大黄鱼腐败菌的低温致腐性及相互作用
2017-12-19杨冬英方慧萍
张 雯, 卞 丹, 杨冬英, 方慧萍, 倪 莉
(福州大学食品科学技术研究所, 福建省食品生物技术创新工程技术研究中心, 福建 福州 350116)
大黄鱼腐败菌的低温致腐性及相互作用
张 雯, 卞 丹, 杨冬英, 方慧萍, 倪 莉
(福州大学食品科学技术研究所, 福建省食品生物技术创新工程技术研究中心, 福建 福州 350116)
以挥发性盐基氮、 三甲胺和气味指纹图谱为表征, 探讨腐败希瓦氏菌(S.putrefaciens)、 静止嗜冷杆菌(P.immobilis)和荧光假单胞菌(P.fluorescens)在低温下对鱼肉的腐败性以及混合菌群在致腐过程中的相互作用. 研究结果表明, 腐败希瓦氏菌具有较强的产挥发性盐基氮特性, 主导腐臭气味; 荧光假单胞菌具有类似的腐败趋势, 但腐败性较腐败希瓦氏菌弱; 静止嗜冷杆菌不具有显著的产挥发性盐基氮等特性, 但能产生更为复杂的气味; 荧光假单胞菌和静止嗜冷杆菌在腐败初期对腐败希瓦氏菌的腐败趋势有抑制作用.
腐败菌; 腐败希瓦氏菌; 静止嗜冷杆菌; 荧光假单胞菌; 致腐性; 大黄鱼
大黄鱼(Pseudosciaenacrocea)是福建省重要的海水养殖鱼类, 养殖产量占全国总产量八成以上[1]. 其中, 90%大黄鱼是以冰鲜方式销售, 因此, 保持鱼的新鲜度是大黄鱼销售的重要环节. 研究结果证实大黄鱼的变质与细菌菌相的变化有关, 腐败希瓦氏菌和假单胞菌是其重要的腐败菌[2-5]. 本研究采用气相色谱(GC)和气相色谱-嗅觉测量法(GC-O)结合主成分分析, 探讨希瓦氏菌、 假单胞菌和冰鲜鱼体常见的嗜冷杆菌对冰鲜大黄鱼的致腐特性以及三者之间的相互作用关系.
1 材料与方法
1.1 材料
1.1.1 供试菌株
腐败希瓦氏菌(Shewanellaputrefaciens)A菌株, K菌株, O菌株; 荧光假单胞菌(Pseudomonasfluorescens)N菌株, 静止嗜冷杆菌(Psychrobacterimmobilis)C菌株. 以上菌株经16S rDNA和脂肪酸鉴定后, 置于30%的灭菌甘油中于-80 ℃条件下贮藏.
1.1.2 主要培养基及制备
牛肉膏蛋白胨培养基: 蛋白胨10 g, 牛肉膏3 g, 氯化钠5 g, 定容至1 000 mL, pH值为7.2~7.4, 121 ℃高压灭菌20 min.
1.2 方法
1.2.1 无菌鱼肉的制备
新1鲜大黄鱼清洗后, 去鳞、 鳍及内脏, 无菌水洗净后, 于超净台内用吸水纸将水吸干, 用解剖刀切取背脊处鱼肉, 去皮, 取大小厚薄相等的鱼块, 每块约5 g. 将鱼块表面用酒精棉擦拭, 经紫外线照射10 min.
1.2.2 接菌贮藏与鲜度动态分析
将供试菌株接菌于牛肉膏蛋白胨液体培养基扩大培养24 h后, 取适量发酵液, 加入无菌水稀释配制成106CFU·mL-1菌悬液. 将无菌鱼块浸泡于菌悬液中约20 s后移至无菌培养皿中, 4 ℃贮藏, 以不接菌的无菌鱼块为对照, 跟踪测定鱼肉中挥发性盐基氮和三甲胺含量, 测定其鲜度变化.
1.2.3 鲜度测定
1) 挥发性盐基氮(TVBN)测定. 参考水产行业标准《水产品中挥发性盐基氮的测定(SC/T 3032-2007)》[6].
