在水分胁迫下乌丹蒿苗期生长和保护酶活性的变化
2018-01-15石文宏张晓明徐振朋满达
石文宏,宛 涛*,蔡 萍,张晓明,徐振朋,满达
(1.内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010019;2.锡林郭勒职业学院草原生态与畜牧兽医学院,内蒙古 锡林浩特 026000)
内蒙古水资源分布不匀、水资源匮乏、利用状况低是我区面临的主要水问题,并且草原作为我区主要植被类型及生态屏障,发展高效节水农业、提高水的利用率显得尤为重要,因此对植物进行干旱胁迫研究,了解植物水分利用率从而更好地提高草原用水率和产草量、减少草原退化沙化面积提供基础理论依据〔1-3〕。
乌丹蒿(ArtemisiawudanicaLiou et W.Wang)属菊科蒿属半灌木,其主根深、侧根发达、地下根茎分枝能力较强,具有抵御风沙、抗旱性强、耐沙埋等生物学特性,是内蒙古特有乡土固沙先锋植物〔4〕。近年来,国内对乌丹蒿进行了不少研究,包括种子库和粘液繁殖体特性、形态与物候特征、有性和无性繁殖模式及染色体核型分析等〔5-8〕。但目前乌丹蒿在苗期干旱胁迫条件下植株生长状况及抗氧化酶活性变化等方面未见报道。本研究以乌丹蒿幼苗为供试材料,采用干旱胁迫法对其进行处理,探索其苗期生长状况及抗氧化酶活性变化,通过分析植株生长状况及抗氧化系统的变化,更好地了解其抗旱机理。
1 材料与方法
1.1 试验材料及培育
乌丹蒿种子于2015年10月采自内蒙古赤峰市翁牛特旗乌丹镇东部的响水乡,2016年7月3日将选好的种子均匀播种于盆高18cm、内径15cm的苗钵共36盆后放置于日光温室中培养(培养基质为栗钙土型风沙土准确称取2kg土后播种,沙埋深度为2cm)。正常浇水(150ml)至叶龄为2片真叶后对其中12盆植株进行水分梯度处理,处理至4片真叶时期测定生长和抗氧化酶指标;随后再选取12盆叶龄为4片真叶的植株进行与上述方法一样的处理至8片真叶时期测定各项指标;最后对剩下的12盆叶龄为8片的植株进行处理至12片真叶时期测定各项相关指标。干旱胁迫设4个处理,以对照组(CK)浇150ml、轻度组(LS)浇120ml、中度组(MS)浇90ml、重度组(HS)浇60ml的方式3d浇1次水,每种含水量重复3次。最后测得的实际含水量依次为40%、25%、15%和5%。
1.2 测定方法
土壤含水量采用浙江托普仪器有限公司土壤水分测速仪测定;株高、茎粗、根长,采用卷尺和游标卡尺直接测量;根鲜重和地上部鲜重,采用直接称量法;侧根数量:每组取30个株植接进行取值;生物量测定称量整盆内植株的地上、地下生物量;根冠比=根干重/地上部干重;过氧化氢酶(CAT)采用紫外吸收法测定;过氧化物酶(POD)采用愈创木酚法测定;超氧化物歧化酶(SOD)采用氮蓝四唑法测定〔9〕。
1.3 数据处理
所有试验数据采用Excel进行数据整理和绘制图表,采用SPSS19.0对数据进行方差分析。
2 结果与分析
2.1 水分胁迫下乌丹蒿植株生长的变化
土壤水分亏缺条件下乌丹蒿3个时期生长发育受到抑制,其株高、茎粗、根长、生物量表现降低趋势;根冠比和侧根数量呈增长趋势,且各时期4个处理之间差异显著(表1,P<0.05)。根据计算4个土壤含水量条件下乌丹蒿苗期生长指标的上升及降低幅度来初步判定抗旱能力,与对照组相比其株高在第一时期轻度处理下的降低幅度最小为11.68%,在第三时期重度处理的降低幅度最大38.72%;茎粗在各处理期HS处理的降低幅度为最大依次是41.30%、27.33%、22.58%;根长在后期LS处理降低最小5.3%,在中期HS处理最大37.35%;与CK相比根冠比在3个生育期HS处理分别增加了4.5%、5.57%、8.12%;乌丹蒿幼苗总生物量在各时期HS处理下的降低最大分别为52.55%、46.49%、38.67%;侧根数量增长说明植株在土壤水分含量较低的情况下通过增加根系面积而吸收更多水分进而保护植株生长。详见表1。

表1 水分胁迫下乌丹蒿苗期生物量及生长指标的变化
注:不同小写字母表示差异显著(P<0.05)。
Note:Different lower case indicate significant differences at 0.05 level.
