两种辣椒雄性不育系主要农艺性状对比分析
2017-11-10黄巍缪武
黄 巍 缪 武
(1.湖南省蔬菜研究所, 长沙 410125; 2. 湖南兴蔬种业有限公司, 长沙 410125;3. 湖南湘研种业有限公司, 长沙 410100)
两种辣椒雄性不育系主要农艺性状对比分析
黄 巍1,2缪 武3
(1.湖南省蔬菜研究所, 长沙 410125; 2. 湖南兴蔬种业有限公司, 长沙 410125;3. 湖南湘研种业有限公司, 长沙 410100)
为了解两种辣椒雄性不育系的农艺性状,对细胞核雄性不育系G06-77-2A、细胞核质互作雄性不育系C06-77-2A及两个不育系的可育系G06-77-2B的株高、开展度、花蕾、雄蕊、雌蕊等方面进行对比分析。结果表明,(1)不育系除在株高和开展度较可育系大之外,其他性状和可育系基本一样;(2)雄性不育系的花蕾及雄蕊干质量都小于其相应的可育系,而雌蕊有异常膨大的现象,这一规律细胞核质互作不育系较细胞核不育系更突出,果实没有显著性差异。可见,辣椒细胞核质互作不育系C06-77-2A及细胞核雄性不育系G06-77-2A的整体性状好,个体间表现基本一致,能够运用于不育性状的回交转育亲本或选配杂交一代亲本。
辣椒;CMS;GMS;农艺性状;对比
辣椒属常异花授粉植物,其杂交优势在抗性、产量及商品性等方面表现明显。杂交种子生产目前主要是靠人工去雄杂交制种,但生产成本高,纯度难以保障。雄性不育技术利用是解决该问题的有效途径之一[1]。
自 1951年 Martin和 Grawford、1958年Peterson先后在辣椒中报道细胞核雄性不育和细胞核质互作雄性不育以来,人们已经发现了数十个细胞核雄性不育突变体及多个细胞核质互作雄性不育源[2]。到目前为止,利用不同类型的雄性不育源已选育出多个辣椒细胞核雄性不育系,并成功实现了细胞核质互作不育类型的“三系配套”。
植物的雄性不育特性使雄性不育植株各器官形态发生变化,其中雄蕊、雌蕊、花药等器官形态有较大变化。本试验研究两种雄性不育系及其可育系植株形态、花器官及果实性状发育,从中寻找规律,为其研究提供参考。
1 试验材料
1.1 试验材料及来源
高辣椒红色素细胞核雄性不育系G06-77-2A、细胞核质互作雄性不育系C06-77-2A及两个不育系的可育系G06-77-2B (G06-77-2B既是细胞核雄性不育系G06-77-2A的可育株,也是细胞核质互作雄性不育系C06-77-2A的保持系),这些材料农艺性状相似,育性稳定,其育性的遗传分离规律清楚。试验所用3个辣椒品系由湖南湘研种业有限公司辣椒课题组选育并提供(表1)。

表1 供试材料类型与来源
1.2 两种类型辣椒雄性不育系的主要特征特性
1.2.1 细胞核质互作雄性不育系C06-77-2A
细胞核质互作雄性不育系C06-77-2A是以9704A为母本,高辣椒红色素高代自交系R06-77-2为父本杂交后,经多代回交、定向选择而育成。C06-77-2A第一次分枝节位11~12节,全生育期120 d左右。C06-77-2A不育株率和不育度均达到100%。不育系花器显示为花药退化,干瘪、瘦小及部分开裂或不开裂,其花朵大小及花瓣开张度与保持系基本一致。碘化反应显示:雄性不育系花粉粒畸形解体,内含物稀少,碘化反应显示不着色或着色浅,保持系花粉粒圆形,内含物充实,碘化反应显蓝色。不育系隔离自交不结实或单性结实成畸形果,果腔内无子可育株与不育株在F2呈3∶1分离。