2) 三甲胺气相色谱测定. 取40 g鱼肉样品均质, 用50 mL 10%三氯乙酸溶液(体积分数, 下同), 超声波振荡浸提1 h后过滤, 滤渣再用50 mL 10%三氯乙酸溶液提取, 合并2次滤液, 用10%三氯乙酸溶液定容至100 mL. 取该提取液4 mL于10 mL的具塞试管中, 加入1 mL的正庚烷, 2 mL 50%的KOH , 迅速塞住, 60 ℃下恒温水浴6 min, 剧烈震荡2 min后静置10 min, 取上层清液10 μL进样测定. 采用SE-30色谱柱; 柱温为50 ℃; 载气为N2(入口压=0.3 MPa); 检测器为氢火焰离子化检测器. 以标准盐酸标定过的三甲胺水溶液制备标准曲线.
1.2.4 鱼样挥发性气味的测定
1) 样品前处理. 预先准备好NaCl饱和溶液, 称取2 g 鱼肉样品(参照1.2.1, 鲜鱼样品无需酒精灭菌; 接菌样品采用106CFU·mL-1的菌悬液进行接菌, 在低温培养箱中4 ℃培养), 放入规格为15 mL的萃取瓶中, 加入5 mL的NaCl饱和溶液, 密封, 置于60 ℃水浴中, 磁力搅拌, 平衡5 min后, 采用PDMS-DVB两相萃取头, 萃取时间为30 min, GC解析时间3 min.
2) 色谱条件. 采用石英毛细管色谱柱(Agilent HP-INNOWax, 30 mm×0.32 mm×0.25 μm); 程序升温条件: 初始温度40 ℃, 保持5 min, 以5 ℃·min-1升温至240 ℃, 保持5 min; 检测器温度: 300 ℃; 进样口温度: 250 ℃; 分流进样: 分流比1∶1; 载气: N2; 载气流速: 1 mL·min-1; 部分载气进入FID检测器, 部分进入嗅闻装置(ODP).
3) 气相色谱-嗅觉测量法(GC-O). 利用气相色谱-嗅觉测量法检测气味特征时, 嗅闻人员熟悉鱼样和气味词汇. 进行嗅闻时, 记录嗅到香气时的保留时间以及气味类型. 为避免嗅闻人员产生嗅觉疲劳, 每人每次检测不超过10 min; 为减少主观偏差, 每个样品需要三个嗅闻人员检测. 不同人员对同一个样品在相同时间出现的气味可能出现不同的描述. 嗅闻小组通过讨论, 筛选和确定最终词汇和相应描述, 如表1所示.

表1 气味感官词及其描述Tab.1 Smell sensory words and the descriptions
数据分析采用Mintab 17对接菌不同菌株的鱼肉GC数据和GC-O数据进行主成分(PCA)分析, 判断不同菌株的致腐特征或鱼肉样品的鲜度.
2 结果与分析
2.1 鱼肉腐败菌的致腐性
腐败希瓦氏菌A产挥发性盐基氮的能力最强, 腐败希瓦氏菌K和O及荧光假单胞杆菌N产挥发性盐基氮的能力中等, 静止嗜冷杆菌C表现微弱, 见图1. 荧光假单胞杆菌N接菌鱼肉后期有很强的产三甲胺能力, 腐败希瓦氏菌A在接菌鱼肉前期(3 d内)较快产生三甲胺, 而腐败希瓦氏菌O和K在接菌鱼肉后期具有较强的产三甲胺能力, 静止嗜冷杆菌C在培养前期产少量三甲胺, 见图2.

图1 接菌鱼肉挥发性盐基氮的变化Fig.1 Changes in TVBN value of fish inoculated with spoilage bacteria

图2 接菌鱼肉产三甲胺测定Fig.2 Changes in TMA value of fish inoculated with spoilage bacteria
检测结果表明, 腐败希瓦氏菌和荧光假单胞杆菌是大黄鱼的重要腐败菌, 不同菌株对鱼肉的致腐性表现出一定差异. 腐败希瓦氏菌A的致腐性最强, 表现为产挥发性盐基氮和三甲胺的能力强; 荧光假单胞杆菌N和腐败希瓦氏菌O和K致腐性强, 表现为接菌鱼肉后期较强的产三甲胺能力; 静止嗜冷杆菌C致腐性弱, 产挥发性盐基氮和三甲胺能力差.