2.2 水分胁迫下乌丹蒿超氧化物歧化酶活性的变化
随着乌丹蒿幼苗在干旱环境胁迫下的生长其根系SOD活性均大于叶片且各处理间有极显著差异,在4片真叶期呈现缓慢增加趋势,在CK处理下根系SOD活性较叶片高62.29U/g·Fw、LS处理下根系活性高于叶片50.42U/g·Fw、MS处理下根系活性高出叶片36.20U/g·Fw、HS处理下根系活性高于叶片34.92U/g·Fw;在8片真叶期幼苗SOD活性在MS胁迫下达到最大值,在CK、LS、MS和HS处理下根系活性依次高于叶片47.54U/g·Fw、30.84U/g·Fw、44.29U/g·Fw和31.14U/g·Fw;在12片真叶期幼苗SOD活性均在MS处理下达到最大峰值,其CK、LS、MS和HS处理下的根系活性依次高于叶片30.69U/g·Fw、40.89U/g·Fw、68.48U/g·Fw和55.63U/g·Fw。如图1所示。
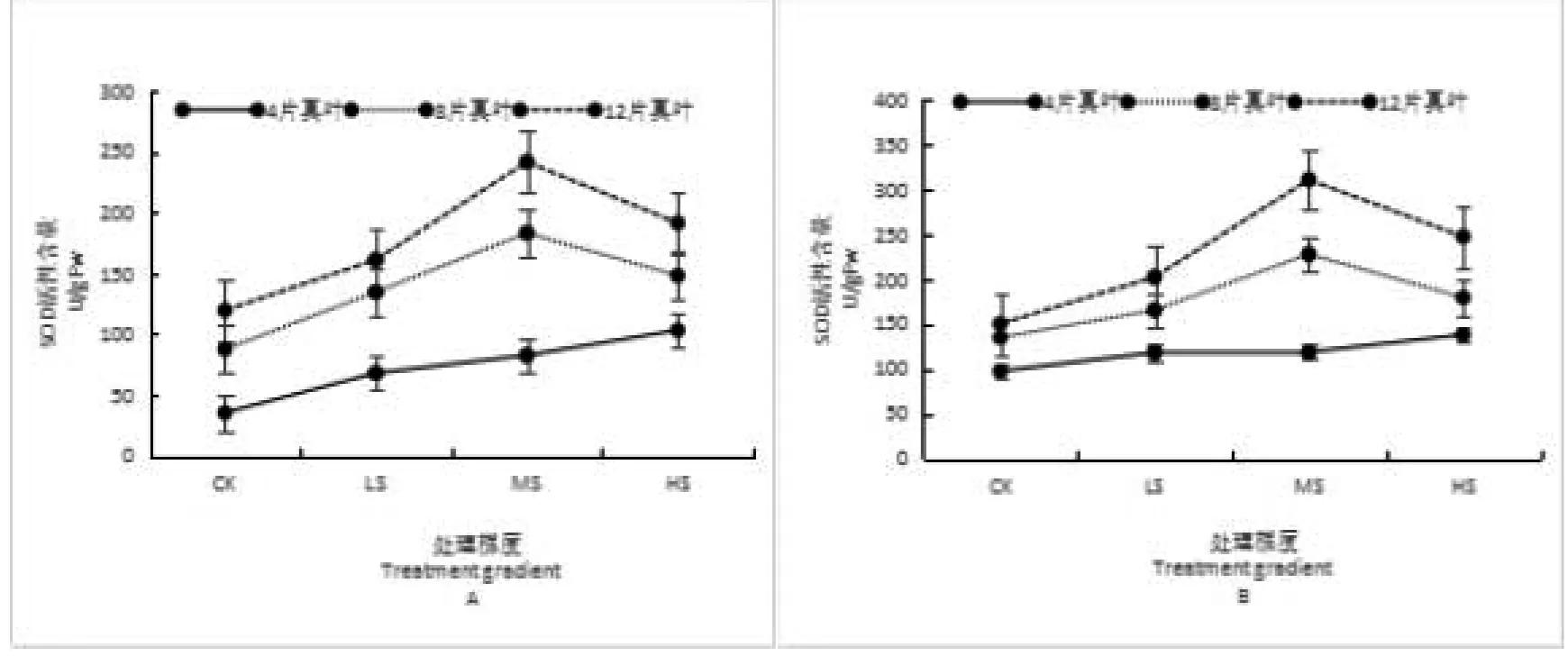
图1 水分胁迫对乌丹蒿幼苗叶与根SOD活性的影响
注:A为叶片,B为根系。不同小写字母表示差异显著(P<0.05)。
Note: A:Leaf;B:Root. Different lower case indicate significant differences at 0.05 level.
