芒针对促进大鼠急性脊髓损伤功能修复的机制研究
2017-10-13李长明谢尚举全仁夫王拓杜伟斌杨宗保
李长明,谢尚举,全仁夫,王拓,杜伟斌,杨宗保
芒针对促进大鼠急性脊髓损伤功能修复的机制研究
李长明1,谢尚举1,全仁夫1,王拓1,杜伟斌1,杨宗保2
(1.杭州市萧山区中医院,杭州 311201;2.厦门大学医学院,厦门 361102)
目的 观察芒针对急性脊髓损伤炎症反应和神经细胞凋亡的影响,研究PI3K/Akt和MAPK/ERK信号转导途径是否参与芒针的神经保护作用,探讨芒针促脊髓损伤修复的具体作用机制。方法 将150只成年雄性SD大鼠随机分为A组、B组、C组、D组和E组,每组30只。采用改良Allen’s法制作大鼠脊髓中度损伤模型,A组为假手术组,不予以损伤脊髓及芒针治疗;B组造模后不予以芒针治疗;C组造模后采用芒针治疗;D组造模前0.5 h鞘内注射LY294002,造模后采用芒针治疗;E组造模前0.5 h鞘内注射PD98059,造模后采用芒针治疗。分别采用BBB评分检测大鼠的自发活动;ELISA检测炎性因子TNF-a、IL-6、IL-1b、NF-kB的含量;TUNEL检测细胞凋亡的程度;免疫组化检测Bcl-2和Bax阳性细胞水平;Western印迹分析p-Akt和p-ERK在脊髓组织的表达,RT-PCR分析Cyt-C和Caspase-3在体内的表达。相对的下行Akt和ERK信号通路通过LY294002和PD98059特异性抑制剂处理分析在体内的抑制作用。结果 炎症反应和PI3K/Akt和MAPK/ERK信号通路抑制的神经元凋亡参与了脊髓损伤大鼠模型的损害,芒针介导的神经保护作用与Bax蛋白阳性神经元数目的减少及Bcl-2蛋白阳性神经元数量的增加有关。芒针治疗能改善大鼠的运动功能,PI3K/Akt和MAPK/ERK信号通路中p-Akt和p-ERK的激活,通过下调Bax蛋白和Bcl-2表达上调,抑制了线粒体凋亡途径关键因子Cyt-C的表达。TUNEL法检测并抑制激活神经元凋亡的Caspase-3的级联。PI3K/Akt和MAPK/ERK信号通路特异性抑制剂LY294002和PD98059的应用抑制了p-Akt和p-ERK的表达。结论 芒针促脊髓损伤修复的神经保护作用可能与炎症反应和PI3K/Akt和MAPK/ERK信号通路的激活、通过下调Bax蛋白和Bcl-2表达上调以及抑制线粒体途径诱导的凋亡有关。
巨刺疗法;芒针;脊髓损伤;炎症反应;p-Akt;p-ERK;Bcl-2;Bax;Cyt-C;Caspase-3;凋亡;大鼠
脊髓损伤是一种临床常见病,常导致脊髓功能的不可逆转、损伤平面以下运动及感觉功能完全丧失,从而引起终身残疾,给家庭和社会带来了沉重的负担。脊髓损伤后的不可逆损伤是由继发性损害所致,而继发性脊髓损伤是由一系列复杂的细胞和分子过程介导的。
炎症反应和凋亡是继发性损伤的重要组成部分,在调节急性和慢性脊髓损伤[1-2]的发病机制中起核心作用。有研究[3-4]报道,减少炎症反应和细胞凋亡能减轻脊髓损伤后的继发性损伤和促进脊髓功能的恢复。PI3K/Akt和MAPK信号转导途径是中枢神经系统几个关键的促生存通路。有研究[5]表明,中枢神经系统存在大量Akt和ERK1/2,其参与神经组织的各个生理和病理过程。Akt和ERK1/2能通过多种途径促进细胞的存活,增加与Bcl-2/Bax的比例,抑制细胞色素C的释放及Caspase级联反应,产生抗凋亡作用[6-10]。LY294002和PD98059是Akt和ERK1/2的特异性抑制剂,能阻断Akt和ERK1/2信号通路产生促凋亡作用[11-13]。
