不同萌发温度下储藏方式和添加GA3对3种荒漠十字花科植物种子萌发的影响
2017-12-01,,·
, , ·
(1.中国科学院新疆生态与地理研究所,中国科学院干旱区生物地理与生物资源重点实验室, 新疆 乌鲁木齐 830011;2.新疆大学资源与环境科学学院,新疆大学绿洲生态教育部重点实验室, 乌鲁木齐 830046;3.乌鲁木齐职业大学旅游学院, 新疆 乌鲁木齐 830002)
不同萌发温度下储藏方式和添加GA3对3种荒漠十字花科植物种子萌发的影响
贾风勤1,2,3,张元明1,塔西甫拉提·特依拜2
(1.中国科学院新疆生态与地理研究所,中国科学院干旱区生物地理与生物资源重点实验室, 新疆 乌鲁木齐 830011;2.新疆大学资源与环境科学学院,新疆大学绿洲生态教育部重点实验室, 乌鲁木齐 830046;3.乌鲁木齐职业大学旅游学院, 新疆 乌鲁木齐 830002)
为揭示土壤种子库种子萌发对外界环境变化的响应策略,设计储藏方式和温度对古尔班通古特沙漠3种十字花科植物菥蓂、庭荠和条叶庭荠种子萌发影响。2个萌发温度(5 ℃/15 ℃、15 ℃/25 ℃)下,以室温干藏种子为对照,设置5种储藏处理(干种子:冬季室外干藏、冬季室外沙埋;吸胀种子:4 ℃、 -10 ℃、冬季室外沙埋)及添加外源GA3的萌发实验。结果表明, 1) 5 ℃/15 ℃萌发条件下,与对照相比,各储藏处理都显著延长了菥蓂和庭荠初始萌发时间,冬季室外沙埋干藏处理显著降低了庭荠种子萌发率。15 ℃/25 ℃萌发条件下,4 ℃、湿藏处理显著延长了3种种子初始萌发时间,同时显著抑制条叶庭荠种子萌发;各储藏处理下菥蓂种子初始萌发时间显著缩短,对萌发率无影响。 2) 添加GA3可使各储藏处理下种子萌发进程由停滞状态转变为持续萌发。 3) 各储藏处理和对照中菥蓂和庭荠种子的潜在萌发率均超过50%,5 ℃/15 ℃温度下大量条叶庭荠种子腐烂或失去活力导致潜在萌发率低于20%;添加GA3对3种种子潜在萌发率贡献不同。
种子库; 储藏方式; 萌发率; 外源激素
一年生植物以种子为繁殖体延续后代种群,种子散布后可能会萌发、死亡或留存在土壤种子库中[1],土壤种子库为群落演替、更新及受损生态系统恢复提供稳定的繁殖体[2-3],尤其是在环境恶劣区域[4-6]。种子萌发是一年生植物生活史的开端[7],从种子到幼苗的转变是许多植物生活史中一个高风险时期[8],因此种子萌发特性也被认为是与幼苗存活、建成以及物种适合度相关的重要功能特征[6]。
种子萌发受自身基因、生理和形态等属性影响[9-11],还与自然环境中非生物因子密切相关。在荒漠地区,能够每晚经受露水及小量雨水反复润湿的浅层或地表种子,较深埋种子萌发能力有所提高[12-13];变温环境通常更有利于多数种子的萌发[14]。也有研究发现,种子对萌发环境波动的敏感程度不同,轻度和深度休眠种子萌发状况均可能由于环境变化传递出的加强或解除休眠信号而选择萌发或继续休眠[15]。也就是说,如果种子立即萌发会遇到不良环境而导致种群适合度下降时,种子会选择推迟萌发而出现休眠,形成短暂或持久土壤种子库,成为增大物种适合度的一种生态策略。
国内许多学者开展了古尔班通古特沙漠不同生境、种子属性等对种子萌发研究,结果表明,干旱和湿润条件下,生物结皮会抑制或促进荒漠建群植物白梭梭以及短命植物涩芥、狭果鹤虱的萌发[16],倒披针叶虫实种子采取分批萌发响应沙丘不同部位水分和风蚀变化[17],小车前种子表面粘液可调节水分以减缓种子萌发速率[18],同时古尔班通古特沙漠植物种子可通过爆发型、过渡型、缓萌型和低萌型的萌发特性作为植物生存的一种风险分摊策略[19-20],这些研究丰富了古尔班通古特沙漠种子生态学和种群生态学的研究内容,对干旱区脆弱生态系统植被保护和保育提供了一定的理论依据。但是,在野外调查过程中观察到,经秋季进入冬季,分布在0~2 cm深土壤种子库表面或浅层土壤中种子,可能由于秋季降雨较多而使呈吸胀状态种子被埋藏在零度以下季节性冻土中长达2~3 个月;也可能存在由于秋季降水少,种子始终暴露或浅埋在干燥土壤环境中依次经历低温、冷冻的温度变化过程。春季积雪消融引起的冻融循环过程也增加了种子库中种子遭遇冷冻事件的几率,同时积雪融水也可能使种子在长达2周时间内处于水淹状态。前期观察发现,将自然风干种子放置于室温或冰箱(4 ℃或 -10 ℃)中储藏一定时间进行萌发实验与种子散布至萌发所经历的环境条件相差甚远,也影响着实验结果,使研究者或管理者无法了解该区域植物种子经历冬季低温后的萌发情况和物种对生境的适应性。
