NAA与切根组合对江南油杉苗内源激素及根系构型的调节效应
2017-12-29何应会刘雄胜
何应会,蒋 燚,姜 英,刘雄胜,李 娟
(广西林业科学研究院 广西优良用材林资源培育重点实验室,广西 南宁 530002)
NAA与切根组合对江南油杉苗内源激素及根系构型的调节效应
何应会,蒋 燚,姜 英,刘雄胜,李 娟
(广西林业科学研究院 广西优良用材林资源培育重点实验室,广西 南宁 530002)
对江南油杉轻基质容器苗的组合控根进行了研究。采用L9(33)正交试验设计方法研究了在芽苗移栽期间进行切根和外源NAA处理对容器苗内源激素含量和根系构型的影响。结果表明,切根和外源NAA共同作用对苗木生长过程中的根径、根尖数、一级侧根均长以及全株鲜质量有显著影响。通过最优组合的评定,筛选出芽苗≥2 cm且幼叶未展期进行切根,留根3 cm或切根1/2+50 mg/LNAA溶液处理为较优控根组合。组合控根有效地减少甚至杜绝了苗木盘根现象的发生。对9个处理4个测定期的内源ZR,ABA,IAA和GA3含量主体效应的检验发现,切根时期、切根方式和NAA浓度对根系内源激素含量均起到了调控作用,同时,不同的控根措施会引起不同种类内源激素含量的变化,因此,组合控根措施的筛选才成为必要。
切根;根尖数;江南油杉;内源激素;根系构型
江南油杉Keteleeria cyclolepis为深根性树种,因根系发达、粗壮,具发达的皮层,能贮藏较多水分。该树种表现出极耐干旱、抗逆性强的特点,江南油杉也因此被作为优良的山地造林树种推广应用[1]。人工容器育苗时主根生长非常快,任其生长可达25 cm以上,根域限制使其主根长而孱弱、畸形,上山造林后树种优点将很难显现。切根是苗木培育过程中干扰根系最为常见的技术[2],通过不同时期的苗木切根,能够调控根系及地上部分的生长,最终提高苗木质量[3]。在马尾松Pinus massoniana[4]、湿地松Pinus elliottii[5]、江南油杉[6]等树种上的研究表明,将芽苗的胚根切除 1/2 有利于苗木质量和造林效果的提高。本试验的目的之一,进一步研究切根时期、切留根长度以及NAA浓度组合控根对江南油杉容器苗根系构型的影响,考察3因素的主效应并筛选最佳组合,从而改善江南油杉容器苗根系质量。另外,内源激素对植物生长发育与形态建成具有调控作用[7],对组合控根影响江南油杉生长的生理途径进行深入探讨,将有助于揭示江南油杉根系调控发生的内在规律。
1 材料与方法
1.1 试验材料