2.3 水分胁迫下乌丹蒿过氧化物酶活性的变化
随着土壤水分的缺失叶片和根系POD活性之间表现显著性差异,在4片真叶期表现缓慢持续上升,8、12片真叶期呈出先上升后下降的变化;且3个时期根系活性较高于叶片。在4片真叶期,其CK、LS、MS和HS处理下根系活性依次高于叶片67.79U/(gFw·min)、65.46U/(gFw·min)、54.26U/(gFw·min)和60.95U/(gFw·min);在8片真叶期,其CK、LS、MS和HS处理下根系活性依次高于叶片64.10U/(gFw·min)、51.28U/(gFw·min)、52.54U/(gFw·min)和59.14U/(gFw·min);在12片真叶期,其CK、LS、MS和HS处理下根系活性依次高于叶片52.16U/(gFw·min)、53.32U/(gFw·min)、64.08U/(gFw·min)和55.61U/(gFw·min)。如图2所示。
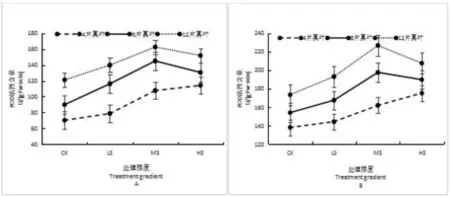
图2 水分胁迫对乌丹蒿幼苗叶与根POD活性的影响
注:A为叶片,B为根系。不同小写字母表示差异显著(P<0.05)。
Note: A:Leaf;B:Root. Different lower case indicate significant differences at 0.05 level.