1.2.2 细胞核雄性不育系G06-77-2AB
辣椒细胞核雄性不育系G06-77-2AB是采用远源杂交后代中发现的不育株,套袋回交并且连续选择而育成的隐性核不育系。其育性受1对纯合隐性基因控制,1对基因由隐性纯合状态变为杂合状态,育性就由不育变为可育。因此,G06-77-2AB两用系中的可育株生成可育和不育两种配子,而不育株只生成不育配子。在大多数状况下,不育×可育或可育株自交后代的分离比例分别是1∶1和3∶1。其中的G06-77-2B可育株植株高矮、雄蕊长短、花瓣大小与高代自交系R06-77-2常规辣椒基本相同;而不育株植株较高、分枝较多、花蕾瘦长、完全无花粉;其余农艺性状与G06-77-2B完全相同。为方便叙述,本文中将G06-77-2AB中分离出的不育株称G06-77-2A(不育系),可育株称G06-77-2B (可育系)。
2 试验方法
2.1 田间栽培与管理
供试材料于2010年2月初播种在湖南农业大学试验田,采用条播方式,每667 m2施氮、磷、钾含量均为16%的俄罗斯复合肥60 kg,菜饼50 kg,磷肥50 kg及钾肥10 kg作底肥。4月20日定植,定苗期6~7片真叶,定苗7 d后每667 m2追施氮、磷、钾含量各为16%的俄罗斯复合肥5 kg。病虫害防治及田间管理同一般大田栽培一样。各材料从6月5日至25日陆续现蕾,6月5日至10日为初花。
2.2 试验材料分级
参考袁志勤[3]、何长征[4]的方法根据花蕾形态特征、大小与花粉发育时期相互关系将花蕾分为7级,见表2。
从6月初开始,各材料相继开花。始花期,从各材料不育系及其相应可育系中任取5株典型株,摘取主花序及以上部2个分枝花蕾。根据观察,确定花蕾长度与发育时期的相应关系,同时摘取各对细胞核质互作不育系C06-77-2A、细胞核不育系G06-77-2A及可育株G06-77-2B,时期1~2、3~4、5~6、7四个阶段花蕾各60、40、30、10个,电子天平称重,然后用解剖针分别剥出雌蕊和雄蕊,分别烘干、称重。计算出各样品不同蕾长时单个花蕾、花蕾雄蕊、花蕾雌蕊干质量。单位mg/蕾,精确到小数点后4位。
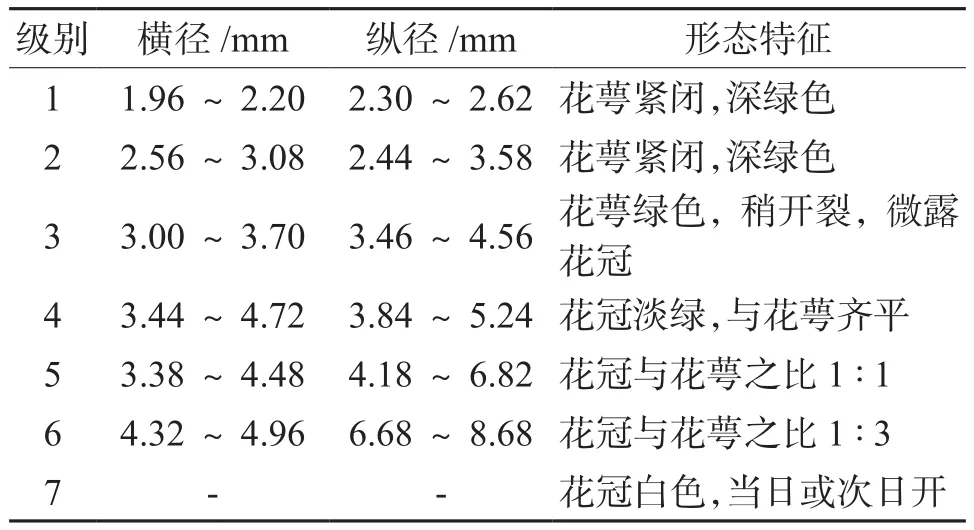
表2 不同时期花蕾的大小、形态特征
2.3 育性田间调查方法
每年6月在湖南长沙及11月在海南三亚对各个材料在盛花期时进行目测花粉量,根据所含花粉的多少将其划分为:(1)多花粉,自交结实率高,单果结籽多;(2)少花粉,自交结实率极低,单果结籽极少;(3)无花粉,花药干瘪无花粉散开,自交不结实或单性结实但无种子。
2.4 花粉量测定
取100个未开裂辣椒花药置于离心管中,30℃恒温下烘干花粉,待烘干并至花粉完全散粉后,向管中加5 mL 20%偏磷酸钠溶液,于涡漩振荡成悬浮液,然后取200 μL成悬浮液并稀释5倍,用微量进样器吸取千分之一体积的悬浮液滴在载玻片上,在倒置显微镜下统计10个视野中花粉粒数目,根据稀释倍数算出花粉粒数目,操作重复3次。
