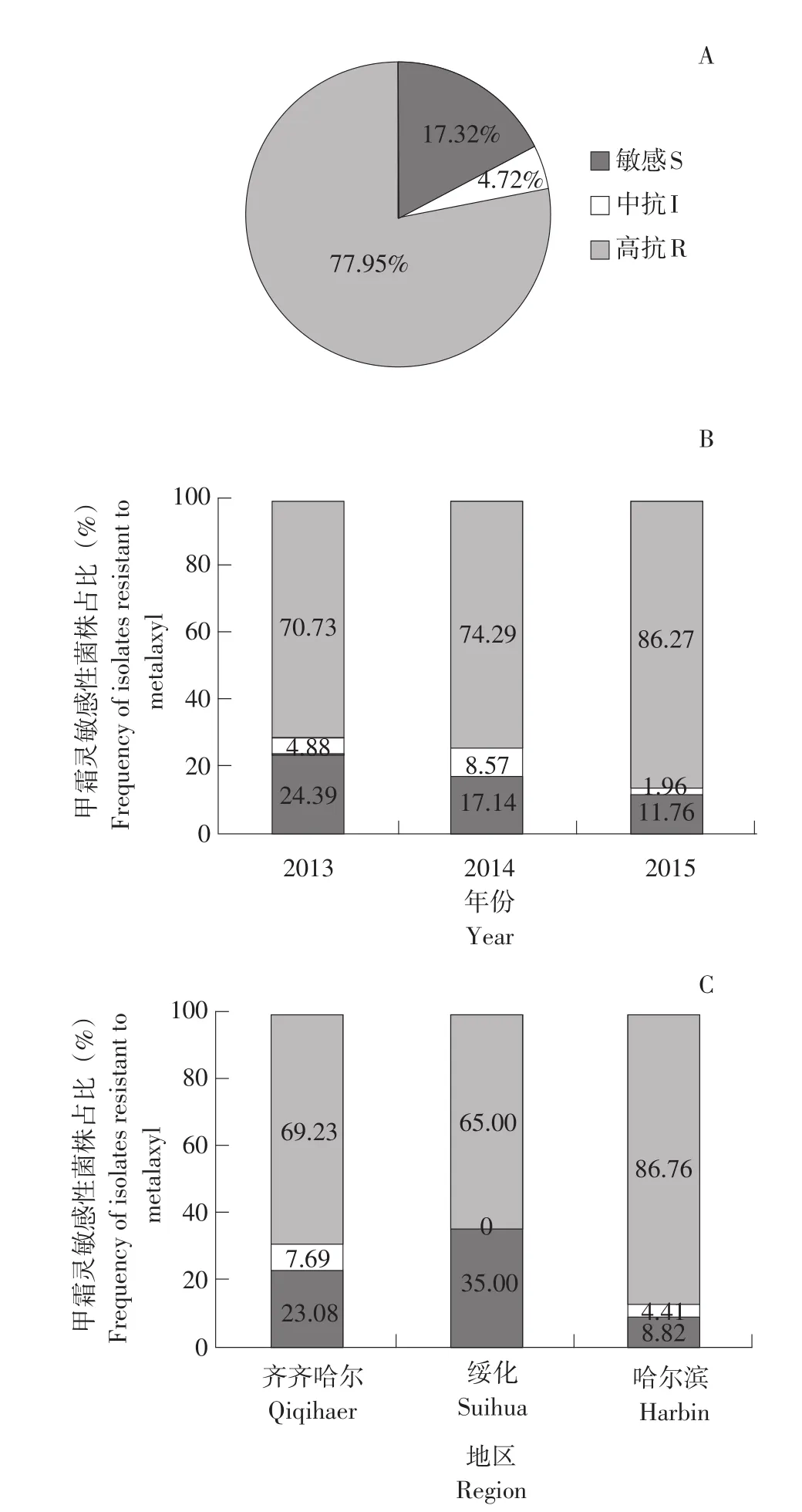
图2 2013~2015年黑龙江省晚疫病菌甲霜灵敏感性占比Fig.2 Frequency of Phytophthora infestans isolatesmetalaxyl sensitivity in Heilongjiang Province during 2013-2015
2.3 生理小种鉴定
采自2013~2015年齐齐哈尔、绥化、哈尔滨和佳木斯4个地区84株马铃薯晚疫病菌株中,共鉴定出37个生理小种类型,全部为复合毒力基因小种,优势小种为1.3.4.7.9.10(占总菌株13.10%)(见表2)。由表2可知,2013、2014和2015年优势小种分别为1.3.4.7.10.11,1.2.3.4.5.6.7.8.9.10.11和1.3.4.7.9.10,发生频率分别为11.11%;21.43%和28.95%。
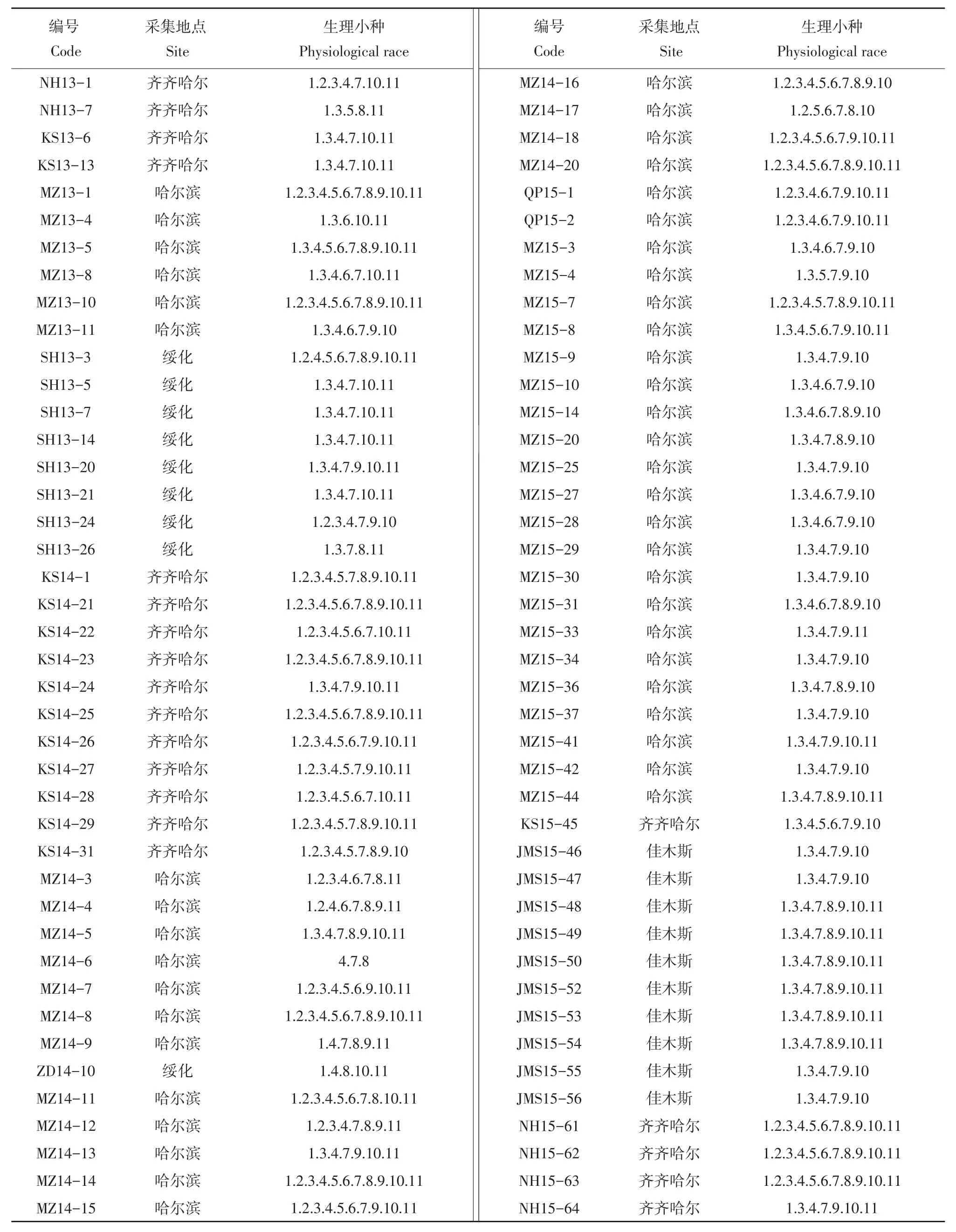
表2 2013~2015年黑龙江省晚疫病菌生理小种鉴定结果Table2 Identification of physiological races of Phytophthora infestans isolates collected from Heilongjiang Province during 2013-2015
由图3可知,毒力基因avr1、avr3、avr4、avr7、avr10各年份均表现较高频率,avr2、avr5、avr6、avr8表现频率呈先升后降趋势,avr7、avr9表现频率3年间逐渐升高,avr11则呈逐步降低趋势。由表3可知,2014年晚疫病菌株复杂程度最高(HG=5.102,HS=2.682,Ci=8.857,Cp=8.167),2013~2015年黑龙江马铃薯晚疫病菌生理小种和小种毒力基因复杂程度呈升高趋势。
