老鸦瓣属植物资源及分子系统学研究进展
2017-11-09陈绍通宋向文韩邦兴
陈绍通,刘 枫,宋向文,赵 群,韩邦兴
(1.安徽中医药大学,安徽 合肥 230012;2.皖西学院,安徽 六安 237012;3.安徽省石斛产业化开发协同创新中心,安徽 六安 237012)
老鸦瓣属植物资源及分子系统学研究进展
陈绍通1,2,刘 枫1,2,宋向文2,3,赵 群2,3,韩邦兴2,3
(1.安徽中医药大学,安徽 合肥 230012;2.皖西学院,安徽 六安 237012;3.安徽省石斛产业化开发协同创新中心,安徽 六安 237012)
老鸦瓣属AmanaHonda为百合科药用植物属,为东亚特有类群。有关老鸦瓣属的研究相对较少,且多集中于物种老鸦瓣Amanaedulis(Miq.) Honda的化学成分、组培、生物学特性等方面,而对于老鸦瓣属内其他种的研究报道极少。本文主要综述了该属植物的系统位置、属内种的分类学地位、生物学特性及分子系统学等多个方面的研究进展。基于老鸦瓣属植物已有的和本课题组前期初步研究的基础,并针对该属中存在的关键科学问题,对可能采用的研究方法提出展望,其研究思路为老鸦瓣属药用植物资源开发奠定基础。
老鸦瓣;老鸦瓣属;植物资源;生物学特性;分子系统学
药用植物老鸦瓣Amanaedulis(Miq.) Honda隶属于郁金香属AmanaHonda,其去除绒毛后的干燥鳞茎为中药“光慈姑”的来源,具有解毒散结、行血化瘀的功效,主治咽喉肿痛、瘰疬和产后瘀滞等症状[1](P361)。由于文献记载混乱等原因,老鸦瓣的药用历史也经历了一系列的变迁。在晋代,鳞茎作为“贝母”入药,但因其有毒,被列为“贝母”的伪品,称为“丹龙睛”;到了宋代,又作为药材“山慈姑”来源入药。此外,古代老鸦瓣鳞茎入药并不去除绒毛,因此又称“毛慈姑”,直到近代,人们在采收加工的时候,为了提高药材的疗效,去除毛壳而改称“光慈姑”[2]。
老鸦瓣属AmanaHonda隶属于百合科Liliaceae,由Honda从郁金香属TulipaL.分离而成立[3],为东亚特有类群,仅分布于中国中东部,日本和朝鲜半岛[4]。老鸦瓣属在百合科中分类地位虽然得到了一些学者的承认[5-6],[7](P38),[8](P925),但对于老鸦瓣属是另立还是归入郁金香属Tulipa的争论仍然存在。此外,面对市场对老鸦瓣的需求量日益增加,而野生自然资源正逐渐下降等矛盾,如何提高老鸦瓣的产量成为人们越来越关注的问题。而人工栽培、组织培养、寻找新药源等方法也成为当前解决问题的重要途径。因此,本文基于课题组前期对老鸦瓣属资源的调查及初步研究,对老鸦瓣属的分类、生物学特性及分子系统学进行综述,为老鸦瓣属的分类及老鸦瓣栽培提供了数据支持,也为寻找新药源及今后产业化的发展奠定基础。
1 老鸦瓣属植物资源
1.1 资源特征及分布
老鸦瓣属Amana隶属于百合科,多年生草本,由Honda从郁金香属分离而成立,在形态上与郁金香属的主要区别在于:花被片背面具有紫红色纵向条纹,花柱与子房近等长,具2~4个对生或轮生苞片,叶常2枚对生且两面无毛[4]。目前,已发现的该类群共5种,分别为老鸦瓣Amanaedulis、二叶郁金香Amanaerythronioides、安徽老鸦瓣Amanaanhuiensis、括苍山老鸦瓣Amanakuocangshanica和皖浙老鸦瓣Amanawanzhensis[9](P89) [10-13]。该属中植物均为早春植物,繁盛生长期为每年的3~4月,5月开始倒苗,喜温暖潮湿环境,以老鸦瓣A.edulis分布最广,而属中其他物种分布较为狭窄且集中,主要以安徽、浙江地区为分布中心,多为各个地区的特有物种,可见图1。
老鸦瓣Amanaedulis(Miq.) Honda为多年生草本,叶通常两片,线形,长达30 cm;花单朵顶生;苞片通常2枚对生,地下部分为椭圆形鳞茎,外包赤褐色膜质鳞片,内密被长柔毛。我国中东部大部分地区(安徽、江苏、浙江、山东、江西、湖北、湖南、陕西、辽宁)都有分布,且多生长在丘陵、岗地等向阳的草地或山坡上。目前也作为治疗肿瘤的常用药[14]。

