长江口海域春季浮游植物的年际变化*
2018-01-12林梅李扬
林 梅 李 扬
(华南师范大学生命科学学院 广州市亚热带生物多样性与环境生物监测重点实验室 广州 510631)
浮游植物是海洋生态系统的初级生产者, 具有种类多、数量大、繁殖快等特点, 是物质循环和能量转化中的关键环节, 具有重要的生态意义(Armbrust,2009; 高亚辉等, 2011; 孙晓霞等, 2011)。作为浮游动物、鱼、虾、贝等直接或间接的食物来源, 浮游植物还是很好的水产饵料生物, 其种类数和细胞丰度可以用来估计海洋渔业资源(高亚辉等, 2011)。此外, 某些浮游植物也是全球重要的藻华原因种(Fryxell et al,2004)。
长江口海域是我国重要的河口水域、传统渔场和水产品养殖基地, 也是我国重要的藻华高发区(Wang et al, 2009)。重要的地理位置和独特的环境特征使之成为各学科领域的理想研究区域(全为民等, 2005)。其中关于长江口浮游植物的研究, 已有诸多报道。尤其是近二十年来, 该海域藻华频发, 浮游植物研究持续备受关注。
另外, 随着人类活动和全球气候变化对海洋环境的影响日益显著, 环境变化下的生物响应逐渐成为研究热点。我国学者在胶州湾、台湾海峡等海域开展了相关研究, 并在部分海域总结性提出“营养盐结构改变”、“硅酸盐含量降低”、“甲藻优势度上升”等观点(林更铭等, 2011; 孙晓霞等, 2011)。长江口海域作为我国沿海的典型代表, 历史研究较为丰富, 但是类似的归纳总结还相对有限(Jiang et al,2014; 贾海波等, 2014), 直接的数据化证据相对较少, 未能从浮游植物群落结构的角度给出全面的响应解析。
基于上述原因, 本文于2014年5月在长江口海域采集浮游植物水样, 分析了浮游植物总量, 以及硅藻和甲藻群落的种类组成、细胞丰度、优势种类组成等。还收集整理了该海域 1986—2014年间的历史数据, 分析了硅藻、甲藻主要群落特征的年际变化, 以及硅藻优势种类细胞粒径的变化趋势, 以期能够发现规律性现象, 明晰浮游植物对海域环境变化的响应途径, 也为深刻理解海洋环境变化对浮游植物的影响提供数据支撑。
1 材料与方法
在 长 江 口 海 域 (27°93′N—32°5′N, 122°27′E—125°52′E)设置了 45个代表性站位(图 1), 于 2014 年 5月搭载“科学1号”科考船进行浮游植物水样的采集。每个站位分别采1m、10m、20m、50m水样各2.5L, 现场加 Lugol’s试剂进行固定。带回实验室后, 转移样品到量筒中, 静置沉淀24h以上, 然后用虹吸管轻轻吸掉上清液。如此操作 2次, 最终将样品浓缩至 10—15mL。从中取 0.1mL样品, 用浮游植物计数框在光学显微镜(Leica DM1000, Leica Microsystems, 中国)下进行浮游植物的种类鉴定和定量计数。
浮游植物种类鉴定主要参考《中国海洋浮游硅藻类》(金德祥等, 1965)、《中国海洋底栖硅藻类》(上、下卷)(金德祥等, 1982, 1991)、《中国海藻志》(第五卷硅藻门第一册中心纲)(郭玉洁等, 2003)、《南海甲藻(一)》(林永水等, 1993)、《中国海域甲藻 I》(杨世民等, 2014)、《Identifying Marine Phytoplankton》(Tomas,1997)。
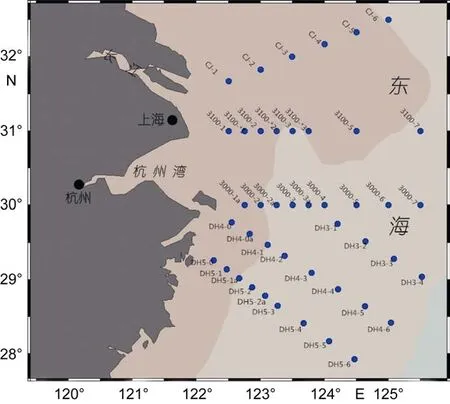