江南油杉球果采自广西贺州市八步区大桂山林场(110°45′13″E,24°09′11″N,平均海拔195 m)。球果采回后立即进行脱籽处理,净种后装布袋冷藏(3 ℃~5 ℃),种子饱满率87%。育苗试验在广西林科院科研苗圃进行,于2016年2月,浸种、消毒后进行沙床催芽,田间发芽率为82%。育苗基质配方为:腐殖表土40%、椰糠45%、腐熟的有机杂肥5%以及珍珠岩10%以体积比混合,轻基质无纺布容器规格12 cm×14 cm。所用控根试剂为萘乙酸可溶性粉剂,总有效成分含量2%。
1.2 试验设计、调查方法和采样方法
试验设计:采用3因素、3水平正交试验设计L9(33),设置9个江南油杉组合控根育苗试验处理,田间采用随机区组排列,每小组苗木75株,3次重复,试验因子、水平及说明详见表1。于2016年3月中旬进行芽苗切根移栽试验,苗木移栽后,采用常规田间管理方式。数据资料为苗木移栽后2个月内的各项数据。
激素测定及采样方法:切根移栽15 d起定期进行根系取样。分别取各处理幼苗10株,用水冲洗干净并用滤纸吸干,剪取根尖部分(5 cm),样品量0.2 g,并随即用锡箔纸包裹并立即投入液氮罐中,适时用干冰保存运送至中国农业大学作物化学控制研究中心分析测定。采用间接酶联免疫吸附法(ELISA)[8],测定样品中的脱落酸(ABA)、玉米素核苷(ZR)、吲哚乙酸(IAA)、赤霉素(GA3)含量。
根系的调查及采样方法:激素测定取样的同时,各处理另随机取15株,测定根径、一级侧根数及长度。用winrhizo根系构型软件(分辨单位0.01 mm)统计根尖数。移栽后2个月测定苗木鲜质量,并统计各处理分叉根形成比例和盘根比例,以单株记录数据。
1.3 统计分析
采用 Excel和SPSS16.0软件对数据进行处理。运用One-way ANOVA单因素方差分析、univariate正交试验方差分析以及SNK多重比较法对数据进行运算。
2 结果与分析
2.1 试验苗根系生长情况
将各试验组合相应生长参数的算术平均值及F检验结果列于表2(单因素方差分析),根系主要参数的主体间效应检验结果列于表3(正交试验方差分析),由表数据可以看出:
2.1.1 根径、根尖数和一级侧根平均长度的对比
在芽苗移栽时,切根和外源激素NAA共同作用对苗木生长过程中的根径、根尖数、一级侧根的平均长度以及全株鲜质量都有显著影响(表2)。主体间效应的F检验结果表明(表3),不同切根方式间江南油杉苗的根径和一级侧根平均长度有极显著差异(F值分别为35.045和9.578,P=0.000<0.01),切根能够显著提高根径的粗度和增加一级侧根的生长长度,尤其是留根3 cm(B2处理)效果更明显,但对根系总的根尖数影响不显著。切根时期和NAA浓度均对根尖数有极显著影响(F值分别为8.978和21.011,P=0.000< 0.01),当茎芽高度<2 cm时,不进行切根和NAA溶液处理芽苗(处理3),移栽后根尖数显著低于其他处理。同时发现,不进行切根的处理(A1B3C3、A2B3C1和A3B3C2)虽有相当好的发根数量,但一级侧根平均长度值较小,控根组合能扩大根幅。不同切根时期和不同NAA浓度对根径、一级侧根平均长度的影响并不显著。由此可知,根系的不同生长指标其主体影响因素并不完全一致。