图1 中国老鸦瓣属植物主要分布区域
二叶郁金香Amanaerythronioides(Baker) D. Y. Tan et D. Y. Hong,又称二叶老鸦瓣、宽叶老鸦瓣,与老鸦瓣相似,同为多年生草本,以叶片和苞片类型而区别于同属的其他种。本种叶两枚,较宽而短,长度在7~15 cm之间,两片叶宽也不相同,宽者常在15~22 mm,窄者常为9~15 mm;苞片通常3或4枚轮生(较少2枚对生)。多生长于海拔600~1 300 m的高山,目前,除安徽、浙江之外,2012年,在江西省云居山也有发现,为江西地理新分布物种[15]。
2001年,沈显生在安徽天柱山发现郁金香属的一新种,并命名为皖郁金香TulipaanhuiensisX. S. Shen。2008年,潭敦炎[16]等对该种新模式进行指定和补充,并重新组合命名为安徽老鸦瓣Amanaanhuiensis(X. S. Shen.) D. Y. Tan et D. Y. Hong。该种为安徽天柱山地区的特有种,株高7.5~15 cm之间,生长在海拔850~1250 m的落叶阔叶林下,溪沟边或灌木丛中,阴湿环境下生长较多。与同属其余种的形态差异在于鳞茎皮薄纸质,内侧被绵毛;苞片(2~)3(~4)枚,轮生(或对生),第一叶长9.6~22.4 cm,宽1.6~2.5 cm,且从基部向上3/4处最宽(潭敦炎等, 2008)。沈显生认为本种与二叶郁金香A.erythronioides近缘,但花被片披针形,花药淡紫色,花柱1~2 mm,柱头歪斜可与之区别[11]。
2007年,Tan等在浙江括苍山地区发现一新种——括苍山老鸦瓣AmanakuocangshanicaD. Y. Tan & D. Y. Hong,分布在海拔600~1 100 m。在形态学上,该种与安徽老鸦瓣和二叶郁金香相似,不同的是地下鳞茎部分皮内侧无毛,叶片最宽处在基部向上的2/3处,果喙长(6.4±0.8)mm。此外,TAN通过17个形态特征对A.edulis、A.erythronioides、A.anhuiensis、A.kuocangshanica进行UPGAM聚类分析,结果发现A.edulis聚在最外支,A.anhuiensis与A.kuocangshanica互为姐妹群,并与A.erythronioides形成并系分支[12]。
2014年,Han等人在安徽宁国的仙霞镇发现老鸦瓣属的另一新种——皖浙老鸦瓣AmanawanzhensisL. Q. Huang, B. X. Han & K. Zhang。该种多生长在600~800 m的竹林下、灌丛中或草地上,鳞片卵形,直径1.5~2.5 cm,内部棕色纸质,柔毛,茎高15~30 cm,苞片通常3枚,不轮生,带状,长0.1~0.5 cm。Han等认为该种与二叶郁金香A.erythronioides相似,但通过苞片长度较短(0.1~0.5cm),花药黄色,花被片易脱落可与其区分开。而该结论也获得了分子系统学的支持,通过ITS+trnL intron联合序列对该属(除括苍山老鸦瓣A.kuocangshanica)之外的其余四种进行分子发育研究,结果显示该种独自形成单支,且获得了高度的支持(BS=93, PP=1.00),相对于老鸦瓣,皖浙老鸦瓣与二叶郁金香聚为一支(BS=79, PP=0.95),由此也说明了,该新种是成立的,且与二叶郁金香亲缘关系更近[13]。
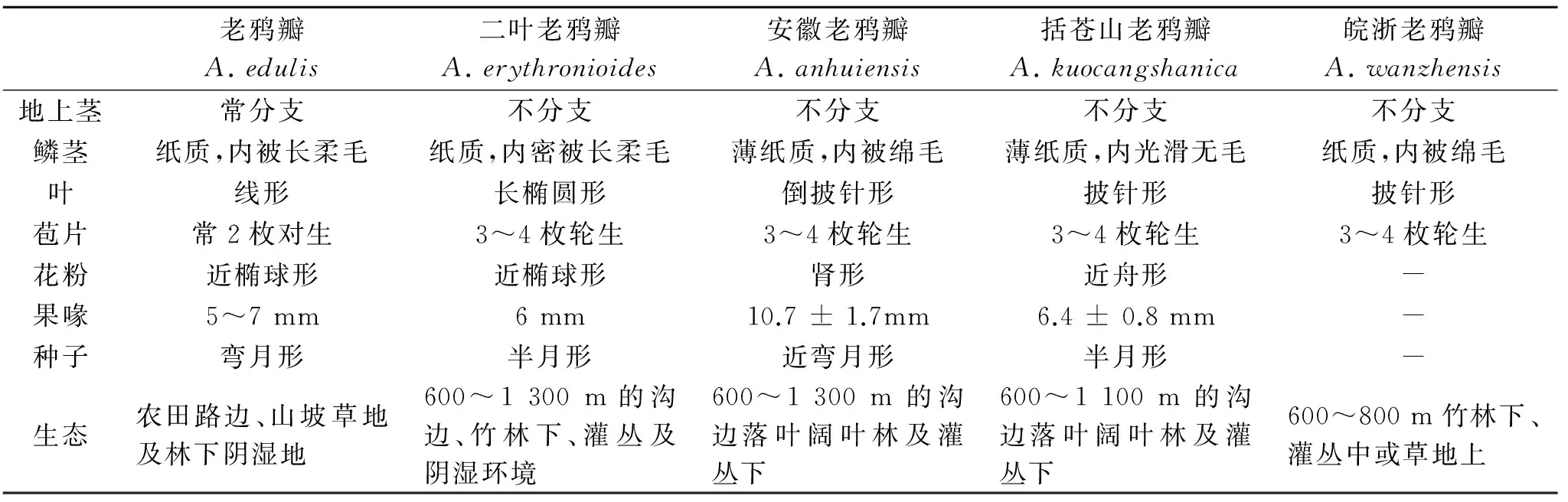
表1 老鸦瓣属内部形态特征比较[17]
1.2 老鸦瓣属植物分类沿革