目前无论中医学还是西医学,仍然没有很好的方法用于治疗脊髓损伤[14]。针灸是中国传统医学的一部分[15],针刺对脊髓的形态结构及功能的影响是多方面的,但至今还未能完全被认知。芒针系由古代“九针”中的“长针”发展而来,具有取穴少、透穴多、针感强、传导快等特点,尤其在治疗病位深、病势急、病因复杂而难愈的顽疾时,能发挥出一般针灸无法达到的疗效。笔者前期研究表明,芒针秩边与水道穴可以明显改善脊髓损伤后尿潴留的情况,使脊髓诱发电位产生变化。然而,芒针对脊髓损伤的具体作用和机制尚不明了。故本实验探讨芒针对脊髓损伤后炎症反应和凋亡的影响,并阐明芒针促脊髓功能修复的PI3K/Akt和MAPK/ERK信号转导途径介导抗凋亡机制,现报告如下。
1 材料与方法
1.1 实验动物
150只健康成年雄性SD大鼠,体重为(250±30)g,清洁级,由厦门大学动物实验中心提供并分笼饲养[SYXK(闽)2013-0006]。所有大鼠饲养于恒温、恒湿环境,每笼3~4只,采用人工照明,12 h光照/黑暗(光照时间7:00—19:00)周期循环自由取食基础饲料和自来水,检疫1星期后分组实验。动物实验经厦门大学实验动物管理委员会批准,并遵循美国国立卫生研究院于1996年修订的《实验动物管理和使用指南》。
1.2 主要试剂与仪器
山羊抗兔磷酸化蛋白激酶B多克隆抗体(P-Akt)、山羊抗兔p-ERK1/2多克隆抗体、山羊抗兔Bcl-2多克隆抗体、山羊抗兔Bax多克隆抗体和山羊抗兔Bad多克隆抗体(Bioword公司,中国);LY294002和PD98059 (Sigma公司,美国);TNF-a酶联免疫吸附测定(ELISA)试剂盒、IL-1b酶联免疫吸附测定(ELISA)试剂盒、IL-6联免疫吸附测定(ELISA)试剂盒和NF-kB酶联免疫吸附测定(ELISA)试剂盒(TSZ公司,美国);原位末端转移酶标记法(TUNEL)试剂盒(罗氏生物公司,北京);JL2B型电脉冲刺激仪(苏州医疗用品厂有限公司,江苏);RM2135型切片机(Leica公司,德国);电泳及转膜仪(Bio-Rad公司,美国)。
1.3 大鼠脊髓损伤模型建立
SD大鼠造模前禁食8 h,实验顺序随机进行。大鼠称重后用10%水合氯醛(0.3 mL/100 g)腹腔注射麻醉,用卵圆钳钳夹无四肢反应后将大鼠俯卧位固定于操作台上,采用改良Allens法制作脊髓中度损伤SD大鼠模型。胸背部(T8-12)正中切口,钝性分离椎旁肌和棘上韧带,暴露上下各1个锥体长度,咬除T9-10棘突及全部椎板,暴露0.5 cm宽硬膜。用质量为10 g的克氏针沿带有刻度的导管从60 mm高处垂直自由下落,打击在直径4 mm、宽2 mm的由薄塑料材料制成的半圆片上,致伤后迅速移开打击物,造成大鼠脊髓后角中度损伤,采用4-0丝线逐层缝合。整个实验过程在(37±0.5)℃的温度下进行,术后皮下注射1.0 mL生理盐水以补充丢失的血容量及每日青霉素(80 U/d)腹腔注射预防感染,单笼饲养,室温保持在20℃~25℃,给予充足的食物和水。每日2次膀胱按摩协助排尿,直至建立反射性膀胱排空。
1.4 动物分组及处理
将150只大鼠随机分为A组、B组、C组、D组和E组,每组30只。A组只打开椎板,不予以损伤脊髓及芒针治疗;B组造模后,不予以芒针治疗;C组造模后,采用芒针治疗,每日1次,共治疗7次;D组术前0.5 h鞘内注射5mg LY294002 10mL(溶于1%DMSO),造模后行芒针治疗,每日1次,共治疗7次;E组术前0.5 h鞘内注射20mmol/L PD98059 10mL(溶于1%DMSO),造模后行芒针治疗,每日1次,共治疗7次。
1.5 芒针治疗方法
C组、D组和E组造模成功后,待大鼠麻醉苏醒后再进行芒针治疗。取秩边、水道穴。穴位定位根据世界卫生组织制定的国际标准。秩边位于臀外下部,股骨大转子与荐椎尾椎结合部连线外1/3与中1/3交点处;水道位于腹部,耻骨联合与胸剑联合中点连线(分13等份)耻骨联合上3等份旁开约2 cm处。详见图1。针刺部位备皮消毒,采用华佗牌0.25 mm×25 mm不锈钢芒针直刺14~15 mm,捻转1 min后留针15 min,其间同侧秩边、水道(每次单侧,左右交替)连接JL2B型电脉冲刺激仪,频率为2 Hz,电流为1~3 mA,共治疗15 min。每日1次,共治疗7次。