图1 长江口海域采样站位图Fig.1 Sampling stations in Changjiang River estuary
利用 Excel 2013软件的统计功能对历史数据进行分析, 并制作柱状图、散点图, 以及相关趋势性分析。运用Surfer 8.0作优势种水平分布图。
2 结果与分析
2.1 2014年春季长江口海域浮游植物群落结构
2.1.1 浮游植物的种类组成 在2014年长江口春季样品中, 共鉴定浮游植物105种, 其中浮游硅藻30属56种, 浮游甲藻19属41种, 以及隶属于金藻门、蓝藻门等的 8个种类。其中硅藻以根管藻属(Rhizosolenia)的种类最多, 鉴定到 8个种类, 占春季硅藻总种类数的 14.2%。其次是角毛藻属(Chaetoceros), 有 5个种类。甲藻以原甲藻属(Prorocentrum)的种类最多, 鉴定到7个种类。浮游硅藻的生态类型以半咸水为主, 中肋骨条藻(Skeletonema costatum)是典型代表; 浮游甲藻的生态类型以沿岸广布种为主, 东海原甲藻(P. donghaiense)是第一优势种。
2.1.2 浮游硅藻的优势种类组成及其水平分布
中肋骨条藻是浮游硅藻的第一优势种, 优势度达0.38。中肋骨条藻的出现频率不高, 主要出现在部分站位的表层水体, 但是其细胞丰度普遍较高, 占浮游植物总细胞丰度的 85.6%。拟菱形藻(Pseudo-nitzschiaspp.)是第二优势种, 优势度为0.04。拟菱形藻的出现频率较高, 在大部分站位均有检出,但细胞丰度不高, 主要分布在表层和10m水层。长江口春季浮游硅藻优势种组成详见表1。
在表层水体中, 中肋骨条藻是第一优势种, 细胞丰度最高达 1863.8×103cell/L, 平均丰度值为18.42×103cell/L, 主要集中在近岸海域, 124°E以东站位几近没有。水平分布图(图2a)显示, 细胞丰度高值区位于长江入海口, 与以往报道的高值区较为接近(高亚辉等, 2003; 谢文玲, 2006)。拟菱形藻是第二优势种(图 2b), 细胞丰度最高值为 41.54×103cell/L, 位于舟山定海附近, 而在长江入海口的分布较少。

表1 长江口春季浮游硅藻优势种组成Tab. 1 List of dominant diatom species in Changjiang River estuary
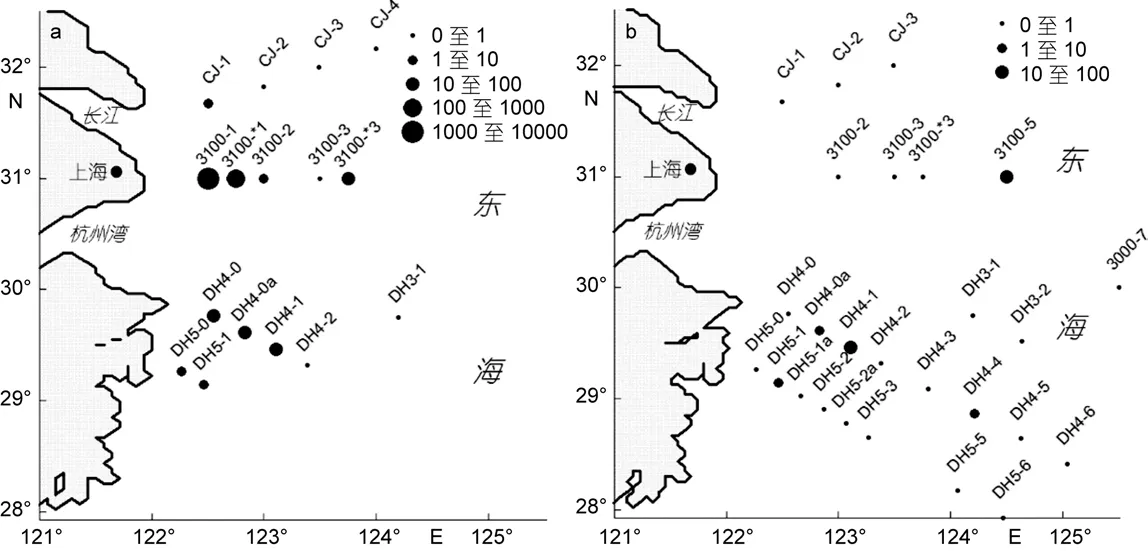
图2 表层水体中浮游硅藻优势种的水平分布图Fig.2 Horizontal distribution of dominant diatom species at water surface
2.2 长江口海域浮游植物年际变化
2.2.1 相关历史资料 由于长江口重要的地理位置和特殊的环境特征, 关于该海域浮游植物的历史研究较为丰富, 尤其是近二十多年的数据相对连续。但是由于年份跨度大, 以往研究在采样和处理方法、观察和鉴定技术、统计学手段上均存在差异; 另外随着科学水平的进步, 对藻类分类学和系统学的认知也发生了改变, 造成不同年份的调查数据在可比性上多有偏差, 以上因素正是中长尺度年际变化规律不易建立的原因所在。本文集中整理了长江口海域在1986—2014年之间的历史文献资料(表 2), 选取其中的春季数据, 进行浮游植物的年际变化分析。需要说明的是, 由于采样方法的不同(网样和水样)、观察技术的差异(光学显微镜的精密度不同), 以及计数统计手段的变化(浓缩样抽样计数法或 Utermöhl计数法等), 导致浮游植物细胞丰度数据的跳跃度较大, 缺乏可比性。此外, 因部分资料数据不完整, 本文将不就细胞丰度的年际变化展开讨论, 而将分析重点放在浮游植物种类数、硅藻和甲藻种类数贡献率、浮游硅藻优势种细胞粒径等三个方面。