近年来,对于老鸦瓣属Amana的系统发育和分类地位一直备受争议。自Honda建属以来,关于该类群是隶属于郁金香属Tulipa还是另立的问题也一直是争论的焦点。其中,较为重要的分类方法当属Miquel[18],Baker[19],Honda[5],Hall[3],Sealy[20],Hoog[21],毛祖美[22](P86-97),Kitamura[6],Satake[7],Ohwi和Kitagawa[23],梁松筠[24],Tamura[25],吴征镒[8],谭敦炎[4],韩邦兴[13]等,可见表2。

表2 老鸦瓣属分类研究
1.3 生物学特性
老鸦瓣属为早春植物,对光照、温度等自然条件要求较高,多生长在向阳的山坡、草地和水边[26]。除了老鸦瓣之外,其余种均分布在海拔较高的山区,如二叶郁金香多生长在海拔600~1300 m的落叶阔叶林下,安徽老鸦瓣生长在海拔850~1250 m的竹林、落叶阔叶林或水边,部分也长在向阳的石头含有腐殖质较多的潮湿处。皖浙老鸦瓣多生长在600~800 m的竹林下、灌丛中或草地上。而老鸦瓣在平原、路边及高海拔地区都有分布。目前,由于老鸦瓣属物种分布的限制,在对于老鸦瓣属的生物学特性的研究中,以老鸦瓣的研究最多,而其余种的研究极少见到报道。
老鸦瓣具有典型的百合科花部特征,属兼性异交类型,异交为主,自交为辅,主要以昆虫为老鸦瓣传粉媒介[27]。梁丽[28]等对老鸦瓣的光合特性进行研究发现,老鸦瓣的光补偿点为1.782 umol/(m2·s),光饱和点约为1 500 umol/(m2·s),表现光量子产额为0.025,说明老鸦瓣具有阴性的特点,对光能的生态适应范围较广,但不宜强光直射。此外有研究发现,在78%左右的光强最适宜老鸦瓣生长[29]。
老鸦瓣种子属胚形态休眠为主的综合休眠型,且刚成熟老鸦瓣种子胚率仅为10%左右。吴正军[27]等为提高种子萌发率,对老鸦瓣种子的萌发特性进行研究,结果发现,通过300 mg·L-1赤霉素浸泡处理24 h后,25 ℃恒温层积70天,再转入5 ℃低温层积可使种胚成熟最快,破眠之后的种子在10、15 ℃的黑暗条件下萌发率可达90%以上。
除了种子繁殖之外,老鸦瓣也可通过鳞茎繁殖,而且相对于种子繁殖,鳞茎繁殖过程所需周期更短。但是,与种子繁殖相同的是,如何打破鳞茎休眠,延长老鸦瓣的生长发育时间是提高产量的关键。杨颖[30]等发现冷藏处理有利于鳞茎提前出苗,并显著缩短出苗时间且利于苗齐,但对出苗率影响不明显,且冷藏后花发育不正常,结果率为0,并认为(4±0.5)℃冷藏6~8周为老鸦瓣鳞茎打破休眠的关键时期,冷藏可延长老鸦瓣生育期并提高鳞茎产量。缪媛媛[31]等通过摘蕾和人工授粉处理老鸦瓣,发现摘蕾有利于老鸦瓣鳞茎生物量的积累从而提高药材产量,而人工授粉显著提高老鸦瓣结实率及种子数量和单粒重。杨小花[32]等根据鳞茎的重量设4个级别:1级(重量>2 g)、2级(重量=1~2 g)、3级(重量=0.5~1 g)、4级(重量<0.5 g),并在生长旺期测量老鸦瓣的生长指标,结果发现3级和4级种球增产效果明显,作为鳞茎繁殖最佳。同时,作者也对老鸦瓣的采收期研究发现,老鸦瓣应在枯萎期或休眠期初期采收为佳[33]。除了上述之外,对于老鸦瓣的组织培养也有报道[34-35]。
2 老鸦瓣属分子系统学研究
近年来,随着测序技术的迅猛发展和大量的DNA数据积累,生物系统学也由早期的利用化石证据、表型形态特征或者解剖结构性状推演生物间系统关系,转入分子系统学时代[36]。目前,分子系统学已成为植物系统发育中的研究热点。相对于传统的方法,分子系统学在面对部分种间形态相似或种内变异较大的疑难物种时,成为学者们选择的重要技术手段,既能弥补传统的不足,又能为植物的系统发育研究提供新的层面的数据支持。在传统意义上,分子系统学可分为两部分,即在种上水平的系统发生学(Phylogenetics)和种下水平的群体遗传学(Population genetics),但随着系统地理学、DNA barcoding理论的相继出现,系统发生学的体系也得到了迅速扩大[37]。
2.1 系统发育研究