十字花科植物是古尔班通古特沙漠植被的重要组成部分[21],从降水丰富的沙漠南缘到降水逐渐稀少的沙漠北部都有十字花科植物分布,生长在活动沙丘顶部、结皮丰富的丘间低地和沙丘坡部等地[22-23]。本实验以分布在3种生境内的十字花科植物为研究材料:菥蓂(Thlaspiarvense),分布在沙漠边缘绿洲路边、田旁等;庭荠(Alyssumdesertorum),分布在沙漠-人工绿洲交错带;条叶庭荠(Alyssumlinifolium),分布在沙漠中,通常与尖喙牻牛儿苗共同成为为沙漠短命植物层片的优势种,通过模拟种子库可能遭遇的环境种子设置湿冷层积、室外干藏、室温干藏等储藏方式研究种子萌发与休眠变化情况,以期弄清古尔班通古特沙漠环境因素波动(温度、土壤水分、埋深)对种子库种子萌发的影响,由于所研究植物位于古尔班通古特沙漠这一特殊区域,本研究根据沙漠降水、种子春萌和秋萌期气温设置了饱和水储藏和更低层积温度以及与外界气温变幅更加接近的萌发变温处理,以探明3种植物种子对不同储藏处理的萌发响应特征,旨在通过沙漠植物种子萌发与休眠特性认识种子自身生物学与生态学特性,并通过掌握沙漠植物从潜在种群变为现实种群条件认识植物的更新机制。
1 实验材料与方法
1.1 实验材料
分布于不同生境的3种植物菥蓂、庭荠和条叶庭荠种子采自古尔班通古特沙漠。2015年6月所选取植株间距大于3 m,种子成熟尚未脱落或少量脱落时进行采集。将采集的种子自然风干后,清理干净,放置于实验室(平均温度为23 ℃)环境中保存备用。实验前对3种十字花科植物供试种子采用TTC法进行活力快速测定,经检测种子活力均为90%以上。
1.2 种子储藏处理及萌发实验
1.2.1 种子储藏方式
大约2 000粒/种植物种子被分别装入由滤纸做成的种子袋中(5 cm×5 cm)。这些信封分别被放在5种储藏处理环境中(表1)。沙埋实验中,种子袋依次被水平地放置在2 cm深含饱和水细沙(种子呈吸胀状态)和干燥细沙中,室内放置24 h后,统一移入冬季室外自然环境;水埋实验中,将种子袋水平放置在盛有15 mL蒸馏水的9 cm培养皿中保证种子呈吸胀状态,每皿1袋。所有培养皿均用棕色自封袋包裹,24 h后分别放置于4 ℃和-10 ℃冰箱中;干藏处理及对照处理(干藏)实验中,种子袋被放置在尼龙网袋(25 cm×25 cm)中,分别置于冬季室外自然环境和实验室环境。每一储藏处理放置6个种子袋。储藏处理实验开始于2015年12月7日,2个月后(2016年2月7日)一次性将所有种子袋从各处理环境中取出,共计36个,进行萌发实验。
表1 储藏实验设计概述

编号储藏处理处理设置种子状态萌发温度(℃)ck室内冷藏处理无沙埋自然干燥5/1515/25A冬季室外冷藏处理无沙埋自然干燥5/1515/25B冬季室外冷藏处理种子埋深2cm自然干燥5/1515/25C4℃,冰箱冷藏处理无沙埋种子吸胀5/1515/25D-10℃,冰箱冷冻处理无沙埋种子吸胀5/1515/25E冬季室外冷冻处理种子埋深2cm种子吸胀5/1515/25
1.2.2 种子萌发实验
在2016年2月8日将所有处理种子移入实验室平衡24 h后,于2月9日开始实验,4月28日结束。将供试种子分别播种于铺有2层滤纸的培养皿(直径9 cm)中,每一培养皿放入50粒种子,加入10 mL蒸馏水。设置5 ℃/15 ℃、15 ℃/25 ℃ 2个萌发温度,每个温度每个储藏处理3次重复,以实验室自然环境下干藏为对照。所有供试种子均置于SPX-150 B-Z型培养箱内,高温12 h(光照),低温12 h(黑暗)进行培养。
1.2.3 GA3添加实验
通过预实验观察到庭荠和条叶庭荠(均存在休眠)在4 ℃湿藏1个月和2个月后均未破除休眠,同时部分储藏处理下菥蓂种子也出现休眠。因此,针对萌发过程中出现萌发停滞现象的种子设计添加100 mg/L 赤霉素(GA3)[24]环节,以进一步观察外界环境因素变化对种子萌发的影响。GA3添加时间为2016年3月2日,即从2016年2月20日起至3月1日观察到除15 ℃/25 ℃萌发温度下的菥蓂种子外,其余各处理下种子一直呈萌发停滞状态。GA3添加量为10 mL/皿,在实验进行过程中及时补充GA3溶液。