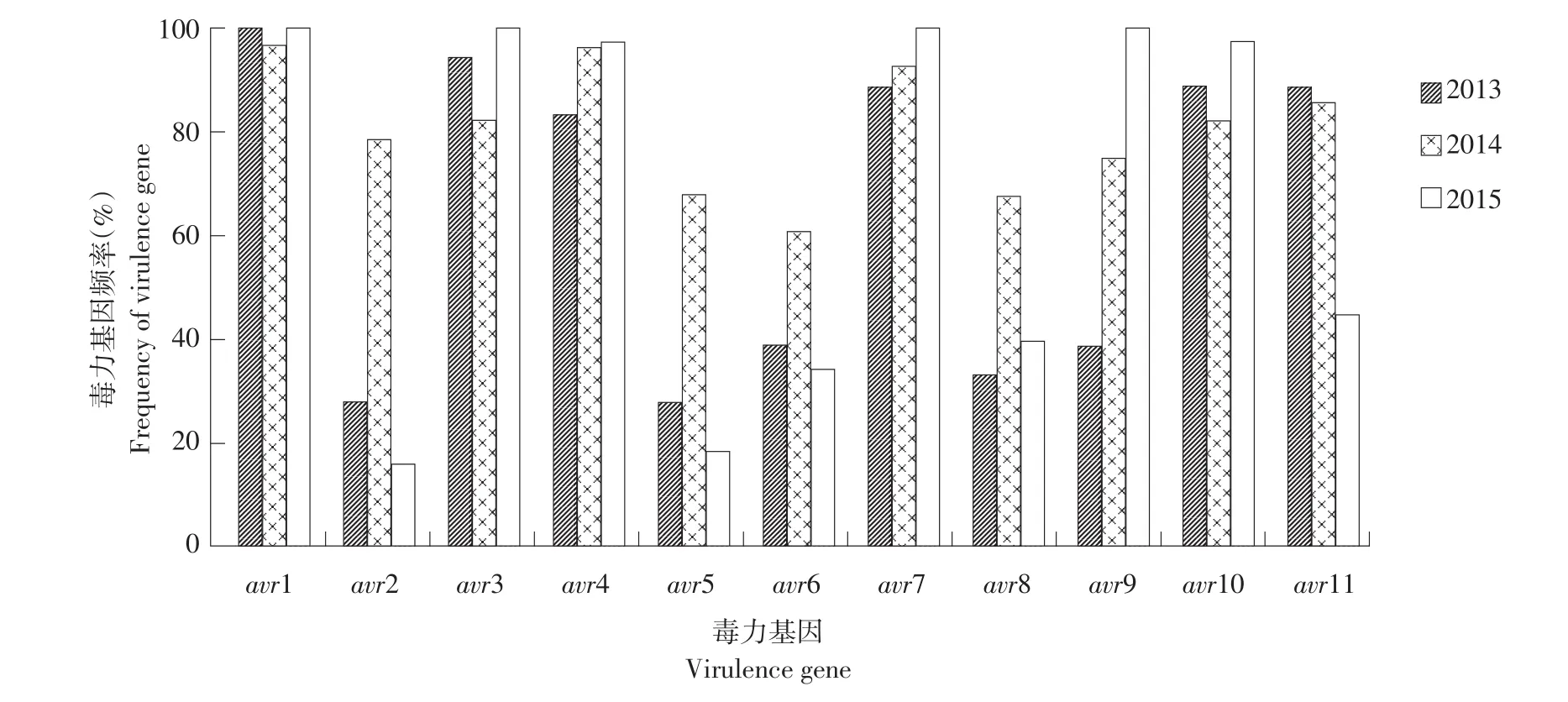
图3 2013~2015年黑龙江省晚疫病菌生理小种毒力基因出现频率Fig.3 Frequency of virulence gene of Phytophthora infestans isolates in Heilongjiang Province during 2013-2015
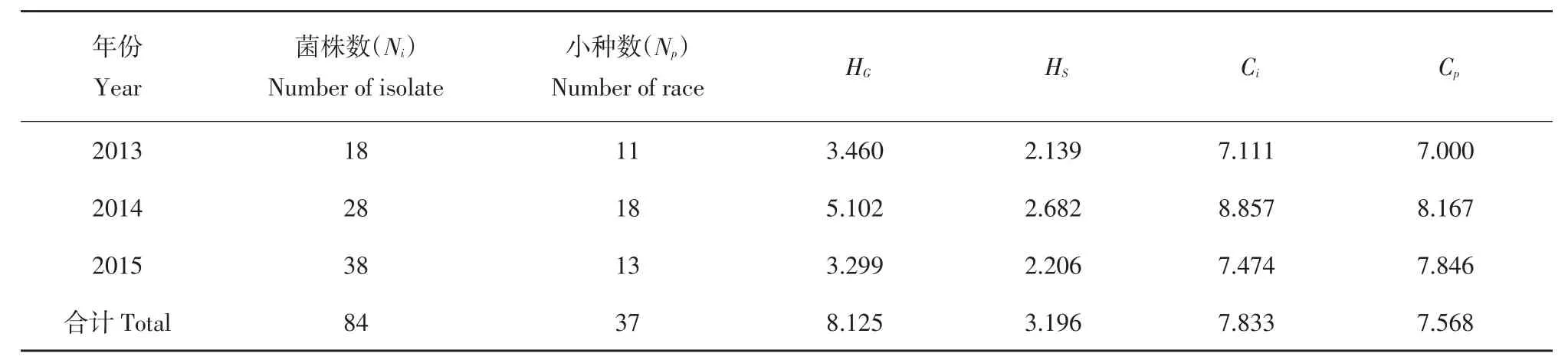
表3 2013~2015年黑龙江省马铃薯晚疫病菌生理小种及生理小种毒力基因复杂性指数Table3 Physiological races and virulence gene complexity of Phytophthora infestans isolates in Heilongjiang Province during 2013-2015
3 讨论与结论
本研究测定2013~2015年采自齐齐哈尔、绥化和哈尔滨晚疫病菌株交配型,发现A2交配型菌株42株,占总菌株31.82%,但绥化市未发现A2交配型菌株。就地域而言,近年齐齐哈尔市A2交配型菌株占比呈先升后降趋势,绥化市未发现A2交配型(2013~2015年),与王腾等[12]研究结果一致;就年际而言,3年间A2交配型菌株占比呈先升后降趋势,与张铉哲等[14]研究结果不同,具体原因尚待进一步研究。A2交配型在黑龙江省多数马铃薯主要产区均有分布,目前A2交配型总占比低于A1交配型,但部分地区A2交配型菌株已成为优势病原菌群(如齐齐哈尔市)。绥化市供试菌株中并未发现A2交配型,这种地区间病菌群体结构差异,可能因种薯贸易造成[7-8]。带病种薯加快A2交配型传播与流行,改变各地区晚疫病菌群体结构[20]。