自21世纪以来,老鸦瓣属的新种相继被发现,由最初建属的2个种(A.edulis和A.erythronioides)到目前的5个种:沈显生[11]发现该属的第3个物种皖郁金香(即安徽老鸦瓣);Tan[12]等发现该属的第4个物种括苍山老鸦瓣;Han[13]等发现第5个物种皖浙老鸦瓣。越来越多的新种被发现,也为老鸦瓣属的分类研究提供了更多的材料。但是,相对于同科的其他的属来说,如郁金香属Tulipa[38]、百合属Lilium[39]、猪牙花属Erythronium[40]等,该属的系统发育研究近乎空白,仅在对百合科其他类群的系统发育研究中,对该属少数种类有过一些零星报道。
谭敦炎[4]等利用ITS和trnL-F序列对广义郁金香属的5个组进行系统发育分析,其中sect.Tulipanum、sect.Leiostemones、sect.Eriostemone和sect.Orithyia聚为一支,而老鸦瓣属Amana类群(包括A.edulis、A.erythronioides、A.anhuiensis、A.kuocangshanica)单独聚为一支,且获得高度的支持(BP=99),并与猪牙花属Erythronium形成并系,聚为一支(BP=100)。但研究结果只说明了老鸦瓣属和郁金香属的物种位于不同的谱系分支,并没有对老鸦瓣属内的系统发育关系进行分析,且未涉及现今发现的所有老鸦瓣属物种。
Allen[41]根据matK基因序列对Erythronium属分析时发现,老鸦瓣Amanaedulis与郁金香属中的其他两个种Tulipamontanavar. chrysantha和Tulipaturkestanica并未形成一支。同样,JOHN[40]利用联合片段(ITS,matKandrps16 introns)对Erythronium属分析,也得到相同的结果,都表明Amana类群独立成一支。但不足的是,研究所用的物种只是包含了老鸦瓣属中的一个或两个,而对于老鸦瓣属整体的研究尚未见到报道。Christenhusz等[38]对郁金香属进行了分子系统学研究,其中老鸦瓣属仅作为外群参与了系统发育树的构建,并没有做进一步的系统发育分析,且老鸦瓣属仅包括老鸦瓣和二叶郁金香2个种。Nguyen等[42]以6个DNA分子片段为分子标记,对百合目的Colchicaceae科进行了系统发育分析,老鸦瓣同样仅作为其中一个外群参加了构树,并没有分析和其他物种之间的系统发育关系。
本课题组前期也对老鸦瓣属的5个种系统发育进行初步研究(未发表),基于nrDNA和cpDNA联合序列构建系统发育树,结果显示,5个来自老鸦瓣属的种都各自形成单系分支。在系统发育树中,老鸦瓣属起先形成两大分支,一个分支由老鸦瓣和括苍山老鸦瓣组成;另一分支由二叶郁金香、安徽老鸦瓣和皖浙老鸦瓣组成,其中安徽老鸦瓣与皖浙老鸦瓣亲缘关系更近,形成姐妹类群,与二叶郁金香再形成并系。
2.2 生物地理学研究
生物地理学(Biogeography)是地理学和生物学的交叉学科,是研究生物分布及分布规律的学科,可分为历史生物地理学和生态生物地理学。其中,生态生物地理学主要以在短时间跨度内空间因子的作用,即自然因素的作用;而历史生物地理学是在大的时间跨度内(百万年),讨论生物的进化历程[43]。近30年来,随着分子生物学的技术改革与进步,促进了历史生物地理学的迅猛发展。1987年,Avise等在早期历史生物学基础上,建立了新的分支—谱系生物地理学(Phylogeography),主要通过对基因的谱系分析和单倍型的时空分布探讨生物与古地质事件(如冰川、地理隔离等)之间的关系,推测物种的起源、分化过程[44-46]。