2.2 鱼体腐变的气味分析
新鲜鱼肉分为6个处理组, 单独接菌3组: 腐败希瓦氏K、 荧光假单胞菌N、 静止嗜冷杆菌C; 混菌两两接菌3组: 希瓦氏菌K和假单胞菌N、 希瓦氏菌K和嗜冷杆菌C、 假单胞菌N和嗜冷杆菌C; 并以不接菌细菌的鱼肉为对照. 将各处理组样品放置4 ℃贮藏, 于0、 3、 6、 9 d取样进行气相色谱-嗅觉测量法(GC-O)检测和气相色谱(GC)检测分析, 查明大黄鱼鱼肉腐变过程产生的气味类型和不同细菌引起鱼肉腐败过程产生的气味特征.
2.2.1 气相色谱-嗅觉测量法(GC-O)检测结果
GC-O检测和PCA法分析结果见图3~5. 不同处理组的鱼肉样品在腐变过程中产生的特征气味有明显差异.
新鲜鱼肉自然特征气味是咸味和腥味(海水鱼特有风味); 未接菌鱼肉在贮藏3 d后, 特征气味为酒味、 米味和腐烂味, 贮藏6 d和9 d特征气味是牛肉膏培养基味和尿骚味, 见图3(a). 与接菌样品相比较, 未接菌贮藏3、 6、 9 d的样品所产生的气味类别较为集中, 气味变化较小, 见图4.
接菌腐败希瓦氏菌K菌株后第3天和第6天的样品产生的特征气味为酸臭、 闷臭; 第9天样品特征气味为臭鸡蛋气味和腐烂味, 见图3(b). PCA分析显示接菌腐败希瓦氏菌K菌株的鱼肉在第3天与第6天气味类型接近, 说明希瓦氏菌致腐性强, 加速了鱼肉腐败进程.
接菌荧光假单胞菌N菌株的鱼肉贮藏3 d的特征气味为牛肉膏培养基气味, 贮藏6 d产生酸臭和草本味, 第9天的特征气味为臭鸡蛋气味、 腐烂味, 见图3(c). PCA分析显示接菌荧光假单胞菌N菌株的鱼肉贮藏3、 6和9 d产生的气味被分在了两个不同象限, 说明与无接菌鱼肉相比, 荧光假单胞菌加速了鱼肉腐败的进程.
接菌嗜冷杆菌C后0和3 d的样品特征气味与新鲜鱼肉相同, 具有咸味和腥味; 6 d样品特征气味为闷臭、 米味和生青; 第9天样品产生复杂气味, 有牛肉膏培养基气味、 菠萝气味、 腐烂气味、 酒味和臭鸡蛋气味, 以酒味和腐烂味最为突出, 见图3(d). PCA分析显示接菌嗜冷杆菌C 菌株的鱼肉贮藏期产生的气味分布在3个不同象限, 气味种类复杂而特征性气味不明显, 表明该菌株的致腐性弱, 鱼肉腐变进程缓慢.
将各菌产生的特征气味综合比较, 具体见图4~5, 分析气味特征变化的规律与致腐差异. 气味的变化趋势如箭号所示: 尽管未接菌鱼肉在贮藏过程中发生气味变化, 但相对而言, 气味变化较小, 始终趋于同一个象限; 由腐败希瓦氏菌(6 d和9 d)和荧光假单胞菌(9 d)在腐败的后期将主导气味特征, 且虽二者都能加速鱼肉腐败的进程, 但与希瓦氏菌相比较, 荧光假单胞菌致腐性较弱; 嗜冷杆菌致腐性弱, 贮藏后期产生的特征气味与另外二菌差异显著.

图3 接菌不同菌株的鱼肉样品4 ℃贮藏气味变化PCA分析图Fig.3 PCA of smell changes about fish inoculated with different strains

图4 腐败菌引起大黄鱼肉腐变气味变化趋势PCA分析Fig.4 Smell changes trends about fish inoculated with spoilage bacteria analyzed by PCA
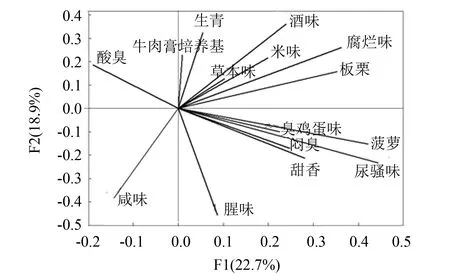
图5 各腐败菌引起大黄鱼腐变气味载荷图Fig.5 PCA loading plot of smell changes about fish inoculated with spoilage bacteria
2.2.2 气相色谱检测分析
大黄鱼鱼体的自然菌群是由多种细菌组成的混合菌群. 为明确各种细菌在致腐过程中的相互影响, 利用气相色谱(GC)分析混合菌株和单菌株接菌鱼肉样本不同时期产生的气味特征. 采用PCA法对GC数据进行分析, 判别样品鲜度和菌株的致腐能力, 见图6.