甲霜灵高抗菌株已普遍存在,并逐渐成为优势菌群[27-29]。本研究结果表明,黑龙江省马铃薯晚疫病菌已对甲霜灵产生较强抗药性,抗性菌株频率呈上升趋势。一方面可能与A2交配型大量发现有关,Pekad等发现在112株A2交配型菌株中,高抗菌株占98%,49株A1交配型菌株中高抗菌株占69%[30];另一方面推测与连续使用甲霜灵类杀菌剂有关[7,31]。就地域而言,施药类型不同,可造成抗性菌株频率差异。金光辉测定2004~2006年黑龙江晚疫病菌甲霜灵敏感性,发现抗性菌株占总菌株74.29%[32]。王腾等测定2011~2013年黑龙江晚疫病菌株甲霜灵敏感性,测定结果显示抗性菌株占83.92%[12]。本研究2013~2015年黑龙江晚疫病菌抗性菌株占比77.95%,说明近10年黑龙江省晚疫病菌对甲霜灵抗药性总体上升。在高抗菌株优势地区应减少甲霜灵类杀菌剂使用,降低晚疫病菌选择压力;在敏感菌株优势区域,合理制定防治方案,注意轮换用药,延长药剂有效使用期限[31,33]。
就生理小种而言,中国马铃薯晚疫病生理小种类型已趋于复杂化[16-17]。王晨从2008~2011年黑龙江省41个晚疫病菌株中鉴定出15个生理小种类型,优势小种为1.2.3.4.5.6.7.9.10.11(24.39%)[34]。金光辉等测定2010年采自黑龙江省84个菌株,鉴定出生理小种类型25个,优势小种为1.3.4.7.8.10.11(27.38%)[35]。本研究在2013~2015年84株晚疫病菌中共鉴定出37个生理小种类型,优势小种为1.3.4.7.9.10(13.10%),说明近年优势小种毒力基因数目和占比显著减少,但其他生理小种类型显著增多。