Han[13]等基于核序列ITS和质体序列trnL intron的部分DNA片段分析了老鸦瓣属4个物种之间的系统发育关系,并证明皖浙老鸦瓣为有效种。然而,在分子系统发育分析中并未包括苍山老鸦瓣,且每个种的样本量较少,同时也没有对该属的属内分化时间和生物地理学进行分析。根据不同物种间的系统发育关系以及各物种的地理分布,可以分析物种的生物地理格局,探讨各物种形成现有地理格局的原因,能够更好地理解物种宏进化机制,同时也能对重要的地质历史事件进行印证[47-51]。所以,有必要对老鸦瓣属的分子系统发育学和生物地理学进行深入的研究。
Vinnersten和Bremer[52]基于叶绿体rbcL序列对百合目主要分支分化时间和生物地理学进行了研究,结果表明郁金香属和猪牙花属的分化时间在13±5.6 mya(million years ago;百万年前)。尽管没有老鸦瓣属的分化时间,但在后续研究中可以首先建立老鸦瓣属与郁金香属和猪牙花属之间的系统发育关系,并利用建立的系统发育树,以郁金香属和猪牙花属的分化时间为时间标定点计算老鸦瓣属的分化时间和属内各物种之间的分化时间。由于老鸦瓣属和猪牙花属的亲缘关系更近[4][39],可以推测老鸦瓣属可能起源于中新世后期,情况是否如此,需要进一步研究才能确定。
谭敦炎[4]等认为中国的浙江和安徽是老鸦瓣属植物的分布中心,Wu[8]等认为老鸦瓣属在中国、日本和朝鲜半岛的隔离分布显示其古老性。由于地质史上较为严重的冰期会导致海平面下降,在此期间台湾可能会和亚洲大陆通过陆地相连,而琉球群岛南部则可能和台湾东北部的陆地相连。这样,在某一个冰盛期,亚洲大陆、台湾和琉球群岛可能是短暂相连的;当冰期结束后,海平面上升,三者之间又会被海面隔离[53]。自从中新世这些岛屿形成以来,短暂的陆地通道的形成和消失已经重复了很多次。因此,越海扩散技能很差的生物物种可能会通过以上冰盛期时短暂的陆地连接通道在亚洲大陆、台湾和琉球群岛之间扩散[54-55]。而老鸦瓣属的分化中心地理位置,以及该属的隔离分布是否存在中国、朝鲜半岛和日本之间在冰盛期的跨海扩散,目前未见报道,需要在这方面做更多的研究。
3 结语
3.1 分类问题
目前,老鸦瓣属Amana在百合科中分类地位虽然得到了一些学者的承认,但对于老鸦瓣属是另立还是归入郁金香属Tulipa的争论仍然存在。随着新种的相继发现,老鸦瓣属已有5个物种,以我国的安徽分布最广,包含4个种。由于物种分布的限制,目前,对于老鸦瓣属的分子系统学研究相对较少,相关的研究也只是对该属内部分种,而针对老鸦瓣属整体来说,并未见到报道。因此,本课题组对该属5个植物种类,用4个DNA序列为分子标记构建老鸦瓣属的系统发育树,确定系统树中各个节点(祖先物种)的分化时间,同时确定各个节点的祖先物种可能的地理分布区,结合各祖先物种当时的古地质条件和古气候条件分析该属的地理起源和物种扩散路径和方向,探讨该属的起源及属内分化原因。
3.2 资源及产业化
近年来,市场对老鸦瓣的需求量日益增加,而野生自然资源正逐渐下降,供求失衡,矛盾日益尖锐。如何提高老鸦瓣的产量成为人们越来越关注的问题。而人工栽培、组织培养、寻找新药源等方法也成为当前解决问题的重要途径。目前,对于老鸦瓣的生物学特性的研究已经进入成熟阶段,不论在栽培和组织培养方面,都有研究报道。特别是在如何提高老鸦瓣产量的人工栽培上,其方法学也已成熟。此外,本课题组已对老鸦瓣属的分子系统学展开初步研究,并对于市场中常见的老鸦瓣伪品进行分子鉴别的研究。所以,大量的基础研究为老鸦瓣的栽培提供了数据支持,也为寻找新药源及今后产业化的发展奠定基础。
[1]谢宗万.全国中草药汇编(上册)[M].第2版.北京:人民卫生出版社,1996,361.
[2]王惠民.老鸦瓣药用历史的变迁[J].中药材,1992(6):35-36.
[3]Honda M.Amana, A New Genus of Liliaceae[J]. Bulletin of the Biogeographical Society of Japan, 1935(6): 19-21.
[4]谭敦炎,张震,李新蓉,等.老鸦瓣属(百合科)的恢复:以形态性状的分支分析为依据[J].植物分类学报,2005(3):262-270.