2.4 水分胁迫下乌丹蒿过氧化氢酶活性的变化
乌丹蒿幼苗CAT活性随干旱胁迫时间的增加,其同一个时期、同一个处理下的根系CAT活性均高于叶片活性,且各处理间存在极显著性差异。在4片真叶期:叶片及根系CAT活性均呈现缓慢上升趋势,在HS处理达到最大值分别为70.82U/(gFw·min)和96.84U/(gFw·min),其CK、LS、MS和HS处理下的根系活性依次高于叶片32.52U/(gFw·min)、26.94U/(gFw·min)、24.23U/(gFw·min)和26.02U/(gFw·min);在8片真叶期幼苗CAT活性在MS胁迫下达到最大值,其CK、LS、MS和HS处理下根系活性依次高于叶片51.80U/(gFw·min)、49.59U/(gFw·min)、51.69U/(gFw·min)和49.83U/(gFw·min);在12片真叶期幼苗CAT活性仍在MS胁迫下达到最大值,其CK、LS、MS和HS处理下根系活性依次高于叶片55.25U/(gFw·min)、50.69U/(gFw·min)、48.67U/(gFw·min)和49.38U/(gFw·min)。如图3所示。
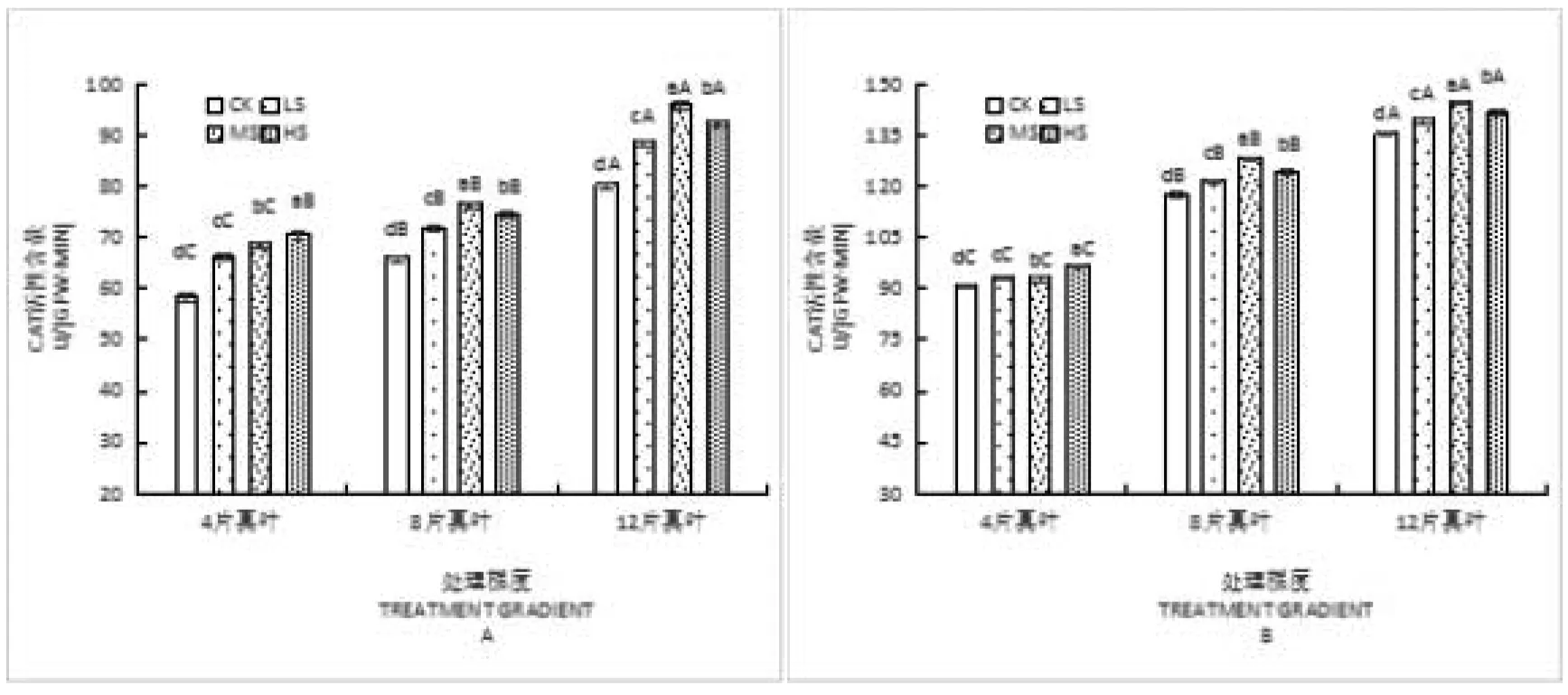
图3 水分胁迫对乌丹蒿幼苗叶与根CAT活性的影响
注:A为叶片,B为根系。不同小写字母表示差异显著(P<0.05)。
Note: A:Leaf;B:Root. Different lower case indicate significant differences at 0.05 level.