3 试验结果
3.1 雄性不育系G06-77-2AB与C06-77-2AB植株形态比较
从表2来看,两种辣椒不育系不育株的植株高度和开展度明显大于可育株,且差异显著;而两种不育系植株株高及开展度不存在显著性差异,仅C06-77-2A在株高和开展度数值上比G06-77-2A更大。两种不育系及可育株的始花在11~12节,叶片卵圆形,绿色,均无明显差异。因此,不育系C06-77-2A与G06-77-2AB不育株与可育株的植株形态比较整齐一致,但不育系株高和开展度较大,可能是不育系没有及时坐果,营养全部用于植株营养生长的结果。
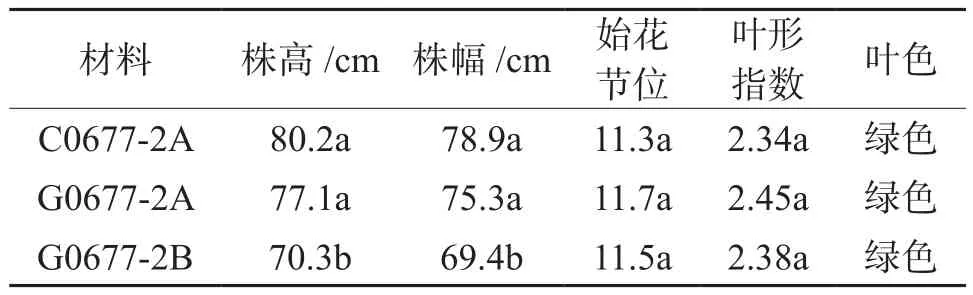
表2 两种不育系与可育系植株形态特性
3.2 两种雄性不育系G06-77-2AB与C06-77-2A花器官特性比较
由表3、图1可知,两种不育系C06-77-2A与G06-77-2AB的不育株和可育株花器官在花瓣数方面无显著差异外,其他方面如花开展度、花柱长度、花药长度、花药形态及花药鲜质量均有显著差异;两种不育系在花开展度方面没有显著差异,但是与可育系之间存在显著差异,不育系花开展度明显大于可育株,以细胞核质互作不育系开展度最大。两种不育系柱头长方面存在显著差异,以细胞核质互作不育系最长,且显著高于可育系。两种不育系花药长度则显著低于可育株,且两种不育系之间也存在显著差异,以细胞核质互作不育系C06-77-2A最短。两种不育系花柱在花蕾成熟待放前外露或在花药内呈扭曲状态。其中还可以观察到细胞核质互作不育系C06-77-2A花药皱缩、干瘪,成熟后为褐紫色或灰白色,不开裂或极少开裂,几乎无花粉;而细胞核不育系G06-77-2A花药长度明显大于C06-77-2A,且花药干瘪、不皱缩,发育成熟后呈紫褐色;可育株花药饱满,呈淡紫色,成熟后易开裂,花粉散出后布满整个花药,如图1所示。人工授粉证明不育株雌蕊正常,子房、胚珠和胎座发育健全。
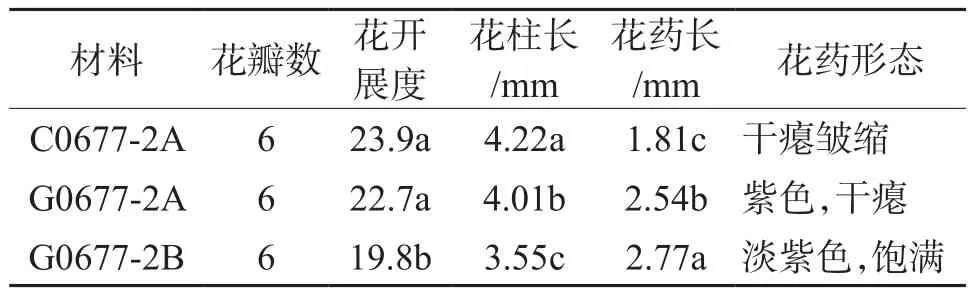
表3 两种雄性不育系G06-77-2AB与C06-77-2A花器官特性

图1 辣椒两种雄性不育系G06-77-2AB与C06-77-2A花器官形态
3.3 两种不育系花蕾及雌雄蕊比较
3.3.1 两种不育系花蕾发育动态
图2表明:
(1)从不同不育系类型比较,两种不育系花蕾质量在各个时期均较保持系(可育株)轻。其中两种不育系比较,C06-77-2A花蕾从时期2开始就较G06-77-2A花蕾轻,一直到花蕾发育成熟。G06-77-2B、G06-77-2A及C06-77-2A的花蕾自身从时期2到时期6质量增加分别为4.4倍、4.2倍、4.1倍,说明辣椒保持系(可育株)花蕾增加的质量较不育系快。
若以不同发育时期两个不育系的花蕾质量为一组,可育对照花蕾质量为另一组,运用DPS进行成对数据的t测验,则辣椒雄性不育系的花蕾干质量明显轻于可育系的花蕾,两者花蕾的质量差异达到5%显著水平。