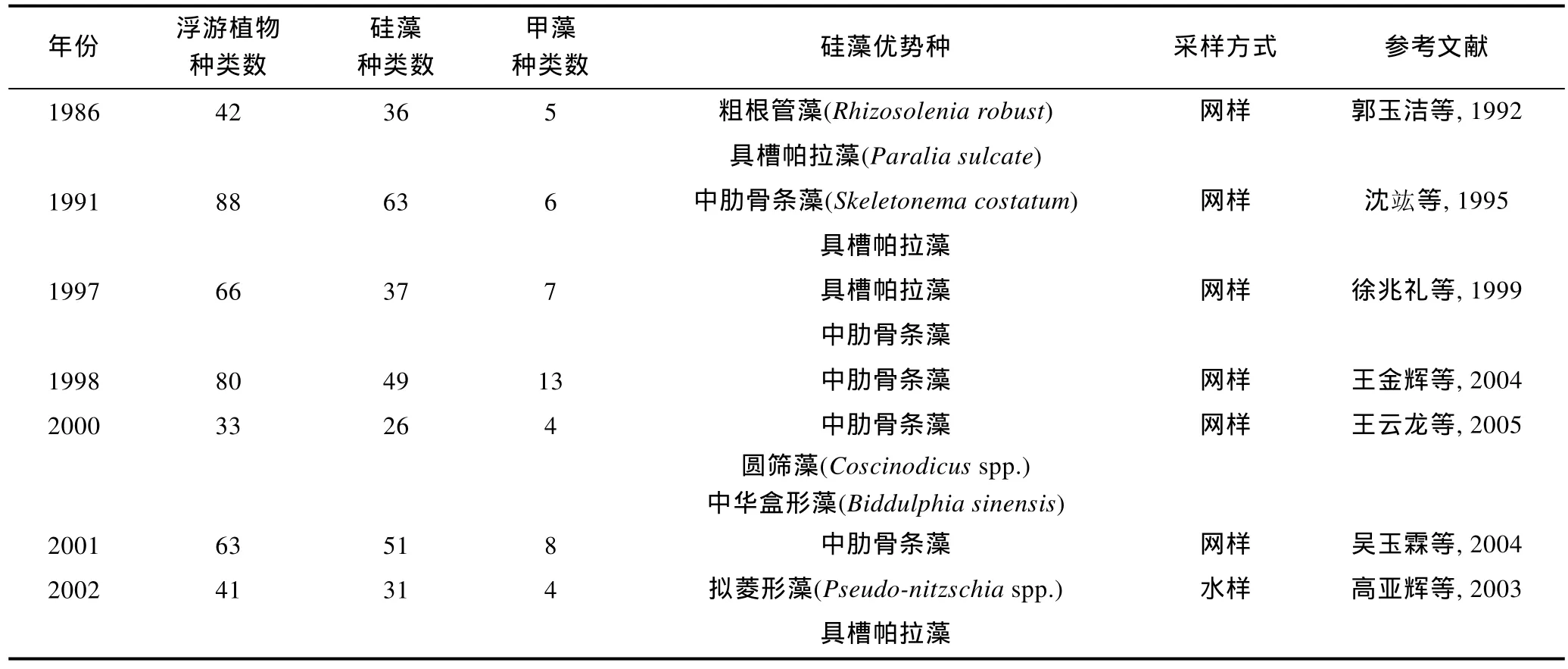
表2 长江口春季浮游植物的历史数据Tab. 2 Historical data of phytoplankton assemblage in Changjiang River estuary in spring during 1986—2014
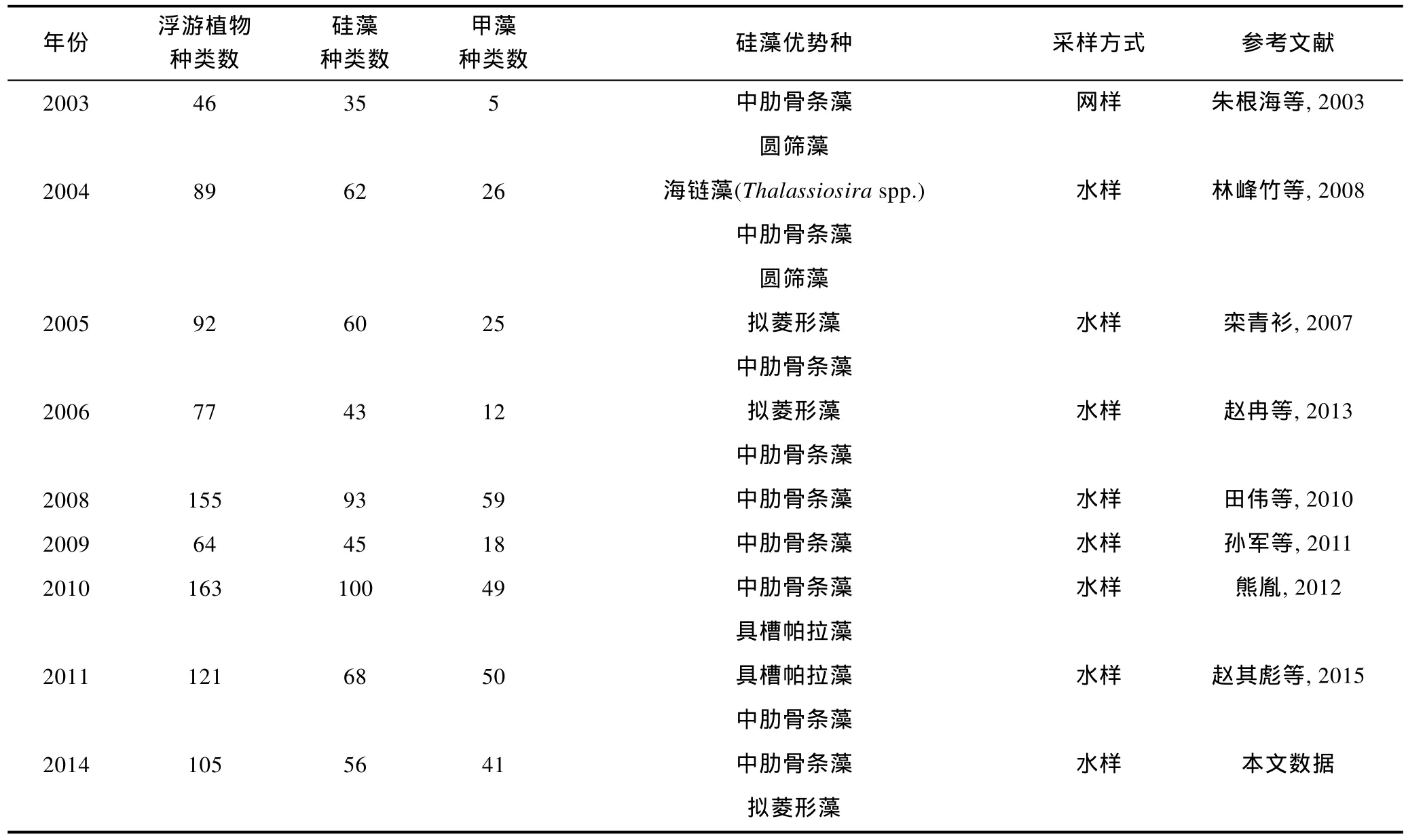
续表
2.2.2 浮游植物种类数的年际变化 按照采样方法的差异, 本文归类整理了春季浮游植物的网样数据和水样数据, 分别比较了其种类数的年际变化。
网样数据中(图 3a), 浮游植物种类数最高为 88种(1991年), 最低为33种(2000年); 浮游硅藻种类数呈现相同的变化规律, 最高有63种(1991年), 最低为26种(2000年); 浮游甲藻种类数最高为 13种(1998年), 最低仅有4种(2000年)。水样数据中(图 3b), 浮游植物最高种类数达到 163种(2010年), 其次是 155种(2008年), 最低为41种(2002年); 浮游硅藻种类数的年际变化基本相似, 最高有100种(2010年), 其次为93种(2008年), 最低为31种(2002年); 浮游甲藻种类数最高为59种(2008年), 其次为50种(2011年), 最低仅有 4种(2002年)。综上结果可以看出, 浮游硅藻的种类数均高于浮游甲藻, 但是浮游植物、浮游硅藻、浮游甲藻的种类数的年际变化趋势均不明显。水样数据(图3b)显示浮游植物、浮游甲藻的种类数均呈现不同程度的上升趋势, 其中浮游甲藻的上升趋势更为明显。
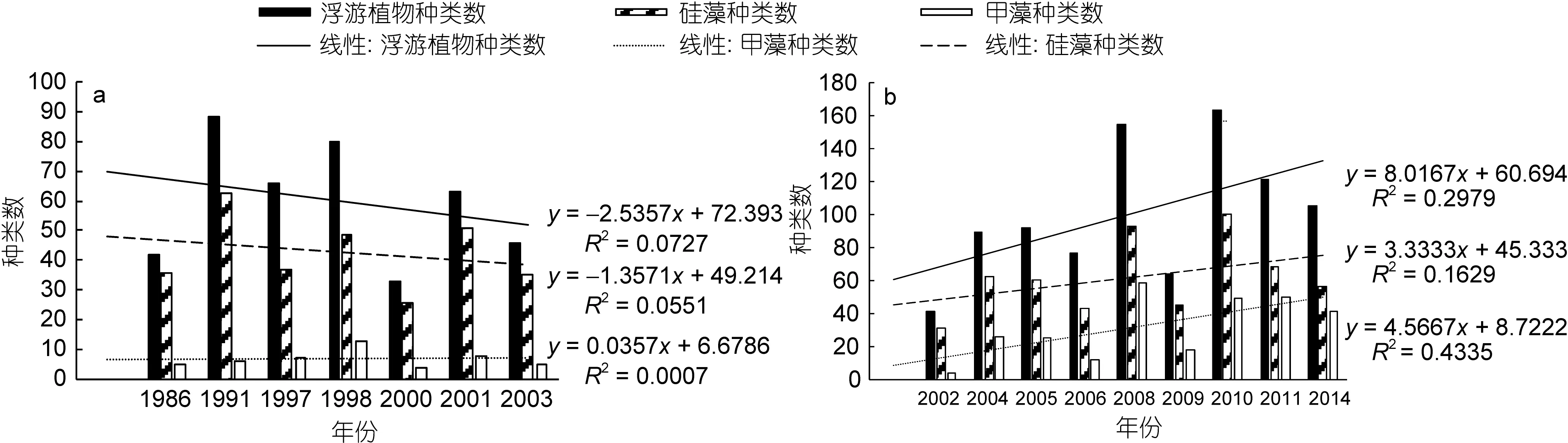
图3 长江口春季浮游植物种类数的年际变化Fig.3 Changes in species number for phytoplankton in Changjiang River estuary in spring during 1986—2003
2.2.3 硅藻、甲藻种类数占有比例的年际变化 从图3可知, 虽然采样方法不同, 但浮游硅藻和浮游甲藻依然是长江口春季浮游植物的重要组成, 其中浮游硅藻稳居浮游植物第一大类群。但是若以种类数占有比例作为分析依据(图4), 则呈现出另一个有价值的趋势。
在网样数据中(图4a), 1986年春季浮游植物的种类数为42, 其中硅藻为36种, 所占比例达到 85.7%;甲藻仅为5种, 所占比例为11.9%。2003年春季浮游硅藻种类数的占有比例为 76.1%, 甲藻为 10.9%。图4a显示在 1986—2003年间, 浮游硅藻、浮游甲藻在浮游植物中比例较稳定, 变化趋势不明显。但在水样数据中(图4b), 2002年春季浮游植物的种类数为41,其中硅藻为31种, 所占比例达到75.6%; 甲藻仅为4种, 所占比例为 9.8%。2008年春季浮游硅藻种类数的占有比例下降为60%, 甲藻则上升为38.1%。再到2011年春季, 硅藻种类数的占有比例进一步下降到56.2%, 甲藻则上升到41.3%。虽然浮游硅藻种类数的占有比例在 56%—75.6%之间波动, 始终处于最大贡献类群, 但整体而言呈下降趋势。而浮游甲藻种类数的占有比例则呈现大幅上升的趋势, 且趋势明显。这说明硅藻和甲藻在浮游植物中的重要性发生了变化,浮游硅藻的重要性逐渐下降, 而浮游甲藻的重要性正在快速上升。