1.3 萌发指标测定
种子萌发实验中,每天统计1次萌发情况,以胚根突破种皮2 mm为萌发标准,萌发实验以连续14 d无种子萌发视为萌发结束。统计种子初始萌发天数、萌发率。其中:初始萌发时间为每个重复中第1粒种子萌发所需时间。
萌发率(%)=萌发种子数(添加GA3前 + 添加GA3后)/供试种子总数×100%
…… (1)
萌发结束后,统计未萌发种子中腐烂种子数。对未萌发但完好种子采用TTC法染色,观察24 h后胚着色情况,如果胚为红色,则为有活力种子且具潜在萌发能力种子。
休眠率(%)=染色种子数/供试种子数×100%
……(2)
潜在萌发率(%)=萌发率+休眠率
……(3)
1.4 数据分析方法
利用SPSS 17.0软件对实验数据进行统计分析。对同一温度下不同储藏处理,同一温度、同一储藏处理条件下添加GA3后3种十字花科植物种子萌发率、初始萌发时间等指标采用单因素方差分析(One-way ANOVA),差异显著性采用Tukey方法进行多重比较。每一指标的图或表均为3次重复的平均值,利用Graphpad Prism 5.0绘图。
2 结果与分析
2.1 种子初始萌发时间和萌发率的比较
与对照相比,5 ℃/15 ℃萌发温度下的5种储藏方式均表现为显著延长了菥蓂和庭荠初始萌发时间,初始萌发时间延后最长分别可达27 d和23 d,条叶庭荠初始萌发时间无显著影响。15 ℃/25 ℃萌发温度下,4 ℃湿藏显著抑制庭荠和条叶庭荠萌发,初始萌发时间分别延后27 d和32 d。同一储藏处理下,15 ℃/25 ℃变温条件显著缩短室温干藏、冬季室外干藏、冬季室外沙埋干藏和冬季室外沙埋湿藏下菥蓂初始萌发时间,缩短时间分别为3,34,33 d和31.3 d;高温萌发也显著缩短室温干藏、-10 ℃湿藏和冬季室外沙埋湿藏条叶庭荠初始萌发时间。
表2 不同温度和储藏方式处理3种植物种子萌发特征

植物种指标培养温度(℃)储藏方式ckABCDE菥蓂初始萌发时间(d)5/159.0c/A37.0a/A35.0a/A36.0a/A19.6bc/A33.3ab/A15/256.0ab/B3.0b/B2.0b/B23.0a/A2.0b/B2.0b/B萌发率(%)5/15100.091.377.491.296.596.715/25100.0100.0100.0100.0100.0100.0庭荠初始萌发时间(d)5/159.0b/A5.0c5.3c32.0a5.0c5.0c15/255.0b/B2.0b10.3b29.0a2.0b2.0b萌发率(%)5/1593.5a/A83.8ab38.4b/B59.7ab58.6ab48.2ab15/2576.1B72.668.76A54.879.263.1条叶庭荠初始萌发时间(d)5/1530.7A23.023.323.322.0A23.3A15/255.0b/B17.3ab2.7b37.0a2.3b/B2.0b/B萌发率(%)5/1520.0B5.2B7.75.15.6B6.9B15/2566.5b/A96.3a/A49.4bc17.4c42.2bc/A31.2bc/A
注:斜杠前不同小写字母表示储藏方式处理之间差异显著(plt; 0.05),斜杠后不同大写字母表示温度处理之间差异显著(plt; 0.05)。下同。
5 ℃/15 ℃萌发时,相对于对照,冬季室外沙埋干藏处理庭荠种子萌发率显著降低,较对照下降了55.1%;5种储藏处理下菥蓂和条叶庭荠种子萌发率与对照无差异。15 ℃/25 ℃条件下,4 ℃湿藏环境显著抑制条叶庭荠种子萌发,总萌发率仅为对照的26.1%;但冬季室外干藏显著促进其种子萌发,是对照的1.45倍。同一储藏条件下,相对于5 ℃/15 ℃萌发温度,15 ℃/25 ℃变温条件促进室温干藏、冬季室外干藏、-10 ℃湿藏和冬季室外沙埋湿藏条叶庭荠种子总萌发率显著提高。该温度条件也显著促进冬季室外沙埋干藏环境下庭荠种子萌发,但对室温干藏的种子则表现为显著抑制。
2.2 种子萌发进程的比较