另外,本研究表明,2013~2015年黑龙江省马铃薯晚疫病菌生理小种复杂性和生理小种毒力基因复杂性总体升高,结合金光辉等[35]和王腾[36]研究,黑龙江省晚疫病菌生理小种复杂程度整体呈升高趋势。其中,2014年研究结果表明,“超级毒力小种”为当年优势小种,尽管此前在黑龙江省已发现“超级毒力小种”,但最高优势小种毒力基因个数未达到11个[37],可能是病原菌有性生殖使毒力基因重组,形成“超级毒力小种”,增加晚疫病抗性育种难度。
本研究在前人研究基础上继续监测2013~2015年黑龙江省晚疫病菌群体结构。目前黑龙江省大部分马铃薯主产区均发现A2交配型,甲霜灵抗性菌株占绝对优势,小种复杂性指数持续升高,晚疫病防治难度加大。在防治方面,建议尽量使用非甲霜灵类杀菌剂,防止杀菌剂单一使用使病原菌出现抗药性;在控制方面,应保证种薯质量安全,避免带病种薯迁移、传播;加强马铃薯抗晚疫病育种研究,从源头控制马铃薯晚疫病发病与流行。
[1] 吕文河,李志燕,雷雪萍,等.离体条件下PEG-6000胁迫对马铃薯组培苗影响[J].东北农业大学学报,2015,46(10):1-9.
[2] Niederhauser J S.The blight,the blighter,and the blighted[J].Transactions of the New York Academy of Sciences,1956,19:55-63.
[3] Spielman L J,Drenth A,Davidse L C,et al.A second world-wide migration and population displacement of Phytophthora infestans?[J].Plant Pathology,2010,40(3):422-430.
[4] 张志铭,王军.中国发生马铃薯晚疫病菌(Phytopthora infestans)A2交配型[J].河北农业大学学报,1996,19(4):62-65.