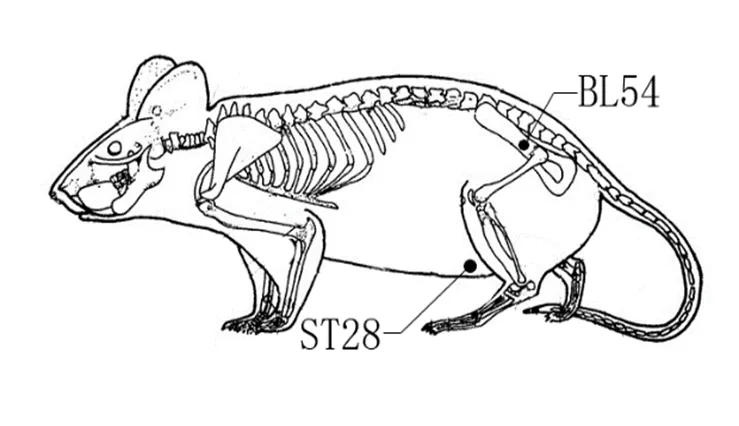
图1 大鼠穴位示意图
1.6 实验取材
每组取10只大鼠分别于术后即刻及术后1 d、3 d、7 d、14 d进行运动功能评分;每组取10只大鼠于术后7 d心脏灌注取受损节段脊髓组织1 cm用于HE、TUNEL和免疫组化检测;每组取10只大鼠于术后7 d在冰上取新鲜段受损节段脊髓组织1 cm用于ELISA、Western印迹和RT-PCR检测。
1.7 检测指标与方法
1.7.1 功能评价
采用Basso-Beattie-Bresnahan(BBB)评分法对大鼠进行功能评价。在本次研究中,笔者对大鼠特定的功能行为进行了评价,如肢体运动、爪掌面、步态协调、脚趾间隙和尾部的位置等。如果没有自发的后肢运动得分为0分,21分表示正常运动。评价时由两位经验丰富且独立的测试者在1个100 cm×100 cm的开放区域内进行,共测试4 min。
1.7.2 光镜
脊髓损伤后7 d,各组大鼠给予10%的水合氯醛(0.3 mL/100 g)和过量肝素生理盐水心脏灌注20 min,继之再用0.1 mol PBS配置的4%多聚甲醛固定液(pH值=7.4)灌注5 min,取受损节段脊髓组织1 cm石蜡包埋,切成纵向6mm厚的石蜡切片,组织片二甲苯脱蜡,加入1X苏木素染液/溶性伊红染液,中性树胶封固,再使用光学显微镜进行分析。
1.7.3 细胞因子的测定
取T9-10节段脊髓组织匀浆提取上清液,取提取的上清液及不同浓度的标准品各孔50mL,用封板膜封板后置37℃温育30 min,洗涤5次,每孔加入酶标试剂50mL,再温育、洗涤、显色,在酶标仪上于450 nm波长处以阴性对照孔调零,检测各孔光密度(OD)值,根据标准液的检测结果得出标准曲线,求出各组样品的实际浓度。
1.7.4 TUNEL检测
切片二甲苯脱蜡和再水化,将切片用Proteinase K工作液处理20 min,然后用PBS洗涤3次,加50mL TUNEL反应混合液于标本上(1号试剂2号试剂以1:14混合),加盖玻片或封口膜在暗湿盒中(37℃ 1 h)。然后用PBS洗涤,滴加50mL 3号试剂(37℃ 30 min),再用PBS洗涤后,滴加100mL新鲜配制的DAB工作液,显微镜下观察随时终止反应。采用苏木素进行复染,中性树胶封固。每张切片随机取5个视野计数TUNEL阳性和阴性细胞,结果以TUNEL阳性细胞数占总细胞的百分比表示。
1.7.5 Bcl-2和Bax的免疫组化定位
石蜡包埋制备6mm厚的组织切片,然后二甲苯脱蜡和再水化,经微波抗原修复,每张切片加1滴或50mL的3%过氧化酶阻断剂,以阻断内源性过氧化物酶的活性,室温下孵育10 min;加1滴或50mL正常非免疫血清,37℃孵育10 min;加50mL的第一抗体(bcl-2,1:40稀释,bs1511;bax,1:20稀释,bs1030);加50mL生物素标记的山羊抗兔IgG(1:200稀释,B-0061),37℃孵育40 min;加50mL辣根过氧化酶标记链亲和素溶液(1:400倍稀释,IH-0061),37℃孵育40 min;加100mL新鲜配制的DAB工作液(AR-0611),显微镜下观察随时终止反应。每一步期间进行0.01 M PBS浸泡,每次5 min,共3次。苏木素复染,中性树胶封固。染色切片用成像软件系统和显微镜检测免疫阳性细胞。
1.7.6 免疫印迹检测p-Akt和p-ERK1/2