(2)不同类型雄性不育系花蕾的发育模式不同。细胞核不育系G06-77-2A及可育株花蕾G06-77-2B从时期1到时期7质量增加的过程呈“∽”形,花蕾质量主要的增长时期是在时期3到时期6,增长的速度最快,花蕾发育在时期2以前及时期6以后增速明显放慢。发育曲线出现两个拐点,前后两个拐点均明显,拐点出现的时期分别在时期2和时期5,两个拐点之间花蕾的发育速度明显大于两头,蕾质量也明显大于C06-77-2A不育系。
(3)不同类型的雄性不育系与可育系花蕾质量的差异值不同。细胞核不育系与可育株花蕾从时期1到时期7质量一直很接近,数值相差很小,各时期差值在5%之内,而细胞核质互作不育系C06-77-2A各个时期花蕾质量均明显小于保持系G06-77-2B,最大差值在时期5,质量差值达到11.7%。
3.3.2 两种不育系雄蕊发育动态
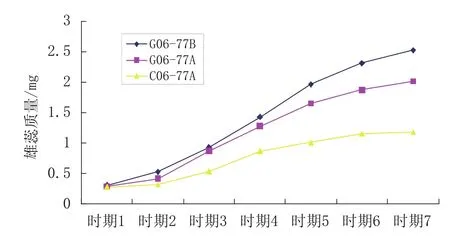
图3 两种不育系雄蕊发育动态
图3表明:
(1)不育系雄蕊质量在各时期都比其对应的可育系轻。 图3中两个不育系雄蕊发育曲线的位置明显低于可育系,从上往下依次是G06-77-2B、G06-77-2A及C06-77-2A,雄蕊干质量以G06-77-2B最大,为2.524 mg/蕾;其次是G06-77-2A,雄蕊干质量2.010 mg/蕾;雄蕊干质量以C06-77-2A最轻,为1.280 mg/蕾,只有正常雄蕊质量的一半左右。两个不育系及保持系雄蕊干质量的排序与花蕾干质量的排序基本一致。当雄蕊干质量到达最大值时,各自增重的倍数为:G06-77-2B增加7.1倍,G06-77-2A增加5.9倍及C06-77-2A增加3.2倍。
若将不同发育时期两个不育系雄蕊质量与一个可育系雄蕊质量进行对比分析,运用DPS统计分析软件,进行成对数据的t测验,细胞核不育系及可育系雄蕊的质量高于雄性细胞核质互作不育系,三者之间干质量差异达到1%显著水平。
(2)不同类型的雄性不育系雄蕊的发育模式不同。从不育系雄蕊发育曲线拐点出现的情况来看,两个不育系类型及保持系(可育株)可分为两类:细胞核不育系G06-77-2A与G06-77-2B为第一类,雄蕊发育曲线两个明显的拐点在时期2和时期5,雄蕊发育呈现为两头慢、中间快。细胞核质互作不育系C06-77-2A为第二类,雄蕊发育曲线仅在时期4出现一个拐点,之前雄蕊质量增加较快,拐点之后雄蕊质量增加缓慢,推测雄蕊花粉已经发生了解体。第一类雄蕊的发育曲线与其花蕾发育的曲线基本一致。
(3)不同类型雄性不育系与可育系雄蕊质量的差异值不同。两种不育系类型雄蕊的最大差异值均出现在花蕾时期7,此时,G06-77-2B雄蕊分别比G06-77-2A、C06-77-2A雄蕊干质量高25.4%和113.5%,细胞核质互作不育系C06-77-2A与 G06-77-2B在时期3开始出现差异,而细胞核雄性不育系G06-77-2A与 G06-77-2B的开始出现差异时期在时期4,估计和不同类型不育系发育有关。
3.3.3 两种不育系雌蕊发育动态

图4 两种不育系雌蕊发育动态
图4表明:
(1)在雌蕊发育过程中两个不育系的雌蕊质量均比其对应的可育系高。