在添加GA3前,5 ℃/15 ℃萌发条件下,所有储藏处理抑制了3种植物种子萌发,萌发率均低于50%且萌发7 d后基本呈停滞状态(图1)。菥蓂种子:除了对照处理下在第9天有种子萌发外,5种储藏处理的种子均无萌发,直至第19.6天湿藏-10 ℃处理下种子开始萌发。庭荠种子:初始萌发时间最早为第5天,分别为冬季室外干藏、-10 ℃湿藏和冬季室外沙埋湿藏的种子,但自第5天后除冬季室外干藏种子有持续萌发外,其余储藏处理下均无种子萌发。同时,4 ℃湿藏种子始终无萌发。条叶庭荠种子:在近22 d的萌发期内,5种储藏处理下种子萌发呈停滞状态且最高萌发率仅为2.27%。表明5 ℃/15 ℃变温环境不适宜3种植物种子萌发且5种储藏处理后种子呈休眠状态(表2,图1)。对于15 ℃/25 ℃条件下的3种种子,随萌发时间延长,5种储藏处理下3种植物种子萌发进程有明显差别。菥蓂种子:除4 ℃湿藏处理种子呈休眠状态外,其余4种处理下种子萌发进程较对照表现为缩短且均呈现出爆发式萌发,萌发率达100%的时间平均为6 d。庭荠种子:冬季室外干藏、-10 ℃湿藏和冬季室外沙埋湿藏种子在萌发期内萌发率均高于对照,尤其是冬季室外干藏处理种子萌发率达87.1%,但总体也表现为萌发7 d后呈萌发停滞现象。条叶庭荠种子:随萌发时间延长,除4 ℃湿藏处理外,其余4种储藏处理下萌发率均表现逐渐上升,最高萌发率也仅为13.3%,萌发7 d后种子萌发呈停滞状态。
于2016年3月2日对萌发停滞(平均时间为19 d,图中虚线处))的所有处理种子添加100 mg/L GA3溶液10 mL/皿。由图1可知,添加GA3后,5 ℃/15 ℃条件下,菥蓂种子:对照及5种储藏处理下均表现出种子休眠破除和萌发进程后期的爆发式萌发态势。庭荠种子:对照、冬季室外干藏、4 ℃湿藏和-10 ℃湿藏处理均能有效破除种子休眠且表现为阶段性爆发式萌发特征。条叶庭荠种子:对照及5种储藏处理下均呈休眠状态。在整个萌发进程中,5种储藏处理下3种植物种子萌发率均低于对照(图1)。15 ℃/25 ℃条件下,4 ℃湿藏处理的菥蓂种子休眠被破除且萌发率达100%。庭荠种子:除冬季室外干藏处理外,其余储藏处理下的休眠种子休眠被破除且均呈现缓慢萌发趋势,GA3对冬季室外干藏处理种子萌发无影响。条叶庭荠种子:室外干藏处理能有效破除种子休眠,相对对照而言其萌发过程表现为快速均匀上升的特征(图1)。
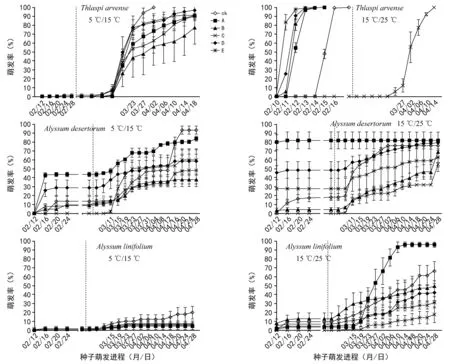
图1 不同温度和储藏方式处理3种植物种子萌发进程(虚线处为添加GA3的时间)
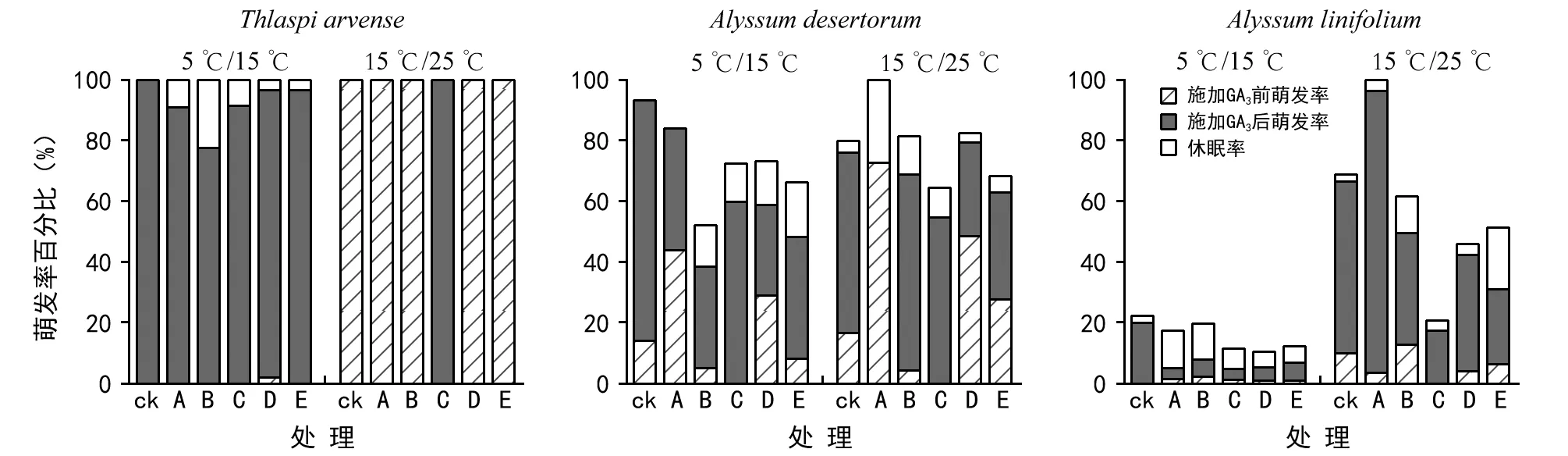
图2 2类萌发率在潜在萌发率中的分布
2.3 种子潜在萌发率的比较
5 ℃/15 ℃下,所有储藏处理和对照处理菥蓂和庭荠种子的潜在萌发率均超过50%,最高达100%;条叶庭荠由于大量种子腐烂或失去活力,潜在萌发率低于25%。
15 ℃/25 ℃萌发条件下,菥蓂种子:各储藏处理下潜在萌发率均达100%;庭荠种子: 不同储藏处理萌发率均可达65%以上,并且冬季室外干藏处理最大预计萌发率为100%,表明冬季室外干藏处理有利于庭荠种子萌发;条叶庭荠种子: 4 ℃、湿藏和- 10 ℃、湿藏分别使53.9%和79.2%的种子失去活力或腐烂,导致潜在发率低于50%(图2)。
3 讨 论
温度是影响种子萌发的重要因素之一,可通过直接影响种子萌发或间接调节种子休眠而决定种子萌发行为。崔现亮等研究表明,5 ℃/25 ℃变温下12种灌木种子平均萌发率最高[25],10 ℃/20 ℃下桔梗种子能够良好萌发[26],15 ℃/25 ℃下碟果虫实能够实现最佳萌发[27],植物这种对环境信号完善的感知体系是其对不可预测的环境自然进化的结果[28]。本研究中,15 ℃/25 ℃能满足除4 ℃,湿藏处理外其他储藏处理下菥蓂种子萌发,且其萌发过程可集中在短短数天完成,呈爆发式萌发即速萌型。因为在古尔班通古特沙漠南缘绿洲,4月平均气温为5 ℃/15 ℃,5月平均气温为10~25 ℃,意味着菥蓂这类非早春短命一年生植物在长期进化过程中发展出适应绿洲5月后平稳气候环境的萌发策略,此策略可确保植物完成生活史,并增加其竞争优势[29]。