总蛋白用总蛋白提取试剂盒提取分离。蛋白浓度使用BCA蛋白定量试剂盒的说明书进行测定。受损节段脊髓样品50~100mg加入相同体积的5×loading buffer上样缓冲液,煮沸10 min。进行SDS-聚丙烯酰胺凝胶电泳,转移至PVDF膜上,室温下孵育兔抗大鼠p-Akt蛋白一抗(1:1000);兔抗大鼠p-ERK1/2蛋白一抗(1:1000),再与辣根过氧化物酶标记的抗兔IgG抗体(1:4000)在室温下孵育,将膜与ECL发光底物,膜与胶片在暗室结合曝光、显影、成像。经Quantity One图像分析软件进行光密度分析,目的条带的光密度与相应的内参蛋白b-actin条带的光密度的比值即为目的蛋白表达的相对表达值。
1.7.7 RT-PCR检测Cyt-C和Caspase-3
总RNA采用RNA自旋试剂盒的说明书进行抽提。反转录体系含4mL DNAse 处理过的RNA、5×BuffermL、1mL dNTPs(10 mM)、1mL Oligo(dT)18 (50mM)、0.5mL随机引物(100mM)、1mL MMLV逆转录酶(200 U/mL)、8.5mL DEPC H2O,共20mL。然后按以下条件进行反转录反应。30℃ 10 min,42℃ 60 min,99℃ 5 min,4℃ 5 min。取出后在0.2 mL PCR管内,依次加入1mL反转录产物(RT产物),各基因上游与下游引物各0.25mL,12.5mL PCR反应mix(Taq酶,Taq buffer, dNTPs),用ddH2O补足体积至25mL。按以下条件进行反转录反应,94℃预变性2 min,94℃变性30 s,56℃/55℃/48℃退火30 s,72℃延伸30 s, 72℃延伸10 min,扩增35个循环。用one-Dscan图像分析软件分析目标条带的灰度值。
从GenBank获得目的基因mRNA的全长序列,利用引物和探针设计软件Primer 5.0设计引物序列。经过Blast分析,引物序列具有特异性。所用的引物及内参照b-actin引物见表1。引物均委托鼎国昌盛生物技术有限责任公司合成。

表1 目的基因的引物信息
1.8 统计学方法
所有数据采用SPSS16.0软件进行统计分析。计量资料以均数±标准差表示,组间比较若满足正态性且方差齐时采用单因素方差分析法和法,方差不齐时选择和法进行方差检验和两两比较,若不满足正态性时采用秩和检验。
2 结果
2.1 功能评估
本实验共用大鼠150只,存活139只,存活率为92.7%,死亡大鼠予以补齐。A组后肢的运动功能(BBB评分)在术后3 d恢复到了21分,且A组造模后各时间点BBB评分与B组比较,差异均具有统计学意义 (<0.01)。B组和C组在术后1 d内BBB评分<2分,随时间的延长,后肢功能逐渐恢复,BBB评分逐渐升高,但两组均未达到术前水平。C组治疗后1、3 d BBB评分与B组比较,差异均无统计学意义(>0.05)。C组治疗后7、14 d BBB评分与B组比较,差异均有统计学意义(<0.05),提示芒针治疗能促进大鼠脊髓功能的恢复,治疗后7 d芒针的疗效开始显现。详见表2。
表2 各组不同时间BBB评分比较 (±s,分)

表2 各组不同时间BBB评分比较 (±s,分)
组别n术后即刻术后1 d术后3 d术后7 d术后14 d A组3020.20±0.8420.40±0.5521.00±0.0021.00±0.0021.00±0.00 B组30 0.20±0.451) 0.60±0.551) 2.20±0.451) 3.80±0.451) 5.20±0.451) C组30 0.20±0.45 0.40±0.84 2.80±0.84 7.40±0.552) 11.60±0.892)
注:与A组比较1)<0.01;与B组比较2)<0.05
2.2 光镜
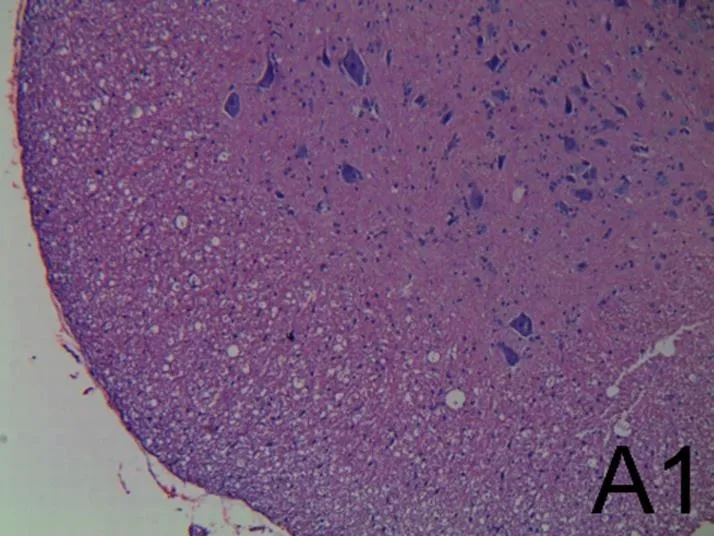