[5] Hall A D, Beal J M. The Genus Tulipa.[J]. International Journal of Plant Sciences, 1940(1):143-146.
[6] Br.-Bl. Coloured Illustrations of Herbaceous Plants of Japan (Monocotyledoneae), by Kitamura S.; Murata G.; Koyama T.[J]. Vegetatio, 1966(5):289-289.
[7]Satake Y, et al.Wild Flowers of Japan (Herbaceous Plants) [M]. Tokyo: Heibonsha Ltd. Publishers, 1982.
[8]吴征镒,路安民,汤彦承,等.中国被子植物科属综论[M].北京:科学出版社,2003.
[9]陈心启,许介眉,梁桦筠,等.中国植物志:第14卷,被子植物门(单子叶植物纲),百合科(一)[M].北京:科学出版社,1980.
[10]梁松筠.百合科(狭义)植物的分布区对中国植物区系研究的意义[J].植物分类学报,1995(33):27-51.
[11]沈显生.中国郁金香属一新种[J].植物分类与资源学报,2001(23):39-40.
[12]Tan DY, Li XR, Hong DY.AmanaKuocangshanica(Liliaceae), a New Species from South-east China [J]. Botanical Journal of the Linnean Society, 2007(154): 435-442.
[13]Han BX, Zhang K, Huang LQ.AmanaWanzhensis (Liliaceae), a New Species from Anhui, China[J]. Phytotaxa, 2014(177):118-124.
[14]阮小丽,施大文.山慈菇的抗肿瘤及抑菌作用[J].中药材,2009,32(12):1886-1888.
[15]熊宇,钱萍,黄萌,等.江西百合科一新记录种——二叶郁金香[J].江西科学,2012(30):317-318.
[16]谭敦炎,李新蓉,洪德元.安徽老鸦瓣新模式的指定与补充描述[J].西北植物学报,2008(28):393-395.
[17]谭敦炎.中国郁金香属(广义)的系统学研究[D].北京:中国科学院(博士学位论文),2005.
[18]Tan D, Xinrong L I, Hong D. Amana Kuocangshanica (Liliaceae), A New Species from South-east China[J]. Botanical Journal of the Linnean Society, 2007, 154(3):435-442.
[19]Baker JG. Revision of the Genera and Species of Tulipeae[J]. Botanical Journal of the Linnean Society, 1874(14): 275-296.
[20]Sealy JR.Tulipaedulis[J] Curtis’s Botanical Magazine, 1957(171):293-293.
[21]Fiedler P L. Life History and Population Dynamics of Rare and Common Mariposa Lilies (Calochortus Pursh: Liliaceae)[J]. Journal of Ecology, 1988, 75(4):977-977.