3 讨论
土壤水分亏缺会使植物体内水分吸收率与蒸腾之间的平衡失调,导致气孔关闭且蒸腾、光合速率下降而影响生长动态,这些对植物的最终影响会通过株高、茎粗、根长、根冠比和生物量的累积体现出来,也是评价植物抗旱性的重要指标〔10-12〕。乌丹蒿幼苗株高、茎粗和根长随着土壤含水量的减少都呈降低趋势,与对照组相比其重度胁迫处理下的值最低,表明水分缺失抑制了乌丹蒿苗期的生长,这与庞世龙等的研究一致〔13〕。3个时期的根系在中、重度干旱处理下表现出侧根数量增多,可见植物通过吸收周围土壤的水分来为植株提供水分,体现了乌丹蒿侧根发达的生物学特性〔4〕。乌丹蒿幼苗3个生育期的生物量均随胁迫时间的延长而减少,说明幼苗生长在干旱胁迫过程中受到明显的影响。植物在逆境中会通过调节地上地下部分生物量来适应环境,因此根冠比和生物量也成为了衡量植物在逆境中适应性的重要指标〔13〕,根冠比和生物量表现了地上地下部分干物质积累的关系,多项研究认为干旱胁迫会使根冠比增大、使生物量下降,与本研究结果相似。
水分亏缺导致植物体内活性氧和自由基的积累,进而打破体内自由基产生与清除的平衡,抗氧化酶可清除植物受到逆境胁迫时所产生的活性氧自由基,SOD清除活性氧自由基时需要POD和CAT的参与。进而保护植物膜。近年来,过氧化物酶、过氧化氢酶和超氧化物歧化酶的活性常用作植物抗旱性的生理指标〔14-16〕。田治国通过对万寿菊属苗期进行高温干旱双重胁迫条件发现逆境使植物体内生物膜受到伤害致活性氧增加〔17〕;陈超,陈香波分别对百脉根幼苗及夏腊梅叶片进行干旱胁迫发现,植物在受深度干旱胁迫时SOD和POD酶活性呈先升高后降低的趋势〔18,19〕。本研究发现,乌丹蒿幼苗初期三个抗氧化酶活性均呈上升趋势,但随后在8、12片真叶时期在土壤含水量下降到5%时,其叶片和根系SOD、CAT和POD活性由最初的上升趋势转变为下降趋势,且根系酶活性均高于叶片。由于胁迫方式能直接刺激根系导致根系酶活性高于叶片,可知土壤干旱对根系的损伤程度大于叶片。SOD活性上升幅度最大、其次是POD和CAT,值得注意的是CAT活性变化不大,说明SOD对水分胁迫的反应更为敏感,是乌丹蒿幼苗对干旱胁迫的主要保护酶。它是植物体内清除自由的最关键保护酶之一,通过增强幼苗O2-的歧化反应,促使SOD清除活性氧自由基、来维护幼苗氧代谢平衡。
4 结论
对乌丹蒿幼苗在其他条件相同的情况下进行水分梯度处理发现,生长过程中株高、茎粗、生物量等形态指标均受到不同程度的阻抑,但5%和15%的土壤含水量却促使侧根数量连续增加。
在3个时期中根系SOD、POD、CAT活性均高于叶片,4片真叶期酶活性随水分的减少而增加;8、12片真叶期在对照、轻度、中度处理组酶活性呈上升趋势,但在重度处理组活性降低,说明幼苗在重度胁迫下胞内保护酶系统失衡导致活性氧增加,加剧膜脂过氧化而损伤体膜使植株受到严重伤害。
综上所述,乌丹蒿在干旱环境的生长过程中通过调节形态及生理指数去适应环境,且拥有较发达的根系说明抗旱性较强。
〔1〕高海明.内蒙古水资源供需问题对策研究〔D〕.中国农业科学院,2007.