这与刘会良等[19]对古尔班通古特沙漠及其周缘地区52种植物萌发策略研究发现爆发型植物主要分布在外界环境较好且稳定区域的结果相同。庭荠所处的沙漠与绿洲交错带气候环境因子变化较绿洲为不稳定,使植物面对高温、干旱和霜冻等胁迫风险增加,其萌发策略为过渡萌发型,遇到有适宜萌发环境时呈高萌发,可以推测在4—6月短命植物生长季内都可能会有庭荠种子萌发。条叶庭荠多生长在沙漠腹地,生存环境条件恶劣且多变,种子采取谨慎萌发策略即渐进扩展式萌发。这种萌发策略也被生长在其他环境条件多变的植物所采用[30-31],这意味着即便存在适宜萌发环境条件依旧会有较多数量种子保持休眠状态,在野外则可保证不利条件下种子在土壤中形成短暂或持久种子库以减少种群灭绝的概率。
对于不同植物种子通过改变储藏方式使种子选择萌发或继续休眠[6]。非冷冻环境下湿藏(0~5 ℃)通常能有效打破种子休眠,这种现象在一些冻土地带植物[32-33]和许多温带植物[34]中都被观察到,但也有研究表明,低温湿藏会降低种子萌发能力[35-36]。在本研究中,4 ℃湿藏处理几乎完全抑制了2个萌发温度下3种十字花科植物种子萌发,这与Shimono和Kudo[37]的研究结果相似。由此可推断由于荒漠植物常年处于干旱、少雨环境中,对水淹环境不耐受。在环境不适宜时(如土壤水分的极大波动),种子进入休眠,采取保持活力留存在土壤种子库中的风险分担策略[38]。对于干旱地区植物种干藏也能打破种子休眠[25],如本研究中5 ℃/15 ℃冬季室外干藏条件下庭荠种子萌发率超过80%,也许是该储藏处理经历了自然条件下冬季低温到春季高温的变温过程,逐渐打破了庭荠种子休眠。Qin J H et al对色木槭(Acermono)种子萌发研究表明,埋藏在5~10 cm冻土层种子萌发率显著高于非冻土层[39]。本研究中,2种冷冻储藏处理未影响菥蓂种子萌发,提高了庭荠萌发率,降低了条叶庭荠萌发率。对庭荠属2物种而言,可能是恒定温度的冷藏不能有效解除种子休眠,这与申建红等[40]研究结论相一致。值得关注的是条叶庭荠种子在冷冻处理后有大量种子失活或腐烂。
为了探索影响储藏处理下种子萌发率低的内在原因,对处于萌发停滞状态的种子进行了添加外源GA3处理,结果表明GA3能破除菥蓂种子休眠,有效减轻庭荠种子休眠,并使条叶庭荠种子休眠得到一定程度释放,这与Yao 等[41]的研究结果相一致。GA3通过促进种子储藏物质转化,增强胚活性对萌发产生影响[10]。GA3还具弱化胚外围结构的作用,促进种皮休眠类种子萌发[42]。在本研究中观察到凡是未萌发菥蓂种子种皮均呈黑色且种子在水中无正常的吸胀反应,添加GA3后种皮呈棕红色且呈爆发式GA3萌发,由此可推断15 ℃/25 ℃萌发时,4 ℃湿藏菥蓂种子休眠的原因是种皮限制种子吸胀所引起。同样,15 ℃/25 ℃时各储藏处理和对照处理下条叶庭荠萌发率和具活力种子数量都有显著提高,推测条叶庭荠种子休眠的原因可能是胚尚未完成后熟所致。
Montiel 和Mintaňa 认为,萌发实验中所得种子萌发率应当为最小萌发率,因为不知道那些没有萌发的种子是活力丧失还是处于休眠状态[43],有活力种子对可能的(潜在的)萌发和更新植物组成土壤种子库是有效的[44-45]。本研究通过添加GA3和种子活力检测实验也证明了这个观点。以4 ℃、湿藏菥蓂种子为例,2个温度条件下,未添加GA3前种子萌发率均为0,未萌发种子受到GA3刺激后,种子萌发率达100%。这表明储藏处理造成的种子无萌发,只是使种子进入休眠状态并保持活力。由此可推断如果适宜种子萌发的时间足够长,几乎所有具活力种子都有可能萌发。
不少荒漠植物种类具粘液种子,如十字花科、车前科植物等,粘液可粘附沙粒或土壤造成种子体积和重量增加,影响着种子散布萌发[46]及在土壤种子库空间分布特征[47],它们对荒漠环境的这些特殊适应机制在荒漠植被构成与演替中具有重要作用。伍晨曦等对古尔班通古特沙漠短命植物小车前种子萌发研究表明,表面粘液通过控制种子吸收水分来调控种子萌发,或抑制或促进[17]。在本研究中,15 ℃/25 ℃条件下庭荠种子一直被大量粘液包被,但冬季干藏非沙埋和-10 ℃、湿藏种子均较对照表现出初始萌发时间早和萌发率高的特征。同时冬季干藏处理下条叶庭荠种子萌发时粘液不明显,但萌发率最高。因此推测储藏方式可能会影响粘液分泌进而影响萌发。值得关注的是,在5 ℃/15 ℃萌发温度下,所有储藏处理下条叶庭荠种子失活和死亡情况严重,占比为77.7%~89.5%,原因还需进一步研究。