考虑到种类数占有比例是样品数据的自身对比,其受采样方法的影响相对有限, 故本文综合了网样和水样数据(图4c), 亦呈现与水样数据相一致的变化趋势, 即“浮游硅藻的重要性逐渐下降, 而浮游甲藻的重要性显著上升”。
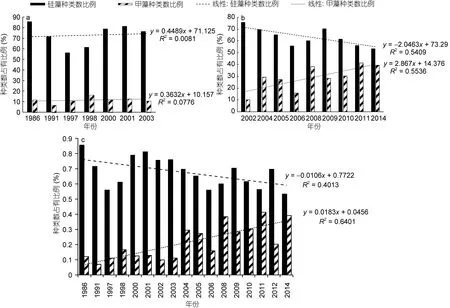
图4 长江口春季硅藻和甲藻种类数占有比例的年际变化Fig.4 Continuous variations in species number proportion for diatoms and dinoflagellates in total phytoplankton in Changjiang River estuary in spring during 1986—2014
以甲藻和硅藻之间的种类数比值为分析依据,亦可得出同样的结论。网样数据(图5a)中, 该比值呈现逐渐上升的趋势, 而在水样数据(图 5b)中, 该比值的上升趋势更为显著。
2.2.4 浮游硅藻优势种类细胞粒径的年际变化
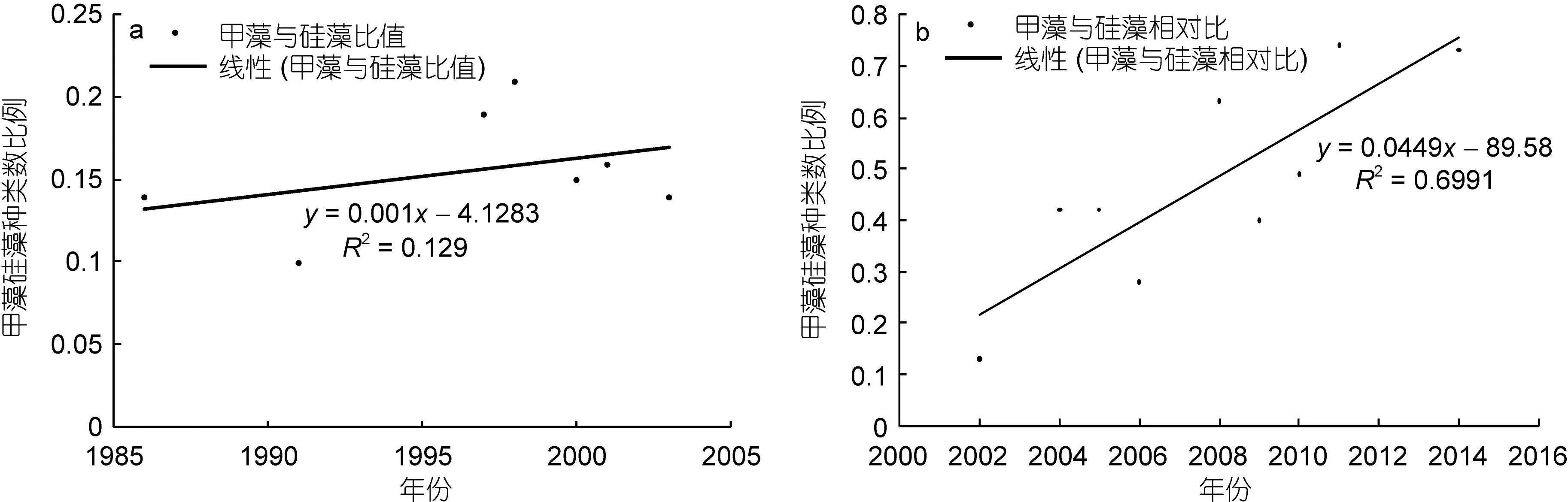
图5 甲藻与硅藻种类数比值的年际变化Fig.5 Changes of the ratio of dinoflagellate species number to diatom species number in Changjiang River estuary in spring during 1986—2014
本文以优势度大于 0.01作为选取浮游硅藻优势种类的标准, 建立了长江口春季浮游硅藻优势种类的年际数据(表 2)。由表 2可知, 中肋骨条藻是该水域最为常见的优势种, 在多个调查年份均有报道, 该种较为集中地分布在长江入海口。中肋骨条藻常以链状群体存在, 是近岸半咸水种的典型代表。其次, 拟菱形藻、具槽帕拉藻(Paralia sulcate)也是常见的优势种, 它们也常以链状群体生活。此外, 圆筛藻(Coscinodiscusspp.)、海链藻(Thalassiosiraspp.)、盒形藻(Biddulphiaspp.)等种类也偶有报道。
结合历史资料中对浮游硅藻优势种类的个体参数记录, 统计上述优势硅藻种类的平均细胞直径。需要说明的是, 由于 2000年之前的文献较少有细胞个体大小的记载, 而2000年之后的相关记录较为全面,因此本文仅对 2000—2014年间的硅藻优势种类进行了细胞粒径的统计分析。假设藻细胞均为球形(Sheldonet al, 1972), 制作硅藻优势种类细胞直径的年际变化图, 并进行线性拟合(图 6)。图 6显示浮游硅藻优势种类的细胞粒径呈现逐渐下降的趋势, 说明长江口优势硅藻的细胞个体越来越小, 其平均直径由 96μm(2000 年)降到 6μm(2014 年)。2000 年、2002—2004年间长江口硅藻优势种类中还有细胞个体较大的盒形藻和圆筛藻, 而在之后的调查中, 再未有此类细胞个体较大的硅藻种类进入优势种行列, 而细胞个体较小的中肋骨条藻、拟菱形藻、具槽帕拉藻则更多地占据优势地位。
3 讨论