表2 各试验组合的主要根系参数统计Table 2 The test statistics of the main root parameters in each group
2.1.2 NAA和切根处理最优组合的评定
进一步对以上(表3)有显著影响效果的因素进行不同水平间的多重比较(SNK法),并绘制出主效应因素不同水平间的试验数据直观图(见图1),从3个因素中筛选得到最佳控根组合A2B1C2和A2B2C2,即切根时期为茎芽≥2 cm,幼叶未展期,切根方式为留根3 cm或切根1/2,NAA使用浓度为50 ppm时为最佳。因考虑实际操作的简便和高效性,剪根时以留根3 cm为宜。
2.1.3 试验苗苗木质量的对比
苗期生物量的积累以及根系质量是检验苗木总体质量的重要参数。表2中各试验组合对应的全株鲜质量、分叉根株数比、盘根株数比这3项指标能够较好地反应苗木质量。从数据可以看出,处理A2B1C2具有最高的生物量,与最优组合的评定结果表现相一致。结合表2和图2可以看出,不切根的处理(A1B3C3、A2B3C1和A3B3C2)盘根现象较为严重,尤其是A2B3C1处理,盘根率达到了60%,非常不利于苗木生长。通过切根,能够使一定比例的苗木主根形成二叉分枝的状态(见图2中的A2B1C2)。分叉根的形成,可以打破主根伸长生长的顶端优势,增加侧根级数以及萌发面积,且根系构型更合理,有效地减少甚至杜绝了苗木盘根现象的发生。
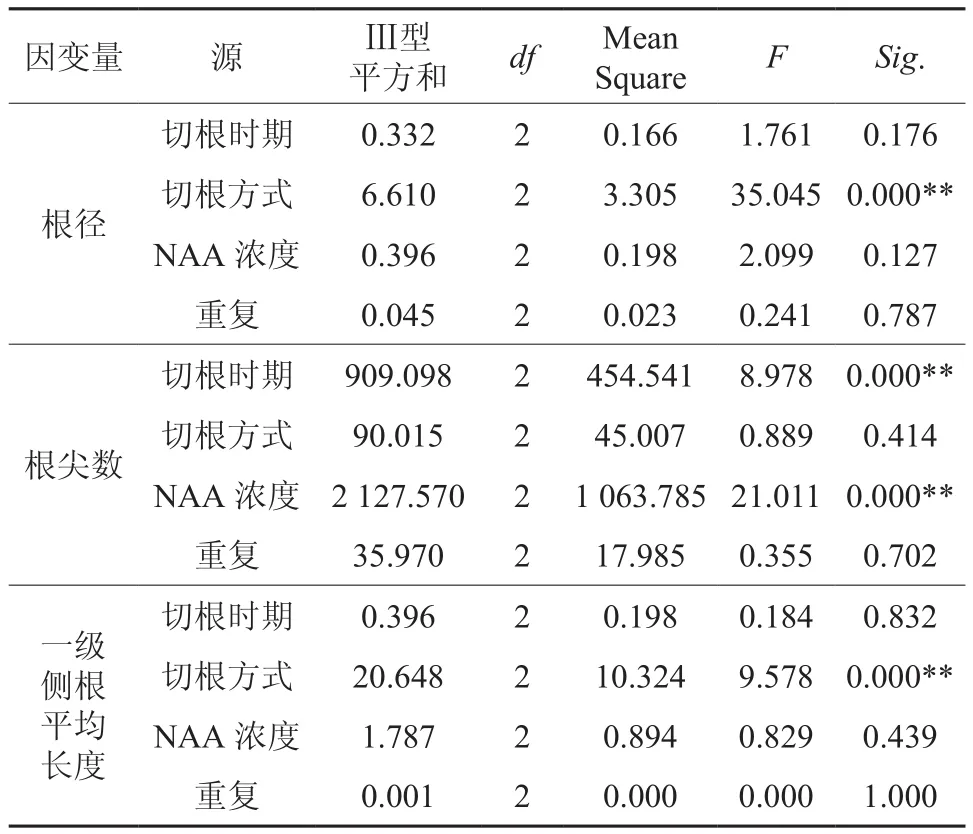
表3 方差分析结果(主体间效应的检验)Table 3 The result of variance analysis (Tests of betweensubjects effects)
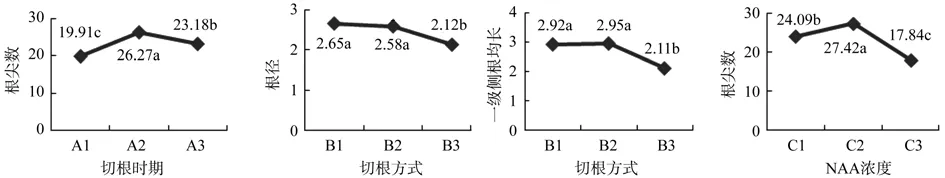
图1 各因素不同水平间的多重比较Fig.1 Multiple comparison between different levels of various factors
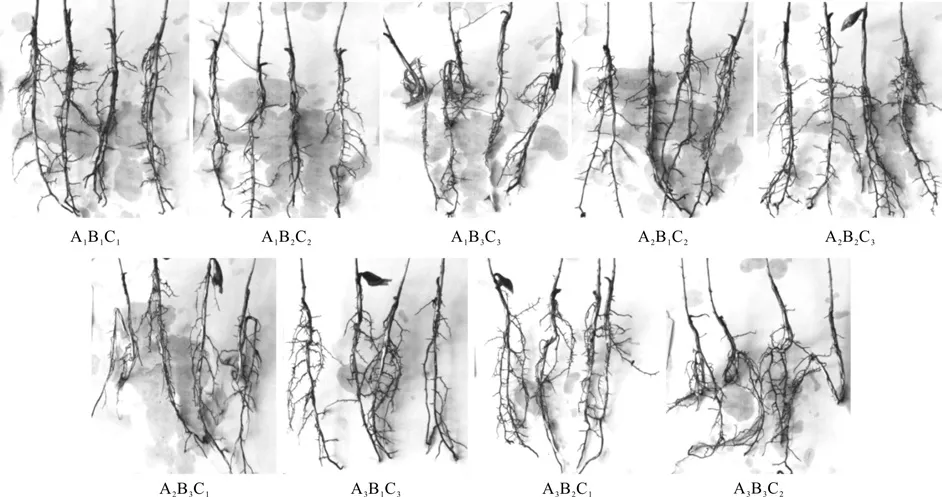
图2 NAA和切根处理后苗木根系外观表现的比较Fig.2 Contrast of seedling root morphological index after NAA and root cutting processing