1) 希瓦氏菌K和假单胞菌N混合接菌鱼肉贮藏3 d时, 以希瓦氏菌K为主导腐败能力增强; 贮藏6 d时, 假单胞菌N占优势, 混合菌株样品与假单胞菌N样品接近; 贮藏9 d时, 混合菌株样品气味特征进入希瓦氏菌K所在象限, 致腐性以希瓦氏菌K为主导, 见图6(a).
2) 希瓦氏菌K和嗜冷杆菌C混合接菌鱼肉贮藏3 d时, 以希瓦氏菌K为主导其致腐能力稍有增强; 贮藏6 d时, 混合菌株样品与嗜冷杆菌C样品接近、 分布于同一象限, 希瓦氏菌K菌株的致腐性受嗜冷杆菌C菌株的抑制; 贮藏9 d时, 混合菌株样品气味特征进入希瓦氏菌K所在象限, 致腐性以希瓦氏菌K为主, 见图6(b).
3) 当假单胞菌N和嗜冷杆菌C混合接菌鱼肉贮藏3 d时, 致腐性强于单菌株接菌样本; 贮藏6 d和9 d时假单胞菌N菌株的致腐性受嗜冷杆菌C的抑制, 这2个时期混合菌株样品产生的气味特征接近嗜冷杆菌C贮藏第6天时气味特征, 腐变进程变缓慢, 见图6(c).
4) 3株菌混合接菌鱼肉加速鱼肉腐败进程, 贮藏3 d的鱼肉腐变水平已接近希瓦氏菌K第6天, 混合接菌鱼肉样品贮藏9 d时的气味特征进入希瓦氏菌K所在象限, 见图6(d).
综上所述, 希瓦氏菌K的致腐性最强、 并主导最终特征气味, 假单胞菌N菌株致腐性中等, 嗜冷杆菌C菌株致腐性弱. 混合菌株接菌鱼肉时, 前期有增强致腐性的趋势, 中期弱致腐菌对强致腐菌有一定的制约作用, 后期鱼肉腐败菌以强致腐菌占优势.

图6 希瓦氏菌K、 假单胞菌N与嗜冷杆菌C混合接菌时致腐互作关系GC数据主成分分析Fig.6 PCA of interaction of S. putrefaciens, P. fluorescens and P. immobilis about spoilage ability by GC analysis
3 总结与讨论
3.1 大黄鱼腐败细菌的致腐性分析
以产挥发性盐基氮和三甲胺的能力为指标, 分析比较各种腐败菌的致腐性. 检测结果表明, 腐败希瓦氏菌(S.putrefaciens)菌株A致腐性强, 表现在产挥发性盐基氮和三甲胺的能力强; 荧光假单胞菌(P.fluorescens)菌株N和腐败希瓦氏菌O和K菌株致腐性中等, 在接菌鱼肉中后期有较强的产三甲胺能力; 静止嗜冷杆菌(P.immobilis)菌株C的致腐性弱, 产挥发性盐基氮和三甲胺能力差.
利用气相色谱GC和PCA分析, 探讨混合菌群中各种细菌在致腐过程中的相互影响. 混合菌株接菌鱼肉时, 菌株间的相互作用表现为: 前期有增强致腐性的趋势, 中期弱致腐菌对强致腐菌有一定的制约作用, 后期以强致腐菌占优势. 本研究的混合菌接菌鱼肉时, 鱼肉腐变中期假单胞菌和嗜冷杆菌对腐败希瓦氏菌的致腐性有一定的制约作用, 这种制约作用可能是由于菌株间空间竞争和营养竞争造成. 据报道在假单胞菌与希瓦氏菌的混合菌群中, 当具产铁载体的假单胞菌达到108CFU·mL-1时, 腐败希瓦氏菌受抑制[7], 在假单胞菌占优势的菌相中, 腐败希瓦氏菌被检出量少[8].