术后7 d,光镜下检查A组(见图2A1、图2A2)脊髓形态结构完整,灰质神经细胞形态正常,分布均匀,细胞膜、细胞核及组织间隙均正常,白质纤维分布均匀,髓鞘形态完整,排列整齐。B组(见图2B1、图2B2)脊髓形态不完整,神经组织残缺;组织重度出血,存在大量血细胞;组织疏松水肿,细胞空泡变性,部分细胞核固缩,神经纤维溶解、消失;灰质神经细胞数量减少,灰质神经细胞肿胀、核裂解甚至消失,细胞周围基质消失呈空泡状;白质纤维减少,分布不均匀,脱髓鞘,形态不完整、相互融合。C组(见图2C1、图2C2)脊髓形态结构基本完整,组织中散在血细胞,组织无水肿;灰质神经细胞形态基本正常,空泡变性减轻,部分细胞仍存在空泡变性,细胞核无明显固缩,白质纤维分布基本均匀,形态完整,无脱髓鞘。提示芒针能有效促进脊髓形态的恢复,效果优于B组。
2.3 抗炎反应
为了测试芒针治疗是否通过调节炎性细胞因子的分泌调控炎症过程,笔者分析了T9-10节段脊髓组织中的TNF-a、IL-1b、IL-6、NF-kB的水平。由表3可见,脊髓损伤7 d后,B组促炎细胞因子(TNF-a、IL-1b、IL-6、NF-kB)的水平均显著升高,与A组比较,差异均具有统计学意义(<0.01)。芒针治疗后,C组促炎细胞因子(TNF-a、IL-1b、IL-6、NF-kB)的水平均显著下降,与B组比较,差异均具有统计学意义(<0.01)。提示芒针能通过抑制炎症反应促进脊髓损伤的恢复。
表3 各组大鼠脊髓组织中TNF-a、IL-1b、IL-6、NF-Kb水平比较 (±s,ng/L)
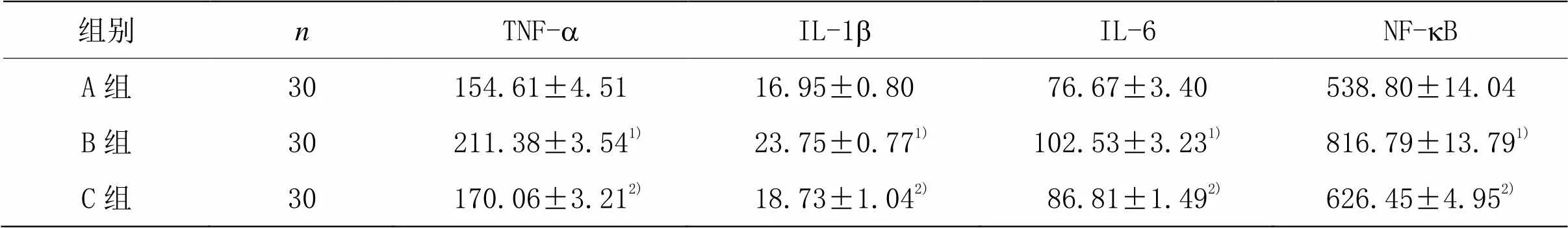
表3 各组大鼠脊髓组织中TNF-a、IL-1b、IL-6、NF-Kb水平比较 (±s,ng/L)
组别nTNF-aIL-1bIL-6NF-kB A组30154.61±4.5116.95±0.8076.67±3.40538.80±14.04 B组30 211.38±3.541) 23.75±0.771)102.53±3.231) 816.79±13.791) C组30 170.06±3.212) 18.73±1.042) 86.81±1.492)626.45±4.952)
注:与A组比较1)<0.01;与B组比较2)<0.01
2.4 抗细胞凋亡的影响
为了检验芒针是否能有效抑制细胞凋亡,笔者采用损伤脊髓组织(距离T10脊髓节段损伤正中0.1 cm)进行TUNEL染色观察。A组因未损伤脊髓,仅有少量凋亡(见图3A)。术后7 d,B组大鼠脊髓损伤节段周围出现大量的凋亡细胞(见图3B),大鼠脊髓组织神经细胞凋亡指数与A组比较,差异有统计学意义(<0.01)。C组经芒针治疗后,凋亡细胞数明显减少(见图3C),神经细胞凋亡指数与B组比较,差异具有统计学意义 (<0.01)。D组和E组分别使用LY294002和PD98059后,凋亡细胞数有所回升(见图3D、图3E),神经细胞凋亡指数与C组比较,差异均具有统计学意义(<0.01)。每张切片选取5~10个不同的视野进行TUNEL阳性细胞计数,阳性细胞率=阳性细胞数/总细胞数。提示脊髓损伤后存在大量凋亡的发生,芒针治疗后减少了凋亡现象,使用LY294002和PD98059后抑制了芒针的作用,说明PI3K/Akt和MAPK/1/2信号通路可能参与了芒针修复脊髓损伤的过程。详见表4。
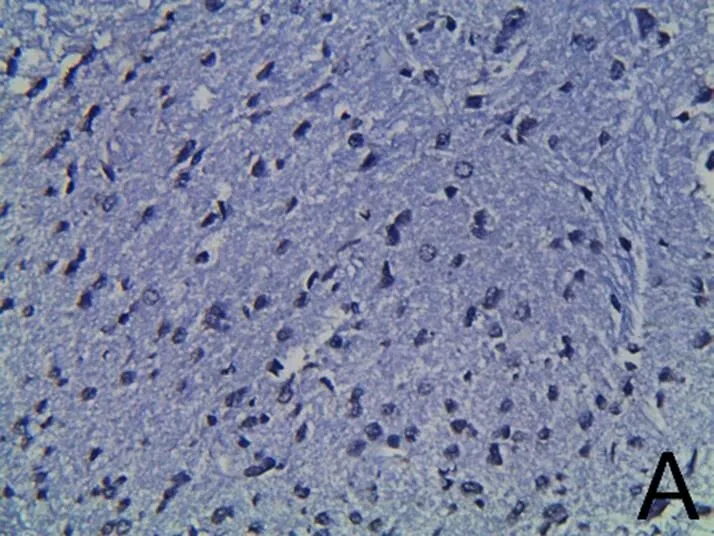
表4 各组脊髓组织神经细胞凋亡指数比较 (±s)

表4 各组脊髓组织神经细胞凋亡指数比较 (±s)
组别n神经细胞凋亡指数 A组300.08±0.02 B组30 0.95±0.021) C组30 0.69±0.062) D组30 0.89±0.023) E组30 0.88±0.043)
注:与A组比较1)<0.01;与B组比较2)<0.01;与C组比较3)<0.01