3.1 浮游硅藻和浮游甲藻种类数的年际变化
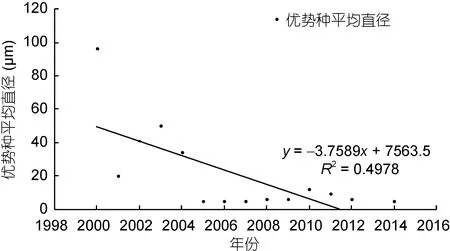
图6 长江口春季硅藻优势种类细胞粒径的年际变化Fig.6 Interdecadal variation in average cell diameter for diatom dominant species in Changjiang River estuary in spring during 1998—2016
曾有学者分析了1990—2002年间长江口海域浮游植物的年际变化(王金辉等, 2004), 认为浮游植物种类数呈现逐渐降低的趋势, 其中硅藻种类数占有比例下降, 而甲藻的种类数占有比例逐年上升。本文研究结论部分赞同上述观点。还有学者研究了在过去50年中长江口海域浮游植物的演替(Jianget al, 2014),认为浮游植物种类数, 以及硅藻和甲藻的种类数均呈现上升的趋势, 但由于甲藻的上升幅度更为显著,从而导致甲藻种类数占有比例逐渐上升, 这与本文的研究结论较为一致。需要注意的是, 若仅仅考虑1986—2003年间的网样数据(图3a), 浮游植物种类数确实呈现出缓慢下降的趋势。若针对水样数据进行相关分析(图3b), 浮游植物、浮游硅藻和浮游甲藻均呈现上升的趋势, 其中浮游甲藻的上升趋势更为明显。本文统计的年际范围更为广泛, 涵盖1986—2014年,或许更能反映真实情况。另一方面, 无论网样数据还是水样数据, 甲藻与硅藻之间的种类数比值均呈现上升趋势, 只是水样数据的上升趋势尤为显著, 这也从另一个角度支持了上述观点, 即浮游硅藻的重要性逐渐下降, 而浮游甲藻的重要性日益显著。与此同时, 研究手段和分析方法的进步, 以及对甲藻关注度的增加, 在一定程度上也促进更多的甲藻种类被发现。
虽然浮游硅藻和浮游甲藻的种类数都呈现上升趋势, 但是甲藻的上升幅度更为显著, 从而导致其占有比例逐年上升, 这可能与长江口海域环境的变化密不可分。有学者总结了长江口近半个多世纪以来的径流量和泥沙含量变化(辛明, 2014), 发现长江口径流量变化不大, 基本在 8.92×1011m3这一平均值上下波动, 但长江冲淡水所携带的泥沙含量逐渐降低, 由1.354×108t(2003 年)下降至 0.718×108t(2008 年)。入海输沙量的减少, 使得长江口海水透明度增加, 有利于浮游植物进行光合作用, 可促进浮游植物种类数和细胞丰度的增加。另外, 入海输沙量的减少, 降低了溶解态硅向海洋的输送通量, 造成长江口海域硅酸盐绝对含量和相对含量的降低, 逐渐形成了不利于硅藻种类竞争的生态环境。
长江口海域营养盐的变化趋势也验证了上述情况(林峰竹等, 2008; Jiang et al, 2014; 辛明, 2014)。长江口海域无机氮和磷酸盐入海通量有所增加, 而硅酸盐入海通量却有所降低。其中无机氮的浓度由22.9µmol/L(1963年)上升到122.9µmol/L(2007年), 累积增加了 436.7%; 磷酸盐的浓度则由 0.12µmol/L(1963年)升高到 1.58µmol/L(2007年), 累计增加了1217%。无机氮和磷酸盐的增加使长江口海域水体富营养化程度加重, 更有利于浮游植物的生长。而硅酸盐则由 303.7µmol/L(1963 年)下降到 94.4µmol/L(2007年), 累积下降约 70.0%, 硅酸盐的降低, 限制了硅藻群落的发展。