2.2 根部内源ZR、ABA、IAA和GA3含量测定结果
2.2.1 ZR、ABA、IAA和GA3含量动态变化情况
控根组合对幼苗生长过程中根系内源ZR含量的影响结果见图3A。试验处理15 d时,ZR含量处理间差异不显著,随着处理时间的延长,差异逐渐显现。处理3(A2B1C2)在30 d和60 d时ZR含量均显著高于其他处理 [含量为 5.26±0.08 μg·g-1和5.49±0.12 μg·g-1],45 d 时 处 理 1(A1B1C1) 的 ZR值最高[4.34±0.18 μg·g-1]。在整个测定期中,处理1(A1B1C1)和处理3(A1B3C3)的ZR含量波动相对较大,但峰值出现却刚好相反;处理8(A3B2C1)和处理9(A3B3C2)几乎无太大变化。
控根组合对幼苗生长过程中根系内源ABA含量的影响结果见图3B。切根移栽后15 d处理间ABA含量差异达到最大(F值为369.943,P=0.000<0.01),此时ABA含量较高的处理依次为2[74.07±0.54 μg·g-1]、4[100.91± 1.05 μg·g-1]、9[109.29±2.12 μg·g-1]随着处理时间的延长,处理间的差异仍显著,但ABA含量差值范围缩小。对各测定期内所有处理计算均值得出,切根移栽后的15 d内源ABA含量基本达到峰值,此后呈现先降低后小幅回升趋势。结合柱形图表现以及正交试验设计方差分析得出,外源激素NAA(C2水平)处理胚根是引起根系ABA含量在15 d时升高的主要因素(F=19.401**)。
控根组合对幼苗生长过程中根系内源IAA含量的影响结果见图3C。试验处理15 d时各处理IAA含量差异达到极显著(F值为130.513,P=0.000 < 0.01),其中处理 4[35.21±0.11 μg·g-1]、5[29.55±0.54 μg·g-1]、9[32.22±0.18 μg·g-1]含量较高,到60 d时这3个处理IAA含量有所下降,而其他6个处理IAA含量有所上升。整个测定期各处理取均值发现,IAA含量仍以处理4、5和9为高。
控根组合对幼苗生长过程中根系内源GA3含量的影响结果见图3D。9个处理在不同测量期GA3含量差异均达到了显著水平,4个测定期中,30 d时GA3含量总体上为最低。处理3和处理6的峰值出现在 45 d时,分别为 6.31±0.15 μg·g-1和 6.10±0.12 μg·g-1,其余处理 GA3含量峰值均在60 d时。
2.2.2 主体效应的检验