3.2 GC-O用于大黄鱼鲜度评价及腐败菌种类辨别
利用气相色谱-嗅觉测量法(GC-O)检测结合PCA分析区分大黄鱼鲜度. 大黄鱼鱼肉在贮藏过程中产生的特征性气味分为三类: 第一类是鲜鱼阶段, 其气味为新鲜鱼肉的自然本体味, 咸味和腥味; 第二类是鱼肉腐败始期和扩展阶段, 产生非刺激性的气味特征, 如香甜味、 米味、 酸臭、 闷臭、 酒味、 腐烂味、 牛肉膏培养基味; 第三类是腐败终结阶段, 产生刺激性的气味特征, 如臭鸡蛋气味、 腐烂味, 另外还包含了菠萝、 草本等气味.
分析各菌气味特征变化的规律与致腐差异. 无接菌鱼肉在贮藏过程中发生气味变化较小; 腐败希瓦氏菌和荧光假单胞菌在腐败后期将主导气味特征, 二者都能加速鱼肉腐败的进程, 但与希瓦氏菌相比较荧光假单胞菌致腐性较弱; 嗜冷杆菌产生腐败性气味弱, 但能产生更为复杂的气味.
利用GC-O分析得到不同菌株对风味产生不同的贡献. 对风味的研究采用质谱或电子鼻分析, 无法得到气味的信息, 此研究体现出GC-O的特色. 采用GC-O和主成分分析, 对数据挖掘能起到很好的作用.
[1] 农业部渔业局. 中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2016.
[2] 郭全友, 杨宪时, 许钟, 等. 对新捕获与流通过程中大黄鱼新鲜度与细菌种群的研究[J]. 食品与机械, 2010, 26(4): 60-63.
[3] 张雯, 倪莉, 黄志清, 等. 冰鲜大黄鱼肠腔细菌类群鉴定及菌群结构分析[J]. 中国食品学报, 2013, 13(12): 188-196.
[4] 张雯, 倪莉, 吕旭聪, 等. 冰鲜大黄鱼鲜度评价及鲜度与细菌菌群的关系[J]. 中国食品学报, 2013, 13(7): 181-188.
[5] 郭全友, 杨宪时, 许钟, 等. 养殖大黄鱼冷藏过程中细菌菌相的变化[J]. 中国水产科学, 2007, 14(2): 301-308.
[6] 中华人民共和国水产行业标准. 水产品中挥发性盐基氮的测定: SC/T 3032-2007 [S]. 北京: 中国农业出版社, 2008.
[7] GRAM L, MELCHIORSEN J. Interaction between fish spoilage bacteriaPseudomonassp. andShewanellaputrefaciensin fish extracts and on fish tissue[J]. Journal of Applied Microbiology, 1996, 80(6): 589-595.
[8] 蓝蔚青, 谢晶, 周会, 等. 不同时期鲳鱼冷藏期间优势腐败菌的多样性变化[J]. 食品科学, 2015, 36(2): 226-231.
Spoilageabilityandinteractionsbetweenspoilagebacteriaisolatedfromlargeyellowcroakerinlowtemperature
ZHANG Wen, BIAN Dan, YANG Dongying, FANG Huiping, NI Li
(Institute of Food Science and Technology, Fujian Center of Excellence for Food Biotechnology,Fuzhou University, Fuzhou, Fujian 350116, China)
Total volatile basic nitrogen (TVBN), trimethylamine (TMA) and GC-O fingerprint were used for characterization the spoilage ability aboutS.putrefaciens,P.immobilisandP.fluorescensin low temperature and reveal the interaction between mixed bacteria. The research results showed thatS.putrefacienswas with strong volatile nitrogen production ability and dominate the putrid odor.P.fluorescenshad a similar trend of spoilage with weaker spoilage ability thanS.putrefaciens.P.immobiliswas with the weakest volatile nitrogen production ability, but it could produce more complex smell.P.fluorescensandP.immobilisinhibited the spoilage in the early storage time.
spoilage bacteria;S.putrefaciens;P.immobilis;P.fluorescens; spoilage ability;Pseudosciaenacrocea
10.7631/issn.1000-2243.2017.05.0748
1000-2243(2017)05-0748-06
S983
A
2016-04-19
张雯(1980-), 女, 博士, 副教授, 从事食品微生物方面的研究, zhangwen@fzu.edu.cn
福建省自然科学基金资助项目(2016J01156); 国家自然科学基金青年科学基金资助项目(31601532); 福州市科技局院校合作资助项目(2016-G-75)
(责任编辑: 蒋培玉)