综上所述,菥蓂、庭荠、条叶庭荠种子在不同处理下采取延迟萌发并形成短暂或持久种子库策略响应不利环境条件,同时采取爆发式萌发、过渡型萌发和谨慎型萌发响应有利环境条件,显示出植物种子萌发对策与栖息地生境具有高度的生态适应性,这也是植物对其生存环境长期适应的结果。
[1]Soriano D,Huante P,Ganboa-deBuen A,et al.Effects of burial and storage on germination and seed reserves of 18 tree species in a tropical deciduous forest in Mexico[J].Oecologia,2014,174:33-44.
[2]Bai W M,Bao X M,Li L H.Effects ofAgtiophyllumsquarrosumseedbanks on its colonization in a moving sand dune in Hunshandake Sand Land of China[J].2004,59:151-157.
[3]Bienau M J,Hattermann D,Krǒncke M,et al.Snow cover consistently affects growth and reproduction ofEmpetrumhermaphroditumacross latitudinal and local climatic gradients[J].Alp Botany,2014,124:115-129.
[4]Dreber N,Esler K J.Spatio-tempotal variation in soil seed banks under contrasting grazing regimes following low and high seasonal rainfall in arid Namibia[J].J Arid Environ,2011,75:174-184.
[5]Ma M J,Zhou X Y,Du G Z.Soil seed bank dynamics in alpine wetland succession on the Tibetan Plateau[J].Plant Soil,2011,346:19-28.
[6]Wang J H,Chen W,Ma R J,et al.Role of short-term cold stratification on seed dormancy break and germination of alien species in southeastern China[J].Plant Ecology,2016,217:383-392.
[7]李雪华,蒋德明,刘志民,等.温带半干旱地区一年生植物种子的萌发特征[J].生态学报,2006,26(4):1 194-1 199.
[8]Harper J.Population biology of plant[M].Academic Press,New York,1977.
[9]Wang L,Huang Z Y,Baskin J M.Germination of dimorphic seeds of the desert annual halophtyeSuaedaaralocaspica(Chenopodiaceae),a C 4 plant without Kranz anatomy[J].Annals of Botany,2008,102:757-769.
[10]Nowag A.Management of seed dormancy inFagussylvatica,FraxinusexcelsiorandPrunusavium.Combined Proceedings of International Plant[J].Propagators’ Society,1998,48:192-199.