对切根时期、切根方式和NAA浓度对根系内源激素含量变化主体效应的检验发现,3种因素对4种内源激素含量变化都有显著或极显著影响。仅列出15 d测定数据的方差分析结果(表4),ZR在15 d时处理间差异不显著,对ABA含量变化的调控作用依次为NAA浓度>切根方式>切根时期,对IAA含量变化的调控作用依次为NAA浓度>切根时期>切根方式,对GA3含量变化的调控作用依次为切根时期>切根方式>NAA浓度。表明,不同的控根措施会引起不同种类内源激素含量的变化,因此,组合控根措施的筛选才成为必要。
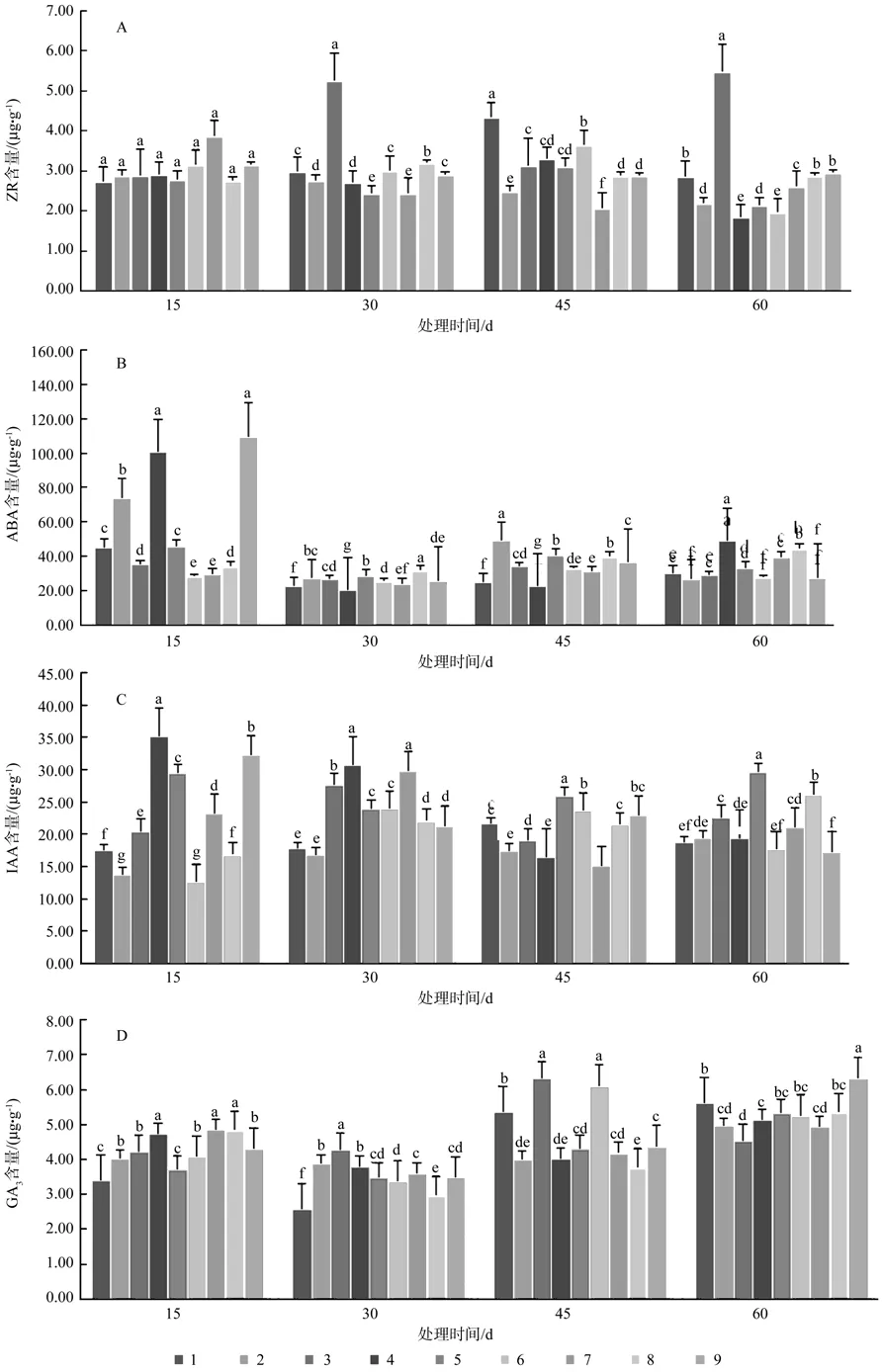
图3 NAA和切根处理对幼苗根内源激素变化动态的影响Fig.3 NAA and root cutting treatment effects on seedling root endogenous hormone changes dynamically
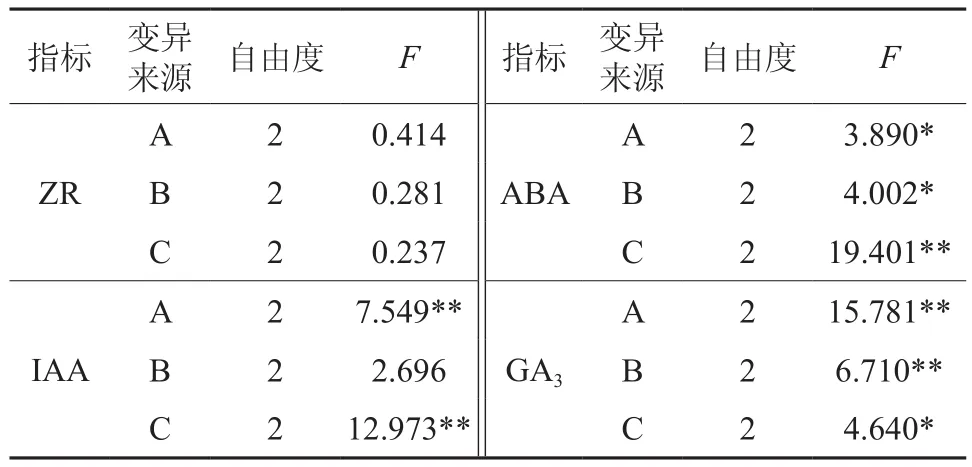
表4 15 d时内源激素数据的方差分析结果Table 4 The result of variance analysis of Endogenous hormone data within 15 days
3 小结与讨论
3.1 NAA和切根处理对根系构型的影响
苗期生物量以及根系质量被认为是检验苗木总体质量的重要参数,而根系在土壤中的分布特征极大地影响着植株获取可利用水肥资源的能力[9]。芽苗切根是指实生容器苗培育过程中,将沙床催播的芽苗在移栽前剪去部分胚根再移入容器的操作。已有研究表明,侧柏、白榆和刺槐苗木的生长量及一级侧根数与主根切断比例有关,且不同的切根比例其一级侧根集中范围有差异,这与本项目前期研究结果相一致[10],但进一步试验发现切根对根系总的根尖数的产生影响并不显著,但能够显著提高根径的粗度和一级侧根均长,而切根时期和外源激素NAA对根尖数有着极显著影响。以往的切根研究描述了诸如侧根数、侧根总长、分布区间等指标,对容器苗盘根现象的改善效果却未做出结论。本试验研究引入分叉根株数比、盘根株数比这2项指标来最终反映根系构型情况。研究得出,通过以切根为主的组合控根,能够使最高达53%的苗木主根形成二叉分枝的状态。分叉根的形成,可以阻断主根的伸长生长,促进增粗,增加侧根萌发面积,根系构型更合理,有效地减少甚至杜绝了苗木盘根现象的发生。试验从3个因素中筛选得到最佳控根组合A2B1C2和A2B2C2,即切根时期为茎芽≥2 cm,幼叶未展期,切根方式为留根3 cm或切根1/2,NAA使用浓度为50 mg/L时为最佳。因考虑实际操作的简便和高效性,剪根时以留根3 cm为宜。