[5] Huang L,Chen X J,He H Y,et al.A preliminary investigation on phenotypic characteristics of Phytophthora infestans from Guizhou Province[J].Agricultural Biotechnology,2016,5(3):29-32.
[6] 马云芳,孙洁平,马丽杰,等.一个马铃薯种质资源圃致病疫霉群体的分析[J].菌物学报,2013,32(5):802-811.
[7] 李洪浩,彭化贤,席亚东,等.四川马铃薯晚疫病菌交配型、生理小种、甲霜灵敏感性及mtDNA单倍型组成分析[J].中国农业科学,2013,46(4):728-736.
[8] 朱杰华.中国马铃薯晚疫病菌群体遗传结构研究[D].保定:河北农业大学,2004.
[9] 金光辉,袁善奎,吕文河,等.黑龙江省马铃薯晚疫病菌生理小种类型及交配型研究[J].东北农业大学学报,2006,37(5):610-614.
[10] 徐生军,郭蓉,王晨,等.马铃薯晚疫病菌对3种杀菌剂的抵抗性与交配型分化[J].东北农业大学学报,2009,40(2):1-6.
[11] 王鹤,朱杰华,杨志辉,等.2009年黑龙江和吉林省马铃薯晚疫病菌表型结构研究[J].植物保护,2012,38(1):151-154.
[12] 王腾,闵凡祥,郭梅,等.黑龙江省马铃薯晚疫病菌交配型及甲霜灵敏感性测定[J].植物保护,2016,42(1):180-183.
[13] 沙海天,万安琪,赵偲,等.2013-2014年北方5省(区)致病疫霉抗药性监测及交配型分析[J].中国植保导刊,2016,36(1):63-67.