[22]Mao ZM. Tulipa L. In: Flora Reipublicae Popularis Sinicae[M]. Beijing: Science Press, 1980.
[23]Nakaike T. New Flora of Japan[J]. Pteridophyta.rev. & Enlarg. 1992(1):417-418.
[24]梁松筠. 百合科(狭义)植物的分布区对中国植物区系研究的意义[J].植物分类学报,1995(33):27-51.
[25]Paul Wilkin. The Families and Genera of Vascular Plants. Volume III. Flowering Plants Monocotyledons Lilianae (Except Orchidaceae), by K. Kubitzki; H. Huber; P. J. Rudall; P. S. Stevens; T. Stützel[J]. Kew Bulletin, 1999, 54(4):1013-1013.
[26]宋向文,王德群,韩邦兴.安徽省老鸦瓣属药用植物资源[J].中国野生植物资源,2014(33):51-52.
[27]吴正军,朱再标,郭巧生,等.老鸦瓣传粉生物学初步研究[J].中国中药杂志,2012(37):293-297.
[28]梁丽,田霞,何云核.老鸦瓣光合生理特性研究[J].林业科技开发,2009(23):72-74.
[29]徐红建,朱再标,郭巧生,等.光强对老鸦瓣生长发育及光合特性的影响[J].中国中药杂志,2012(37):442-446.
[30]杨颖,朱再标,郭巧生,等.冷藏对老鸦瓣鳞茎休眠解除及栽后生长情况的影响[J].中国中药杂志,2015(40):48-52.
[31]缪媛媛,朱再标,郭巧生,等.摘蕾和人工授粉对老鸦瓣生长和产量的影响[J].中国中药杂志,2014(39):2016-2018.
[32]杨小花,缪媛媛,郭巧生,等.不同等级老鸦瓣生长繁殖特征研究[J].中草药,2015(46):3746-3750.
[33]杨小花,郭巧生,朱再标,等.不同采收期老鸦瓣生物量积累及药材品质研究[J].中国中药杂志,2016(41):624-629.
[34]朱丽芳,史俊,朱再标,等.老鸦瓣芽茎组织培养初步研究[J].中草药,2014(45):563-568.
[35]朱丽芳,徐超,朱再标,等.TDZ和NAA对老鸦瓣不定芽诱导和丛生芽增殖的影响[J].中国中药杂志,2014(39):3030-3035.
[36]Suárez Díaz E, Anaya Muoz VH. History, Objectivity, and the Construction of Molecular Phylogenies[J]. Stud Hist Phil Biol Biomed Sci, 2008(39): 451-68.
[37]徐宏发,王静波.分子系统学研究进展[J].生态学杂志,2001(20):41-46.
[38]Christenhusz M J M, Govaerts R, David J C, et al. Tiptoe through the Tulips-cultural History, Molecular Phylogenetics and Classification of Tulipa (Liliaceae)[J]. Botanical Journal of the Linnean Society, 2013, 172(3):280-328.
[39]Rønsted N, Law S, Thornton H, et al.. Molecular Phylogenetic Evidence for the Monophyly of Fritillaria and Lilium (Liliaceae; Liliales) and the Infrageneric Classification of Fritillaria[J]. Molecular Phylogenetics & Evolution, 2005, 35(3):509-527.
[40]JOHN CB, CLENNET, et al. Phylogenetic Systematics of Erythronium (Liliaceae):Morphological and Molecular Analyses[J]. Botanical Journal of the Linnean Society, 2012(170): 504-528.
[41]Allen GA, Soltis DE, Pamela S. Soltis. Phylogeny and Biogeography of Erythroniutn (Liliaceae) Inferred from Chloroplast matK and Nuclear rDNA ITS Sequences[J]. Systematic Botany, 2003(28): 512-523.
[42]Nguyen TPA, Kim JS, Kim JH. Molecular Phylogenetic Relationships and Implications for the Circumscription of Colchicaceae (Liliales)[J]. Botanical Journal of the Linnean Society, 2013(172): 255-269.
[43]赵铁桥.历史生物地理学进展[J].昆虫分类学报,1992(1):35-48.
[44] Avise JC, Arnold J, Ball RM, et al.. Intraspecific phylogeography: the mitochondrial DNA bridge between population genetics and systematics[J]. Ecology, Evolution, and Systematics, 1987(18): 489-522.