图4中两个不育系雌蕊曲线的位置均在一个可育系雌蕊曲线上面,在发育时期内,两个不育系的雌蕊质量均比其对应的可育系大。依据雌蕊质量高低排序依次为:C06-77-2A、G06-77-2A、G06-77-2B,不育系雌蕊的自身增重倍数也比可育系大。
将雌蕊发育不同时期两个不育系雌蕊质量与一个可育系雌蕊质量进行对比分析,t测验,表明辣椒雄性细胞核质互作不育系雄蕊的质量明显大于细胞核不育系及可育系,三者之间干质量的差异达到1%显著水平。
(2)不同类型的雄性不育系雌蕊的发育模式不同。从雌蕊发育曲线拐点出现情况来看,两个不育系可分为两类:C06-77-2A雌蕊发育曲线拐点出现不明显,在花蕾时期4前的一段时间内,2个供试不育系中,C06-77-2A的雌蕊质量最大。G06-77-2A为另一类,雌蕊发育曲线出现了一个明显的拐点,拐点出现在时期5左右,拐点前后发育速度显著不同。在拐点出现之前雌蕊的发育慢,而出现拐点后的发育加快明显。
3.3.4 两种雄性不育系G06-77-2AB与C06-77-2A果实性状比较
由表4可以看出,两种不育系经可育株人工辅助授粉后,果形方面两者表现基本相似,味辣,均为小牛角形,但在果实横径、纵径、单果质量、果肉厚等单位性状方面两种不育系略微高于可育株,差异均未达显著性水平。
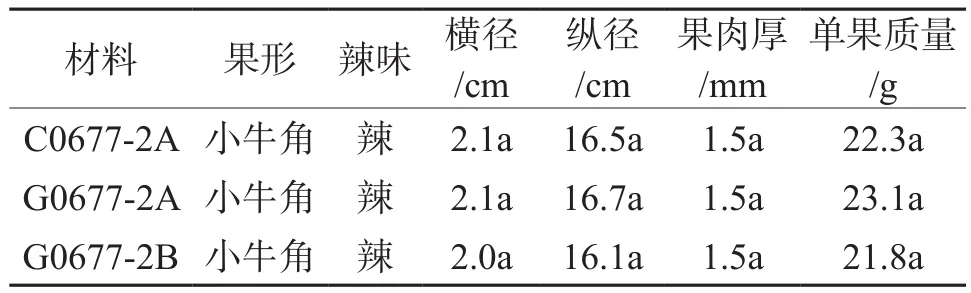
表4 辣椒两种雄性不育系G06-77-2AB与C06-77-2A果实性状
综上所述,随着细胞核质互作不育系转育代数的增加及核不育系兄妹交代数的增加,选育获得的辣椒细胞核质互作不育系C06-77-2A及细胞核雄性不育系G06-77-2AB的整体性状优良,个体间表现基本一致,能够运用于不育性状的回交转育亲本或选配杂交一代亲本。
4 讨论
4.1 辣椒雄性不育系雌蕊的异常膨大
本研究在花蕾发育的各个时期,两个不同类型辣椒雄性不育系雌蕊的质量均高于各自相对应的可育系。从花蕾发育时期1至时期7,两个不育系雌蕊质量自身平均增加了10.8倍与8倍,而同期对应的1个可育系自身仅平均增加7.4倍,不育系雌蕊比各自对应的保持系雌蕊的平均质量大26.9%。不育系雌蕊质量占花蕾质量的比例增长较快,如C06-77-2A,不育系雌蕊质量比例从时期1时的8.1%增加到了时期7时的16.1%,将近增加了一倍;而同期相对应的可育系雌蕊质量比例从时期1时的11.3%只增加到了时期7时的13.4%,仅增加2.1%。细胞核不育系G06-77-2A也存在这一现象,只是没有细胞核质互作不育系C06-77-2A明显,显示辣椒不育系的雌蕊发生了异常膨大的现象。
4.2 雄性不育系的花蕾及雄蕊干质量均小于其相应的可育系
聂明建等[5]对甘蓝型油菜雄性不育系的花蕾及雌雄蕊干物质变化动态进行了研究,结果表明,雄性不育系花蕾和雄蕊干物质质量显著小于保持系,而雌蕊则明显大于保持系。本研究结果显示:辣椒两种雄性不育系雄蕊质量在时期2开始出现差异,在时期3至时期5差异迅速加大,到时期7达到顶峰;两个不同类型辣椒雄性不育系的花蕾和雄蕊干质量及花药长度都小于其可育系。统计分析表明,两对不育系与其各自对应的可育系花蕾及雄蕊质量差异均达1%显著水平,这一研究结果表明,不育系雄蕊在花蕾长度时期3时就与可育系雄蕊有显著差异。