[11]Doody C N,O’Reilly C.Effect of long-phase stratification treatments on seed germination in ash[J].Annals of Forest Science,2011,1:139-147.
[12]Long R L,Kranner I,Panetta F D,et al.Wet-dry cycling extends seed persistence by reinstating antioxidant capacity[J].Plant Soil,2011,228:511-519.
[13]González-Zeruche L,Vazquez-Yanes C,Gamboa A,et al.Natural priming ofWigandiaurensseeds during burial:effects on germination,growth and protein expression[J].Seed Science Research,2001,1:27-34.
[14]Liu K,Baskin J M,Baskin C C,et al.Effect of diurnal fluctuating versus constant temperatures on germination of 445 species from the eastern Tibet Plateau[J].Plos One,2013,8:e69364.
[15]Martinez-Ghersa M A,Ghersa C M,Benech-Arnold R L,et al.Adaptive traits regulating dormancy and germination of invasive species[J].Plant Species Biology,2000,15(2):127-137.
[16]聂华丽,张元明,吴楠,等.生物结皮对5种不同形态的荒漠植物种子萌发的影响[J].植物生态学报,2009,33(1):161-170.
[17]陈正霞,刘彤,刘尊驰,等.沙生植物倒披针叶虫实在古尔班通古特沙漠的分布[J].生态学报,2016,36(13):4 064-4 073.
[18]伍晨曦,孙羽,冯固.小车前(PlantagominutaPall.)种子表面粘液物种的吸水特性及其对干旱环境中萌发的影响[J].生态学报,2009,29(4):1 849-1 858.
[19]刘会良,宋明方,段士民,等.古尔班通古特沙漠及周缘52种植物种子的萌发特性与生态意义[J].生态学报,2011,31(15):4 308-4 317.
[20]Liu H L,Shi X, Wang J C,et al.Effects of sand burial, soil water content and distribution pattern of seeds in sand on seed germination and seedling survival ofEremospartonsongoricum(Fabaceae),a rare species inhabiting the moving sand dunes of the Gurbantunggut desert of China[J]. Plant Soil,2011,345:69-87.
[21]Qian Y B,Wu Z N,Wang Z C,et al.Relationship of spatial heterogeneity for vegetation and aeoliansand soil properties on longitudinal dunes in Gurbantunggut Desert[J].China Environment Earth Science,2013,69:2 027-2 036.
[22]Song Y Y,Zhou C B,Zhang W H.Vegetation coverage,species richness,and dune stability in the southern part of Gurgantunggut Desert[J].Ecology Research.2011,26:79-86.
[23]Fan L L,Li Y,Tang L S,et al.Combined effects of snow depth and nitrogen addition on ephemeral growth at the southern edge of the Gurbantunggut desert,China[J].Journal of Arid Land,2013,5(4):500-510.
[24]黄代红,赵娟,袁军文,等.室外人工种植抱茎叶独行菜种子破除休眠的初步研究[J].新疆农业科学,2011,48(3):480-486.
[25]崔现亮,罗娅婷,毕廷菊,等.储藏和萌发温度对青藏高原东缘12种灌木种子萌发的影响[J].生态学杂志,2014,33(1):23-32.
[26]刘有军,刘世增,纪永福,等.碟果虫实种子萌发对策及生态适应性[J].生态学报,2010,30(24):6 910-6 918.
[27]刘自刚,沈冰,张雁.桔梗种子萌发对低温、干旱及互作胁迫的响应[J].生态学报,2013,33(8):2 615-2 622.
[28]García-Fayos P,Bochet E,Cerdà A.Seed removal susceptibility through soil erosion shapes vegetation composition[J].Plant Soil,2010,334:289-297.
[29]Li X H,Li X L,Jiang D M,et al.Germination strategies and patterns of annual species in the temperate semiarid region of China[J].Arid Land Research and Management.2006,20(3):195-207.
[30]Venable D L,Lawlor L.Delayed germination and dispersal in desert annuals:escape in space and time[J].Oecologia,1980,46:272-282.
[31]Mandák B.Germination requirements of invasive and non-invasiveAtriplexspecies:A comparative study[J].Flora-Morphology, Distribution,Functional Ecology of Plants,2003,198:45-54.
[32]Chambers J C,MacMahon J A,Brown R W.Germination characteristics of alpine grasses and forbs:a comparison of early and late seral dominants with reclamation potential[J].Reclamation and Revegetation Research,1987,6:235-249.
[33]Densmore R D.Effect of day length on germination of seeds collected in Alaska[J].American Journal of Botany,1997,84:274-278.
[34]Probert R J.The role of temperature in the regulation of seed dormancy and germination.In:Fenner M (eds) Seeds:the ecology of germination in plant communities[M].CAB International,Oxon:261-292.
[35]Schütz W,Milberg P.Seed dormancy in Carex canescens:regional differences and ecological consequences[J].Oikos,1997,78:420-428.
[36]Milberg P,Andersson L.Does cold stratification level out differences in seed germinability between populations[J].Plant Ecology,1998,134:225-234.