[45]Reem E, Douek J, Paz G, et al.Phylogenetics, Biogeography and Population Genetics of the Ascidian Botryllus Schlosseri, in the Mediterranean Sea and beyond[J].Molecular Phylogenetics & Evolution, 2017(107):221-231.
[46]Cardoso D, Harris D J, Wieringa J J, et al.. A Molecular-dated Phylogeny and Biogeography of the Monotypic Legume Genus Haplormosia, a Missing African Branch of the otherwise American-Australian Brongniartieae Clade[J].Molecular Phylogenetics & Evolution, 2017(107):431-442.
[47]Sakaguchi S, Qiu Y X, Liu Y H, et al. Climate Oscillation during the Quaternary Associated with Landscape Heterogeneity Promoted Allopatric Lineage Divergence of a Temperate Tree Kalopanax Septemlobus, (Araliaceae) in East Asia[J]. Molecular Ecology, 2012, 21(15):3823-3823.
[48]Wang W, Lin L, Xiang X G, et al.. The Rise of Angiosperm-dominated Herbaceous Floras: Insights from Ranunculaceae[J]. Scientific Reports, 2016(6):27259-27259.
[49]Koch R A, Wilson A W, Séné O, et al.. Resolved Phylogeny and Biogeography of the Root Pathogen Armillaria and its Gasteroid Relative, Guyanagaster[J]. Bmc Evolutionary Biology, 2017,17(1):33-33.
[50]Chen LY, Chen JM, Gituru RW, et al. Eurasian Origin of Alismatidae Inferred from Statistical Dispersal-vicariance Analysis[J]. Molecular Phylogenetics and Evolution, 2013(67): 38-42.
[51]Burke JM, Bayly MJ, Adams PB, et al. Molecular Phylogenetic Analysis ofDendrobium(Orchidaceae), with Emphasis on the Australian SectionDendrocoryne, and Implications for Generic Classification[J]. Australian Systematic Botany, 2008(21): 1-14.
[52]Vinnersten A, Bremer K. Age and Biogeography of Major Clades in Liliales[J]. American Journal of America, 2001(88): 1695-1703.
[53] Br.-Bl. Coloured Illustrations of Herbaceous Plants of Japan (Monocotyledoneae), by Kitamura S.; Murata G.; Koyama T.[J]. Vegetatio, 1966(5):289.
[54]Toda M, Nishida M, Matsui M, et al. Allozyme Variation Among East Asian Populations of the Indian Rice Frog Rana Limnocharis, (Amphibia: Anura)[J]. Biochemical Systematics & Ecology, 1997, 25(2):143-159.
[55]Lin SM, Chen CA, Lue KY. Molecular Phylogeny and Biogeography of the Grass Lizards GenusTakydromus(Reptilia: Lacertidae) of East Asia[J]. Molecular Phylogenetics and Evolution, 2002(22): 276-288.
ResearchProgressofPlantResourcesofAmanaHondaandItsMolecularSystematics
CHEN Shaotong1,2, LIU Feng1,2, SONG Xiangwen2,3, ZHAO Qun2,3, HAN Bangxing2,3
( 1.AnhuiUniversityofChineseMedicine,Hefei230012,China;2.WestAnhuiUniversity,Lu’an237012,China;3.SynergeticInnovationCenterofDendrobiaIndustrializationDevelopmentinAnhuiProvince,Lu’an237012,China)
The genus Amana Honda belongs to the tribe Lilieae (Liliales: Liliaceae) and it is a group confined to East Asia. At present, there are relatively few studies on the Amana Honda, and more concentrated on the chemical composition, tissue culture and biological characteristics of Amana edulis (Miq.) Honda, and few reports on other species in this genus. In this paper, we reviewed the advances in the phylogenetic position, the taxonomic status of the genus, biological characteristics and molecular systematics of the genus. Based on the existing research and the preliminary research the group, and the key scientific problems existing in this genus, it is of great significance for the development, utilization and protection of Amana Honda medicinal plant resources.
Amana edulis (Miq.) Honda; Amana Honda; plant resources; biological characteristics; molecular systematics
R284
A
1009-9735(2017)05-0102-07
2017-09-12
安徽省自然科学基金(1508085MH203);中央本级重大增减支项目(2060302);中央财政林业科技推广示范项目(2016TG05);安徽高校自然科学研究项目(KJ2016SD61);现代农业产业技术体系建设专项(CARS-21)。
陈绍通(1992-),女,河南原阳人,硕士研究生,研究方向:中药资源开发与利用;通信作者:韩邦兴(1978-),男,安徽枞阳人,硕士生导师,教授,研究方向:中药资源。