4.3 不育系雄蕊败育后的物质可能被部分转移到了雌蕊
聂明建等[5]还认为:大多数不育系雌蕊雄蕊的发育曲线存在拐点,其出现拐点的时期相同,但方向完全相反。暗示可能雄蕊败育后降解的生物大分子物质被部分运到了雌蕊,才引起了雌蕊干质量的异常增加。在本研究中辣椒雄性不育系雌蕊和雄蕊的发育曲线上都有拐点,而且雄蕊出现拐点的不育系其雌蕊发育曲线同样有拐点出现,其出现的时期基本一致。只不过在拐点出现后不育系雄蕊和雌蕊的发育速度发生了明显变化,雄蕊速度降低而雌蕊速度加快。雄蕊发育曲线无明显拐点出现的不育系,则雌蕊也无明显拐点,如C06-77-2A雌、雄蕊发育曲线均不出现拐点。笔者认为这种情况应该不是一种简单的巧合,很有可能是在雄蕊败育后降解的生物大分子物质被部分转移到了雌蕊上,从而出现雌蕊异常膨大情况。正是由于雄性不育雌雄蕊的相互关系,才使辣椒雄性不育系雌雄蕊的发育曲线同时出现了完全相反方向上的拐点。
在试验中,雄性不育系最显著的特点是雄蕊发育落后,明显小于可育系,而雌蕊异常膨大,明显大于可育系。不育系雌蕊的异常膨大与正常受精后的膨大有着某些相似之处,只不过正常辣椒雌蕊被授粉后还可继续发育直到成熟,而雄性不育系雌蕊没有受精,虽有膨大但不会成活。在此特别说明本试验由于取样时正常辣椒花粉未成熟,还不能正常授粉受精,所以子房不会膨大。
在以往辣椒雄性不育系的研究中,人们主要关注的是雄蕊,相比雌蕊的关注较少。事实上,在雌雄同花作物中,雌雄蕊之间有着千丝万缕的关联,两者为一个整体,任何一方发育异常都会打破平衡,都会影响到对方。辣椒雄性不育系花蕾发育中,雌蕊发育速度的异常加快,是造成雄蕊败育的原因,还是导致雄蕊败育的结果?也就是说,在雄蕊败育过程中,雌蕊的作用是主动夺取雄蕊养分,还是被动接收雄蕊败育后外运的养分,孰因孰果,还需要经过进一步深入仔细地研究后,才能得出结论。因此,在雄蕊败育过程中雌蕊在这中间究竟起到了什么作用应该引起广泛的关注。
[1]耿广东, 姜虹, 杨红, 张素勤. 辣椒胞质雄性不育分子生物学研究进展[J]. 安徽农业科学, 2012(1): 41-43
[2]唐冬英, 邹学校, 刘志敏. 辣椒雄性不育的研究和利用进展[J]. 湖南农业科学,2002(4):15-18
[3]袁志勤, 边正午, 韩万禄. 辣椒花粉母细胞减数分裂过程的观察[J]. 河北农业大学学报,1989, 2(12): 93-98
[4]何长征. 辣椒细胞质雄性不育的细胞学及生理机理研究[D]. 长沙: 湖南农业大学,2004
[5]聂明建,王国槐, 陈光尧. 五种不同类型甘蓝型油菜雄性不育系花蕾及雌雄蕊干物质变化动态的比较研究 [J].武汉植物学研究, 2006(5): 446-450
Comparison on Agronomic Traits of Two Types of Male Sterile Lines in Hot Pepper
HUANG Wei1,2MIAO Wu3