[37]Shimono Y, Kudo G. Comparisons of germination traits of alpine plants between fellfield and snowed habitats[J].Ecological Research, 2005, 20:189-197.
[38]赵惠新,李群,周晶,等.短命植物独行菜种子萌发过程对低温的耐受特性[J].云南植物研究, 2010, 32(5):448-454.
[39]Jihong Qin, Qing Liu. Oxidative metabolism-related changes during germination of mono maple (AcermonoMaxim.) seeds under seasonal frozen soil[J].Ecological Research, 2010, 25:337-345.
[40]申建红,曾波,施美芬,等.储藏方式和时间对三峡水库消落区一年生植物种子萌发的影响[J].生态学报,2010,30(33):6 571-6 580.
[41]Yao S X,Lan H Y,Zhang F C.Variation of seed heteromorphism inChenopodiumalbumand the effect of salinity stress on the descendants[J].Annals of Botany,2010,105:1 015-1 025.
[42]Bassel G W,Zielinska E,Mullen R T,et al.Down-regulation of DELLA Genes is not essential for germination of tomato,soybean,and Arabidopsis seeds[J].Plant Physiology,2004,136:2782-2789.
[43]Montiel S,Mintaňa C.Seed bank dynamics of the desert cactus Opuntia rastrera in two habitats from the Chihuahuan Desert[J].Plant Ecology,2003,166:241-248.
[44]Baskin C C,Baskin J M.Seeds.ecology,biogeography,and evolution of dormancy and Germination[M].Academic Press,San Diego,1998.
[45]Mandujano M C,Golubov J,Mintaňa C.Dormancy and endozoochorous dispersal ofOpuntiarastreraseeds in the southern Chihuahuan Desert[J].Journal of Arid Environment,1997,36:259-266.
[46]Western T L.The sticky tale of seed coat mucilage:production,genetics,and role in seed germination and dispersal[J].Seed Science Research,2012,22:1-25.
[47]Chambers J C.Seed movements and seedling fates in disturbed sagebrush steppe ecosystems:implications for restoration[J].Ecological Applications,2000,10(5):1 400-1 413.
Effect of Storage Condition and Addition of GA3on Three Brassicaceae Species Seed Germination Under Different Germination Temperature
JIAFengqin1,2,3,ZHANGYuanming2,Tiyip·Tashpolat2
(1.Key Laboratory of Oasis Ecology Ministry of Education,College of Resources and Environment Sciences,Xinjiang University,Urumqi 830046,China;2.Key Laboratory of Biogeography and Bioresources in Arid Land,Xinjiang Institute of Ecology and Geography,Chinese Academy of Sciences,Urumqi 830011,China;3.Tourism College,Urumqi Vocational University,Urumqi Xinjiang 830002,China)
In order to understand the responses of seedlings emerging from the soil seed bank to outside environmental conditions, we studied effects of storage condition and temperature on germination of three Brassicaceae species in Gurbantunggut desert, includingThlaspiarvense,AlyssumdesertorumandAlyssumlinifolium.Taking dry seeds stored at room temperature as control, five different storage conditions were applied in our study,including (a)dry seeds stored outside in winter;(b)dry seeds buried in soil in winter;(c) imbibed seeds stored at 4 ℃;(d)imbibed seeds stored at -10 ℃; and (e)imbibed seeds buried in soil in winter. After storage,seeds were incubated at 5 ℃/15 ℃ or 15 ℃/25 ℃ for germination.Effect of exogenous GA3on seed germination was also studied. Our results showed that (1) under 5 ℃/15 ℃ regime,comparing to the control, each storage treatment significantly increased initiatory time of seed germination forTh.ArvenseandA.desertorum,germination percentage of the dry seeds buried in soil in winter forA.desertorumwas significantly reduced; initial germination time and germination percentage ofTh.arvense,A.desertorumandA.linifoliumwere shortened and increased at 15 ℃/25 ℃,respectively.In addition,under 15 ℃/25 ℃ regime,initial germination time and germination percentage of imbibed seeds stored at 4 ℃ forA.linifoliumwere significantly prolonged and inhibited, respectively. (2) Exogenous GA3facilitated consistent germination of stored seeds.(3)Germination potential of.Th.ArvenseandA.desertorumexceeded 50% under all storage condition. At 5 ℃/15 ℃, germination potential ofA.linifoliumwas lower than 20%,due to the fact that many seeds decayed or lost viability under this temperature regime.The contribution of exogenous GA3to potential germination varied among these 3 species.
soil seed bank; storage condition; germination rate; exogenous hormone
2016-11-13
科技部全球变化重大科学研究计划(973项目)(2014 CB 954200)和新疆自然科学基金(2015211 C 292)资助。
贾风勤(1973—),女,在读博士研究生,主要从事植物种群生态方面的研究;E-mail:jfqanan@sina.com。
张元明,E-mail:zhangym@ms.xjb.ac.cn。
10.16590/j.cnki.1001-4705.2017.01.007
S 330.2
A
1001-4705(2017)01-0007-08