综上所述, 长江入海输沙量的减少, 加强了浮游植物的光合作用, 可促进其种类数和细胞丰度的增加。但是与此同时, 该海域无机氮和磷酸盐呈现增加趋势, 硅酸盐含量却大幅降低, 限制了硅藻群落的发展, 使得浮游硅藻在浮游植物中的占有比例越来越低, 而浮游甲藻的占有比例和重要性大幅增加。
3.2 浮游硅藻优势种类的变化特征
在以往报道的长江口浮游硅藻优势种类组成中,链状群体种类占据较大的比重, 如中肋骨条藻、拟菱形藻、具槽帕拉藻、海链藻等, 说明链状群体硅藻种类在该海域具有更好的环境适应性, 在营养盐、光照等环境资源竞争中更易占据优势, 较易成为优势种(何青等, 2009; Jiang et al, 2014; Valiela, 2015)。首先,群体细胞可以使细胞较长时间抵御下沉, 更好地停留在真光层进行光合作用。其次, 在对营养盐的摄取中, 群体细胞在下沉过程中较单细胞有更大的细胞营养盐通量(Karp-Boss et al, 1996); 另外, 群体细胞能够减少浮游动物的摄食(Fryxell et al, 2004)。因此,相较于单细胞种类, 群体细胞种类对环境的适应力更强。
硅藻优势种类的细胞粒径小型化也是一种适应环境的表现, 因为小个体种类具有更高的细胞表面积与体积比值, 更有利于对营养盐的吸收。环境因素对细胞粒径也有显著影响。有学者在室内半连续培养条件下, 分析了不同 Si∶P供应比例对藻细胞粒径的影响, 结果显示两者之间具有显著关系(Grover, 1989), 研究表明在高Si∶P条件下, 藻细胞有更大的细胞粒径, 而在低Si∶P条件下, 小个体的藻细胞更具有生存优势。同样, 在高营养盐条件下,硅酸盐的增加可以促进硅藻大个体细胞的生长, 而在低硅酸盐浓度条件下, 无机氮和磷酸盐的增加可以促进硅藻小个体细胞的生长。在对长江口营养盐的长期研究中, 辛明(2014)发现长江口海域硅酸盐含量呈现逐渐降低的趋势, 而无机氮和磷酸盐含量则逐年上升。大个体硅藻种类对硅酸盐的需求更高,对硅酸盐的依赖性也更强, 因此, 在硅酸盐含量降低的情况下, 小个体硅藻种类更易占据竞争优势。Grover(1989)的室内营养生理研究工作正好可为长江口海域浮游硅藻优势种类小型化的趋势提供合理的理论解释。
4 结语
(1) 2014年春季长江口海域共鉴定浮游植物105种, 其中浮游硅藻30属56种, 浮游甲藻19属41种。浮游硅藻以半咸水种为主, 中肋骨条藻是第一优势种。浮游甲藻以沿岸广布种为主, 东海原甲藻是典型代表。
(2) 在过去30年中, 浮游植物、浮游硅藻、浮游甲藻的种类多样性均呈现不同程度的上升趋势, 其中甲藻的上升趋势最为明显。甲藻种类数在浮游植物种类数中的占有比例呈上升趋势, 而硅藻所占比例呈下降趋势。
(3) 浮游硅藻优势种类的细胞粒径呈现小型化趋势。这与长江口海域硅酸盐含量减少、无机氮和磷酸盐增加、入海输沙量减少等因素密切相关。
致谢 感谢暨南大学赤潮与海洋生物学研究中心的吕颂辉教授和高健同学提供2014年春季浮游甲藻的相关数据。
王云龙, 袁 骐, 沈新强, 2005. 长江口及邻近水域春季浮游植物的生态特征. 中国水产科学, 12(3): 300—306
王金辉, 黄秀清, 刘阿成等, 2004. 长江口及邻近水域的生物多样性变化趋势分析. 海洋通报, 23(1): 32—39
田 伟, 孙 军, 樊孝鹏等, 2010. 2008年春季东海近海浮游植物群落. 海洋科学进展, 28(2): 170—178