3.2 NAA和切根处理对根部内源ZR,ABA,IAA和GA3含量的影响
幼苗生长期体内激素变化趋势是相当复杂和多向的,试验控制条件不同、测定时期的不同均会对数据结果产生影响[11-13]。根是细胞分裂素CTK的主要来源,且CTK 优先供应生长旺盛的部位[14],在已发现的30多种细胞分裂素中,玉米素(Z)和玉米素核苷(ZR)是高等植物体内主要的并可转运的细胞分裂素[15]。本试验在15 d时各处理间根系ZR含量无显著差异,此后波动较大,极可能与根茎叶各组织ZR含量的分配以及试验因素、水平对其转运速率的影响有关。ABA 被认为是一种最为直接的应激响应激素[16],在本试验中也得到了体现,15 d时处理间ABA和IAA含量差异极显著,但ABA含量随后都较大幅度下降,且各处理间差值范围缩小,而IAA含量一直保持着较大差异。本试验对9个处理4个测定期的内源ZR,ABA,IAA和GA3含量主体效应的检验发现,切根时期、切根方式和NAA浓度对根系内源激素含量均起到了调控作用,同时,不同的控根措施会引起不同种类内源激素含量的变化,因此,组合控根措施的筛选才成为必要。
本研究的研究涉及NAA浓度、切根方式和切根时期3种因素对江南油杉实生容器苗根系构型及内源激素的的影响,对于控根试剂的选择目前未能做到全面,赤霉素和生长素在该树种幼苗控根中是否有相同甚至更好的效果需试验验证。同时,控根处理后内源激素在根茎叶中的分配及运转,进而与高生长的关系还需进一步研究。
[1] 广西壮族自治区林业科学研究院. 广西树木志 [M]. 北京: 中国林业出版社, 2011: 11.
[2] Riley L E, Steinfeld D. Effects of bare root nursery practices on tree seedling root development: an evolution of cultural practices at J. Herbert Stone nursery [J]. 2005, New Forests, 32(2/3):107-126.
[3] Chauhan S K, Sharma G K, Charan S. Effect of root culturing and fertilizers on morphology and outplanting survival of Ulmus villosa seedlings[J]. Indian Forester, 2008, 134(11): 1493-1504.
[4] 林志鹏.切根育苗对马尾松苗木及幼林生长的效应[J].西南林学院学报, 2000, 20(4): 196-199.