[14] 张铉哲,郝璐,李微,等.黑龙江省马铃薯晚疫病菌交配型、瑞毒霉敏感性及mtDNA单倍型分析[J].吉林农业科学,2015,40(5):58-62.
[15] Koh Y J,Goodwin S B,Dyer A T,et al.Migrations and displacements of Phytophthora infestans populations in East Asian countries[J].Phytopathology,1994,84(9):922-927.
[16] 王立,惠娜娜,李建军,等.甘肃省马铃薯主产区晚疫病菌生理小种组成与分布[J].中国蔬菜,2013(22):70-74.
[17] 杨丽娜,段国华,覃雁瑜,等.2012年云南省会泽县马铃薯晚疫病菌小种结构分析[J].热带作物学报,2016,37(1):158-163.
[18] Sedegui M,Carroll R B,Morehart A L,et al.Genetic structure of the Phytophthora infestans population in Morocco[J].Plant Disease,2007,84(2):173-176.
[19] 孙秀梅,马颜亮,白雅梅,等.黑龙江省马铃薯晚疫病菌对甲霜灵药剂的敏感性测定[J].中国马铃薯,2009,23(2):72-74.
[20] 叶广继.青海马铃薯晚疫病菌群体遗传多样性研究[D].西宁:青海大学,2008.
[21] 穆立义.植物化学保护研究法[M].北京:中国农业出版社,1994.
[22] Oyarzun P J,Pozo A,Ordnez M E,et al.Host specificity of Phy⁃tophthora infestans on tomato and potato in Ecuador[J].Phytopathology,1998,88(3):265-271.
[23] 赵青,郑峥,李颖,等.四川省马铃薯晚疫病菌群体表型和遗传变异的分析[J].菌物学报,2016,35(1):52-62.
[24] Black W,Mastenbrock C,Mills W R,et al.A proposal for an international nomenclature of races of Phytophthora infestans and of genes controlling immunity in Solanum demissum[J].Enphytica,1953,2(3):173-179.
[25] Groth J V,Roelfe A P.The concept and measurement of phenotypic diversity in Puccinia graminiis on wheat[J].Phytopathology,1987,77(10):1395-1399.
[26] Andrivon D.Races of Phytophthora infestans in France,1991-1993[J].Potato Research,1994,37(3):279-286.
[27] 何佳昱,丁明亚,杨志辉,等.2012年河北省围场县马铃薯晚疫病菌表型及基因型分析[J].江苏农业科学,2015,43(10):179-182.
[28] Harbaoui K,Hamada W,Vleeshouwers V G A A,et al.Characterization of Phytophthora infestans isolates collected from potato and tomato crops in Tunisia during 2006-2008[J].Potato Research,2013,56(1):11-29.
[29] Savazzini F,Galletti S.Phenotypic and genotypic characterization of Italian Phytophthora infestans isolates[J].Phytopathologia Mediterranea,2015,54(3):524-530.
[30] Pekad F Z,Cooke D E L,Puglisi I,et al.Characterization of Phy⁃tophthora infestans populations in northwestern Algeria during 2008-2014[J].Fungal Biology,2017,121(5):467-477.
[31] 张大为,惠娜娜,王立,等.甘肃省马铃薯致病疫霉交配型组成及其对甲霜灵的抗药性[J].西北农业学报,2014,23(6):184-188.
[32] 金光辉.黑龙江省马铃薯晚疫病菌群体结构及抗源评价[D].哈尔滨:东北农业大学,2007.
[33] 杨继峰,朱小琼,国立耘,等.内蒙古西部地区马铃薯晚疫病菌的交配型分布及对3种杀菌剂的敏感性[J].华北农学报,2011,26(5):16-20.
[34] 王晨.马铃薯晚疫病菌的表现型和SSR基因型分析[D].哈尔滨:东北农业大学,2012.
[35] 金光辉,李学湛,王玉成,等.年际间干旱对晚疫病菌生理小种复杂性的影响[J].植物保护,2017,43(4):167-173.
[36] 王腾.黑龙江省马铃薯晚疫病菌群体结构研究及块茎抗病性鉴定[D].大庆:黑龙江八一农垦大学,2015.
[37] 金光辉,王腾,吕文河.黑龙江省马铃薯晚疫病菌群体结构及抗病种质资源研究[M].北京:中国农业出版社,2016.