(1. Hunan Institute of Vegetables, Changsha 410125, China; 2. Hunan Xingshu Seed Industry Co., Ltd.,Changsha 410125, China; 3. Hunan Xiangyan Seed Industry Co., Ltd., Changsha 410100, China)
In order to understand the agronomic traits of two types of hot pepper male sterile lines, a genic male sterile (GMS) line G06-77-2A, a cytoplasm-nuclear male sterile (CMS) line C06-77-2A and a fertile line G06-77-2B of the two male sterile lines were analyzed on plant height, plant expansion, bud, stamen and pistil in this study. The results showed that (1) the other traits of the two male sterile lines except being taller in plant height and larger in plant canopy were basically the same as those of the male fertile line; and (2) the weights of flower bud and stamens of the male sterile lines were less than those of their respective fertile lines, but the pistil of the male sterile lines appeared abnormally enlarged, in which the CMS line was more prominent than the GMS line,and there was no signi fi cant difference in their fruits. It was concluded that the CMS line C06-77-2A and the GMS line G06-77-2A were of overall superior traits, with basically consistent individual performance in them, and could be used each as a parent in the backcross transformation of sterile trait or a parent in the selective breeding of hybrid F1.
hot pepper; CMS; GMS; agronomic trait; comparison
2017-05-19
黄巍(1978-),男,助理研究员,主要从事蔬菜品种选育及新品种推广