[5] 罗盛健. 湿地松切根育苗及其造林对比试验[J]. 防护林科技 ,2006(4): 22 -24,37.
[6] 何应会, 蒋 燚, 刘 菲, 等.江南油杉切根移栽根系参数及其与生长的关系 [J]. 中南林业科技大学学报,2016,36(7):12-16.
[7] Lopez M A, Bannenberg G, Castresana C. Controlling hormone signaling is a plant and pathogen challenge for growth and survival[J]. Curr Opin Plant Biol,2008, 11(4) : 420-427.
[8] 郝建军, 康宗利,于 洋. 植物生理学实验技术[M]. 北京: 化学工业出版社, 2007: 124-133.
[9] 郭起荣,陈 红,冯 云,等 . 用根箱法研究毛竹实生苗根系构型 [J]. 中南林业科技大学学报,2014,34 (4):6-9,24.
[10] 杨喜田,王广磊,赵 宁,等 . 不同切根处理对林木幼苗根系侧根生长的影响 [J]. 河南农业大学学报 ,2010,44(2): 155-159.
[11] 章建红,陈志颖,阮 晓,等.DHAP 胁迫对天山云杉幼苗内源激素含量变化的影响[J].林业科学,2014,50(4):121-128.
[12] 杨喜田,陈久美,唐 妍,等.侧柏幼苗切根后根系内源激素含量的变化[J].河南农业大学学报,2011,45(1):66-70.
[13] 王秀英,张大惠,李恩彪.植物生长调节物质HKL-4对栽培黄花乌头幼苗根内源激素变化动态的影响[J].江苏农业科学,2010(6):386-387.
[14] 刘晓娟.根域限制对牡丹碳代谢和内源激素变化的影响[D].河南:河南农业大学,2012.
[15] Howell S H, Lall S, Che P. Cytokinins and shoot development[J].Trends in Plant Science, 2003, 8(9): 453-459.
[16] Sreenivasulu N, Harshavardhan V T, Govind G,et al.Contrapuntal role of ABA: does it mediate stress tolerance or plant growth retardation under long-term drought stress[J]. Gene,2012, 506(2): 265-273.
Regulative effect of the NAA and root-cutting combination on the endogenous hormone and root architecture ofKeteleeria cyclolepisseedlings
HE Yinghui, JIANG Yi, JIANG Ying, LIU Xiongsheng, LI Juan
(Guangxi Key Laboratory of Superior Timber Trees Resource Cultivation /Guangxi Zhuang Autonomous Region Forestry Research Institute, Nanning 530002, Guangxi, China)
A combined root control method over light media containerKeteleeria cyclolepisseedlings was studied. Applying an orthogonal experimental design in array L9(33), this research studied the effect of root-cutting and exogenous NAA treatments on contents of endogenous hormone and root architecture of container seedlings during their transplantation period. Analytical results indicate that rootcutting and exogenous NAA combination has jointly exerted signi ficant in fluences on root diameter, number of root tips, average length of first-order lateral root, and total fresh mass. Through evaluation, a relatively optimal combination, in which root cutting was conducted to retain root length by 3 cm when seedling length was ≥2 cm before the young-leaf expansion period, or the root was cut by 1/2, plus treatment with 50 mg/LNAA solutions, was achieved; the combined root control has effectively reduced or eliminated the occurrence of girdling seedling roots. A test on the effect of 9 treated subjects’ contents of endogenous ZR, ABA, IAA and GA3during 4 measurement periods was conducted, finding that the stages and methods of root cutting, as well as NAA concentrations, all have a regulative effect on the content of endogenous hormones in the root system; in the meantime, different root-control measures result in content variations in different types of endogenous hormones. Therefore, conducting screening on combined root-control measures is necessary.
root-cutting; root tips number;Keteleeria cyclolepis; endogenous hormone; root architecture
S791.15;S723.1
A
1673-923X(2017)08-0011-07
10.14067/j.cnki.1673-923x.2017.08.003
2016-12-20
国家林业公益性行业科研专项(201304108);广西林业科技项目(桂林科研 [2015] 第 33 号)
何应会,工程师
蒋 燚,教授级高级工程师,博士;E-mail:jy68@163.com
何应会,蒋 燚,姜 英,等. NAA与切根组合对江南油杉苗内源激素及根系构型的调节效应[J].中南林业科技大学学报,2017, 37(8): 11-17.
[本文编校:文凤鸣]