2.5 Bax和Bcl-2免疫反应的分布及其变化
表5 各组脊髓组织Bcl-2、Bax阳性表达情况比较(±s)
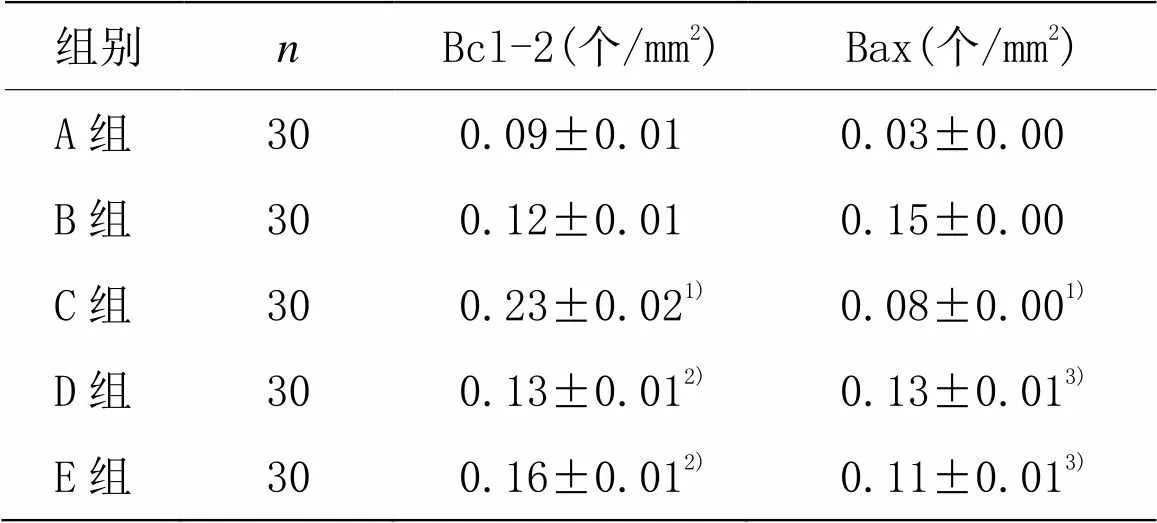
表5 各组脊髓组织Bcl-2、Bax阳性表达情况比较(±s)
组别n Bcl-2(个/mm2) Bax(个/mm2) A组300.09±0.010.03±0.00 B组300.12±0.010.15±0.00 C组30 0.23±0.021) 0.08±0.001) D组30 0.13±0.012) 0.13±0.013) E组30 0.16±0.012) 0.11±0.013)
注:与B组比较1)<0.01;与C组比较2)<0.01,3)<0.05
脊髓损伤后7 d,B组Bcl-2的阳性细胞数明显减少(见图4B1),Bax的阳性细胞数明显增加(见图4B2)。经芒针治疗后,C组Bcl-2的阳性细胞数明显增加(见图4C1),Bax的阳性细胞数明显减少(见图4C2),与B组比较,差异均有统计学意义(<0.01)。D组Bcl-2的阳性细胞数有所下降(见图4D1、图4E1),Bax的阳性细胞数有所增加(见图4D2、图4E2),与芒针组相比,差异均具有统计学意义(<0.01,<0.05)。提示芒针可以通过上调Bcl-2和下调Bax的表达产生抗凋亡作用。详见表5。

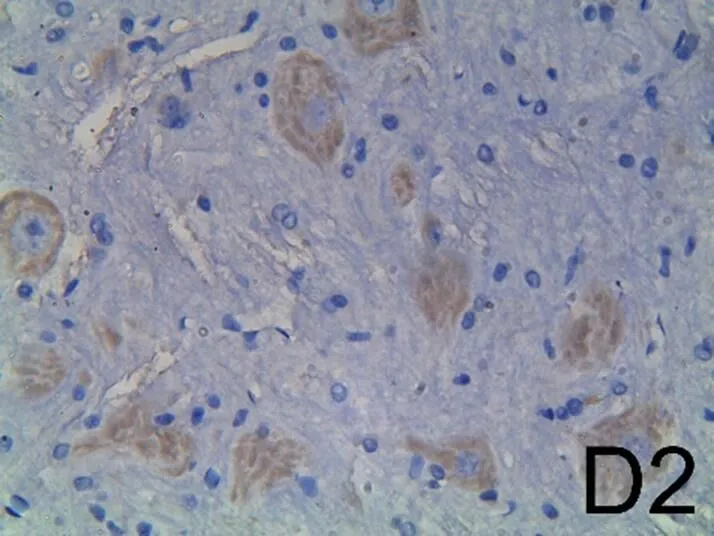
注:A1为A组Bcl-2,A2为A组Bax,B1为B组Bcl-2,B2为B组Bax,C1为C组Bcl-2,C2为C组Bax,D1为D组Bcl-2,D2为D组Bax,E1为E组Bcl-2,E2为E组Bax
2.6 PI3K/Akt和ERK1/2信号通路的影响
信号转导过程的激活,可以影响疾病的转归。为了检验PI3K/Akt和ERK1/2信号通路是否参与了芒针治疗的过程,笔者采用新鲜脊髓组织(距离T10脊髓节段损伤正中0.1 cm)进行Western Blot检测。结果显示, B组脊髓损伤后p-Akt和p-ERK1/2表达明显升高,与A组比较,差异均具有统计学意义(<0.01)。C组p-Akt和p-ERK1/2显著增高,与B组比较,差异均有统计学意义(<0.01)。D组和E组p-Akt和p-ERK1/2表达均降低,与C组比较,差异均有统计学意义(<0.05,<0.01)。提示芒针治疗可能与PI3K/Akt和ERK1/2信号通路有关。详见表6及图5、图6。
表6 各组受损节段脊髓组织p-Akt和p-ERK1/2表达情况比较 (±s,ng/L)
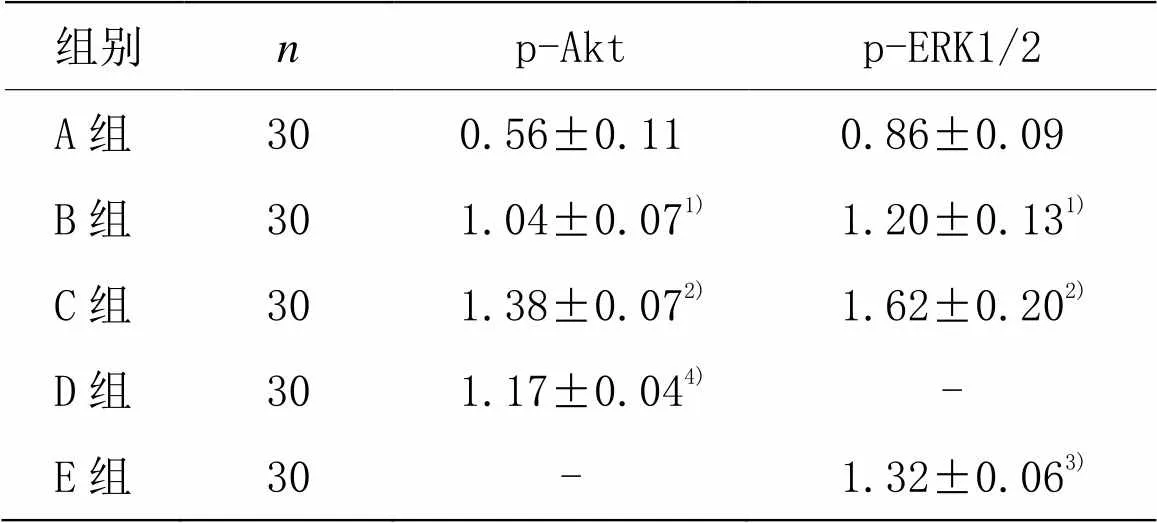
表6 各组受损节段脊髓组织p-Akt和p-ERK1/2表达情况比较 (±s,ng/L)
组别np-Aktp-ERK1/2 A组300.56±0.110.86±0.09 B组30 1.04±0.071) 1.20±0.131) C组30 1.38±0.072) 1.62±0.202) D组30 1.17±0.044)- E组30- 1.32±0.063)
注:与A组比较1)<0.01;与B组比较2)<0.01;与C组比较3)<0.01,4)<0.05

图5 术后7 d各组受损节段脊髓组织p-Akt表达情况
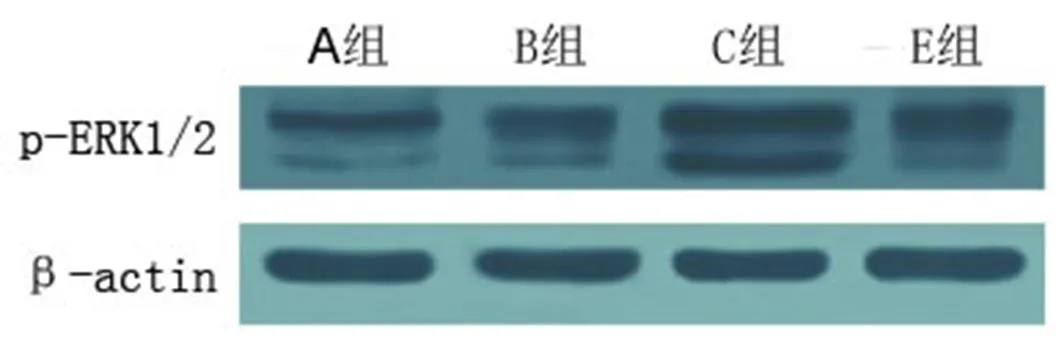
图6 术后7 d各组受损节段脊髓组织p-ERK1/2表达情况
2.7 线粒体凋亡途径的影响
表7 各组受损节段脊髓组织Cyt-C和Caspase-3的mRNA含量比较 (±s,ng/L)

表7 各组受损节段脊髓组织Cyt-C和Caspase-3的mRNA含量比较 (±s,ng/L)