〔2〕刘球,李志辉,吴际友,等.红椿幼苗对干旱胁迫及复水生理响应的典型相关分析〔J〕.西北农林科技大学学报(自然科学版),2015,43(10):35-44.(2015/9/91).
〔3〕朝博,乌云,乌恩.气候变化背景下内蒙古草原水资源保护与可持续利用〔J〕.中国草地学报,2012,34(05):99-106.
〔4〕内蒙古植物志编辑委员会.内蒙古植物志,第四卷(第二版)〔M〕.呼和浩特,内蒙古人民出版社,1993:476-641.
〔5〕郑星星.科尔沁沙地三种沙蒿的形态和物候特征比较研究〔D〕.内蒙古师范大学,2014.
〔6〕YongcuiWang,Busso Carlos Alberto,DemingJiang,et al. The role of sexual vs. asexual recruitment of Artemisia wudanica in transition zonehabitats between inter-dune lowlands and active dunes in Inner Mongolia,China〔J〕. Solid Earth,2016,7(2):621-629.
〔7〕马君玲.典型沙生植物乌丹蒿的沙生适应机制——植冠种子库和粘液繁殖体〔J〕.中国农学通报,2014,30(07):1-4.
〔8〕邰丽华,王铁娟,敖登花,等.六种蒿属沙生半灌木染色体核型分析〔J〕.内蒙古师范大学学报(自然科学汉文版),2012,41(04):420-427.
〔9〕李合生.植物生理生化实验原理和技术〔M〕.北京:高等教育出版社,2000.
〔10〕杜彩艳,段宗颜,潘艳华,等.干旱胁迫对玉米苗期植株生长和保护酶活性的影响〔J〕.干旱地区农业研究,2015,33(03):124-129.(2015-06-24).
〔11〕吴敏,张文辉,周建云,等.干旱胁迫对栓皮栎幼苗细根的生长与生理生化指标的影响〔J〕.生态学报,2014,34(15):4223-4233.(2014-03-04).
〔12〕桑子阳,马履一,陈发菊.干旱胁迫对红花玉兰幼苗生长和生理特性的影响〔J〕.西北植物学报,2011,31(01):109-115.
〔13〕庞世龙,欧芷阳,申文辉,等.干旱胁迫对蚬木幼苗表型可塑性的影响〔J/OL〕.中南林业科技大学学报,2017,37(05):21-25.(2017-05-04).
〔14〕FaezehMahdavikia,Mohammad Jamal Saharkhiz,AkbarKarami. Defensive response of radish seedlings to the oxidative stress arising from phenolic compounds in the extract of peppermint (Menthapiperita L.)〔J〕. Scientia Horticulturae,2017,214(5):133-140.
〔15〕谢小玉,马仲炼,白鹏,等.辣椒开花结果期对干旱胁迫的形态与生理响应〔J〕.生态学报,2014,(13):3797-3805.
〔16〕Kalir A, Poljakoff-Mayber A. Changes in activity of malate dehydrogenase, catalase, peroxidase and superoxide dismutase in leaves of Halimioneportulacoides (L.)Aellen exposed to high sodium chloride concentrations〔J〕. Annals of Botany, 1981, 47(1): 75-85.
〔17〕田治国.万寿菊属植物耐热性与抗旱性的评价及生长生理特性的研究〔D〕.西北农林科技大学,2012.
〔18〕陈超,赵丽丽,王普昶,等.百脉根对干旱胁迫的生长、生理生态响应及其抗旱性评价〔J〕.水土保持学报,2014,28(03):300-306.
〔19〕陈香波,田旗,张丽萍,等.不同透光率和土壤含水量对夏蜡梅枝条生长量及叶片部分生理指标的影响〔J〕.植物资源与环境学报,2010,19(04):70-77.