朱根海, 许卫忆, 朱德第等, 2003. 长江口赤潮高发区浮游植物与水动力环境因子的分布特征. 应用生态学报, 14(7):1135—1139
全为民, 沈新强, 韩金娣等, 2005. 长江口及邻近水域富营养化现状及变化趋势的评价与分析. 海洋环境科学, 24(3):13—16
孙 军, 田 伟, 2011. 2009年春季长江口及其邻近水域浮游植物—物种组成与粒级叶绿素 a. 应用生态学报, 22(1):235—242
孙晓霞, 孙 松, 吴玉霖等, 2011. 胶州湾网采浮游植物群落结构的长期变化. 海洋与湖沼, 42(5): 639—646
杨世民, 李瑞香, 董树刚, 2014. 中国海域甲藻 I(原甲藻目、鳍藻目). 北京: 海洋出版社, 1—156
吴玉霖, 傅月娜, 张永山等, 2004. 长江口海域浮游植物分布及其与径流的关系. 海洋与湖沼, 35(3): 246—251
何 青, 孙 军, 2009. 长江口及其邻近水域网采浮游植物群落. 生态学报, 29(7): 3928—3938
辛 明, 2014. 长江口海域关键环境因子的长期变化及其生态效应. 青岛: 中国海洋大学博士学位论文, 44—102
沈 竑, 徐 韧, 王桂兰, 1995. 上海市海岛周围水域浮游植物的调查研究. 海洋通报, 14(4): 26—37
林永水, 周近明, 1993. 南海甲藻(一). 北京: 科学出版社,1—115
林更铭, 杨清良, 2011. 全球气候变化背景下台湾海峡浮游植物的长期变化. 应用与环境生物学报, 17(5): 615—623
林峰竹, 吴玉霖, 于海成等, 2008. 2004年长江口浮游植物群落结构特征分析. 海洋与湖沼, 39(4): 401—410
金德祥, 陈金环, 黄凯歌, 1965. 中国海洋浮游硅藻类. 上海:上海科学技术出版社, 1—230
金德祥, 程兆第, 刘师成等, 1991. 中国海洋底栖硅藻类(下卷). 北京: 海洋出版社, 1—437
金德祥, 程兆第, 林均民等, 1982. 中国海洋底栖硅藻类(上卷). 北京: 海洋出版社, 1—323
赵 冉, 孙 军, 宋书群, 2013. 2006年春季长江口及其邻近水域浮游植物. 海洋通报, 32(4): 421—428
赵其彪, 孙 军, 李 丹等, 2015. 东海低氧区及邻近水域浮游植物的季节变化. 生态学报, 35(7): 2366—2379
贾海波, 邵君波, 胡颢琰等, 2014. 三峡水库蓄水前后长江口及其邻近海域浮游植物群落结构的变化及原因分析. 海洋通报, 33(3): 305—314
徐兆礼, 白雪梅, 袁 骐等, 1999. 长江口浮游植物生态研究.中国水产科学, 6(5): 52—54
栾青衫, 2007. 长江口及其邻接水域浮游植物群集生态学研究.青岛: 中国海洋大学硕士学位论文, 13—19
高亚辉, 梁君荣, 陈长平等, 2011. 海洋硅藻多样性与生态作用研究. 厦门大学学报(自然科学版), 50(2): 455—464
高亚辉, 虞秋波, 齐雨藻等, 2003. 长江口附近海域春季浮游硅藻的种类组成和生态分布. 应用生态学报, 14(7):1044—1048
郭玉洁, 杨则禹, 1992. 长江口区浮游植物的数量变动及生态分析. 海洋科学集刊, 33: 167—189
郭玉洁, 钱树本, 2003. 中国海藻志 第 5卷 硅藻门 第 1册中心纲. 北京: 科学出版社, 1—493