组别nCyt-CCaspase-3 A组301.00±0.020.91±0.12 B组30 1.45±0.031) 1.28±0.031) C组30 1.04±0.082) 0.95±0.051) D组30 1.21±0.093) 1.10±0.063) E组30 1.31±0.023) 1.17±0.023)
注:与A组比较1)<0.01;与B组比较2)<0.01;与C组比较3)<0.01
A组因未损伤脊髓,Cyt-C和Caspase-3仅少量表达。B组脊髓损伤后促进了Cyt-C和Caspase-3的激活,与A组比较,差异具有统计学意义(<0.01)。C组显著抑制Cyt-C和Caspase-3的表达,与B组比较,差异有统计学意义(<0.01)。D组和E组Cyt-C和Caspase-3的表达均有所升高,与C组比较,差异均有统计学意义(<0.01,<0.05)。提示芒针治疗后可以促进Cyt-C和Caspase-3的降低,这种作用可能与PI3K/Akt和ERK1/2信号通路有关。详见表7及图7、图8。
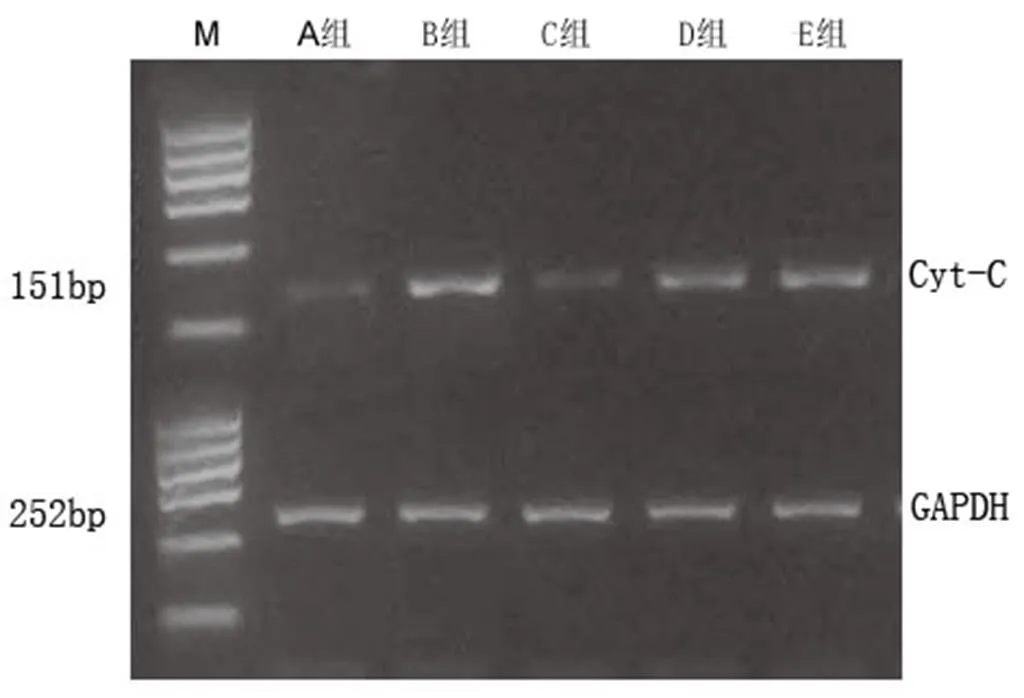
图7 术后7 d各组大鼠受损节段脊髓组织Cyt-C的mRNA含量表达情况
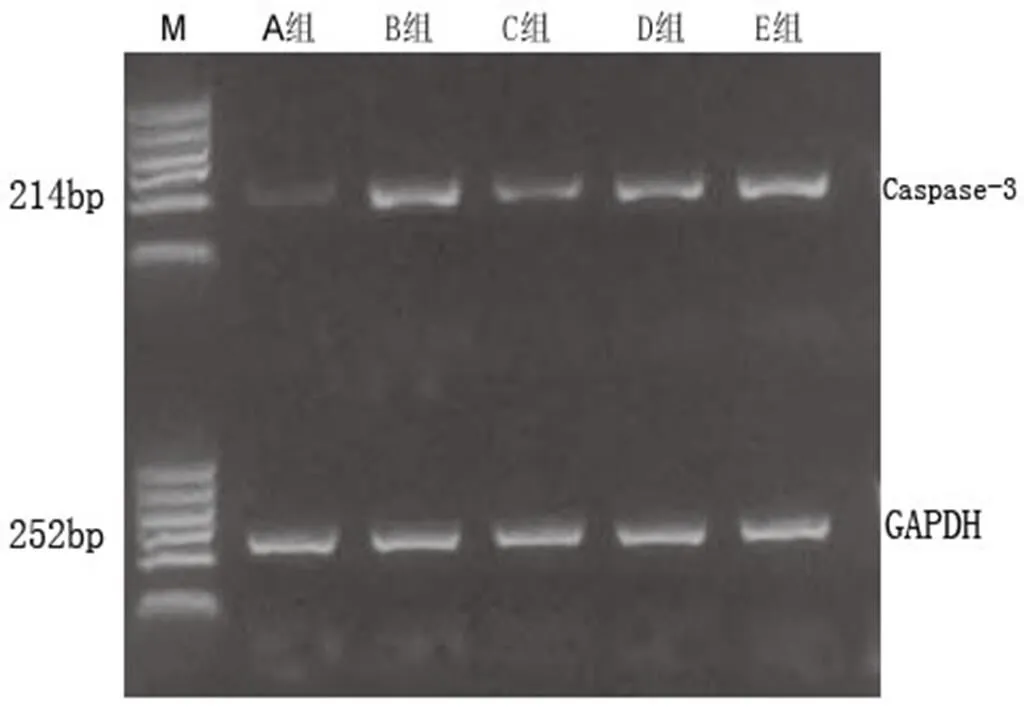
图8 术后7 d各组大鼠受损节段脊髓组织Caspase-3的mRNA含量表达情况
3 讨论
芒针由古代九针之一的“长针”发展而来,因针体长、刺入深,具有取穴少、透穴多、针感强、传导快等特点,被广泛应用于各种疾病的治疗中,包括神经系统疾病中的神经根炎、多发性神经炎、瘫痪、胃肠疾病以及运动系统、精神系统、妇科等方面的疾患[16-17]。本研究结果显示,①芒针促进了脊髓损伤的功能性恢复;②炎症反应和细胞凋亡参与了芒针的治疗作用;③进一步研究发现,PI3K/Akt和ERK1/2信号通路参与了芒针抗凋亡作用;④PI3K/Akt和ERK1/2信号通路的激活是通过线粒体凋亡途径产生抗凋亡作用实现的。
脊髓损伤的治疗一直是神经科学研究的一大问题。长期大量的研究证明,针刺治疗急性脊髓损伤可以减轻炎症反应和脊髓继发性损伤,改善感觉和运动功能,并改善晚期神经功能障碍[18-19]。本研究结果显示,芒针治疗7 d后,损伤大鼠BBB评分及形态学观察均有好转,运动功能恢复明显,脊髓形态结构尚完整。在此基础上,笔者认为芒针治疗急性脊髓损伤是有疗效的,其参与了脊髓损伤的神经保护作用。
众所周知,免疫炎症反应是脊髓继发性损伤的主要机制之一。小胶质细胞在继发性神经损伤发挥重要的作用。激活的小胶质细胞会释放TNF-a、IL-1b、IL-6等促炎性因子,从而加重局部炎症反应。TNF-a通过与肿瘤坏死因子受体(TNFR)结合,活化积聚炎症细胞,激活内皮细胞及神经细胞,促进NF-kB的活化及IL-1b、IL-6、TNF-a自身的分泌,释放细胞毒素(如氧自由基、蛋白水解酶等),诱导花生四烯酸代谢产物的释放及脂质过氧化,破坏细胞膜性结构,导致炎症加重,增加血脊髓屏障的通透性,使损伤的脊髓进一步损伤,并可通过多种途径诱发凋亡[20]。大量研究[21-24]证明,针灸具有抗炎、镇痛、抗脂质过氧化、防止细胞受损、抗凋亡等作用。本研究结果发现,脊髓损伤后,大鼠NF-kB、TNF-a、IL-1b、IL-6等促炎性因子含量和神经元细胞凋亡率明显增高,经芒针治疗可明显降低伤段脊髓组织中NF-kB、IL-6b、IL-1b、TNF-a的浓度,并使神经元细胞凋亡率明显下降,具有明显的炎症抑制和抗凋亡作用。