谢文玲, 2006. 东海典型海域浮游硅藻群落结构与动态研究.厦门: 厦门大学博士学位论文, 22—61
熊 胤, 2012. 长江口及邻近海域浮游植物群落结构研究. 广州: 华南师范大学硕士学位论文, 15—38
Armbrust E V, 2009. The life of diatoms in the world’s oceans.Nature, 459(7244): 185—192
Fryxell G A, Hasle G R, 2004. Taxonomy of harmful diatoms. In:Hallegraeff G M, Anderson D M, Cembella A D eds. Manual on Harmful Marine Microalgae. Paris, France: United Nations Educational, Scientific and Cultural Organization,465—510
Grover J P, 1989. Effects of Si: P supply ratio, supply variability,and selective grazing in the plankton: an experiment with a natural algal and protistan assemblage. Limnology and Oceanography, 34(2): 349—367
Jiang Z B, Liu J J, Chen J F et al, 2014. Responses of summer phytoplankton community to drastic environmental changes in the Changjiang (Yangtze River) estuary during the past 50 years. Water Research, 54: 1—11
Karp-Boss L, Boss E, Jumars P A, 1996. Nutrient fluxes to planktonic osmotrophs in the presence of fluid motion.Oceanography and Marine Biology, 34: 71—107
Sheldon R, Prakash A, Sutcliff W, 1972. The size distribution of particles in the ocean. Limnology and Oceanology, 17(3):327—340.
Tomas C R, 1997. Identifying Marine Phytoplankton. San Diego,Chile: Academic Press, 1—858
Valiela I, 2015. Marine Ecological Processes. 3rd ed. New York,U.S.A.: Springer-Verlag, 364—440
Wang J H, Wu J Y, 2009. Occurrence and potential risks of harmful algal blooms in the East China Sea. Science of the Total Environment, 407(13): 4012—4021