细胞凋亡是神经细胞损伤后细胞程序性死亡的重要形式。处于细胞凋亡的早期阶段的信号转导过程的激活,是细胞凋亡发生的必要前提。PI3K/Akt和ERK1/2信号通路的活化是调节细胞的增殖、分化、代谢、凋亡的主要途径[25-26]。活化的Akt(p-Akt)被认为是调节神经元生存的中枢性控制因子,与神经元的发育、分化、轴突的生长以及介导突触的可塑性等密切相关。Akt的凋亡抑制作用是由Yao R等[27]首次发现的,其研究表明抑制PI3K/Akt的活性能抵消神经生长因子(NGF)抑制细胞凋亡促细胞存活的作用。Jung SY等[7]在研究跑步机运动通过激活PI3K/Akt通路在大鼠降低脊髓损伤诱导的细胞凋亡中发现脊髓损伤后降低了NT-3和IGF-1蛋白的表达,p-Akt/Akt的比率与Bcl-2/Bax的比例明显下降,Caspase-3的表达明显增加。这项研究表明在脊髓损伤后跑步机上锻炼通过抑制细胞凋亡促进运动功能的恢复,且锻炼的有益效果可以归因于通过激活PI3K/Akt途径的增加的神经营养因子表达的影响。ERK1/2细胞信号传导通路是由一个小GTP蛋白连接活化的受体酪氨酸激酶和胞浆蛋白组成的级联反应[28],使ERK在神经损伤的区域迅速激活(p-ERK1/2),减少神经损伤对机体带来的影响。在外伤性脑损伤中,急性挫伤区的神经元可发生凋亡,并且持续数天至数星期,此过程中ERK1/2与Caspase家族蛋白被激活,引起兴奋性氨基酸增多,使细胞内的钙离子和氧自由基增加,促进了神经元的凋亡作用[29]。本研究也显示,芒针治疗后,大鼠p-Akt、p-ERK1/2的表达明显增强,说明PI3K/Akt和ERK1/2信号通路有可能参与了芒针的抗凋亡作用。LY294002和PD98059分别为PI3K/Akt和ERK1/2信号通路的特异性抑制剂,特异性抑制Akt和ERK1/2的磷酸化,因而阻断信号通路下游的级联反应。有研究[11]发现,帕金森病中促红细胞生成素的抗凋亡作用是通过PI3K/Akt/GSK-3b/ Caspase-3信号通路实现的,而这种抗凋亡作用可以被PI3K抑制剂LY294002和GSK-3b抑制剂氯化锂抵消。Lu KT等[13]通过使用ERK/MAPK通路特异性抑制剂PD98059,发现抑制ERK/MAPK通路的激活可以减轻大鼠TBI模型引起的脑组织水肿和神经功能损害。本实验在脊髓损伤前0.5 h通过鞘内注射LY294002和PD98059,并进行芒针治疗后发现,注射过特异性抑制剂LY294002和PD98059的大鼠p-Akt和p-ERK1/2表达较芒针组少,差异有统计学意义(<0.05,<0.01)。因此验证了PI3K/Akt和ERK1/2信号通路确实参与了芒针的抗凋亡作用。
线粒体是细胞的“能量供应器”,不仅参与了多种生理过程的调节,还通过线粒体凋亡途径在细胞凋亡中发挥重要作用[30]。当细胞受损时,线粒体膜的通透性改变,将会释放Cyt-C和Caspase,通过信号级联激活细胞死亡程序。细胞色素C是线粒体介导凋亡途径的关键因子。Akt可阻止线粒体释放细胞色素C及凋亡诱导因子AIF,抑制Caspase瀑布级联反应,从而发挥抗凋亡的作用。Akt还能磷酸化Bcl-2家族成员BAX的Ser184位点,抑制Cyt-C释放,负调控促凋亡功能[31]。ERK1/2的激活不但可以上调Bcl-2等抗凋亡蛋白的表达,而且可以诱导其活化[32-33]。另外,活化的ERK1/2能使Bad ser112位点磷酸化(p-Bad),p-Bad与Bcl-2或Bcl-xL发生解聚,并且与磷酸丝氨酸结合蛋白14-3-3结合,游离的Bcl-2或Bcl-xL则促进细胞的抗凋亡作用。Luchetti F等[10]发现褪黑激素能够通过提高bcl-2水平降低超氧阴离子的产生、线粒体损伤和Caspase依赖的细胞凋亡,并在细胞质减少Cyt-C的释放,且增加了ERK的激活,ERK1/2抑制剂PD98059的应用,ROS增加、线粒体功能障碍和细胞凋亡。本研究采用免疫组化检测Bcl-2和Bax的阳性细胞数,发现芒针治疗后Bcl-2阳性细胞数增加,Bax阳性细胞数减少。Cyt-C和Caspase-3在芒针干预后也显著降低,凋亡受到抑制。特异性抑制剂LY294002和PD98059预处理后,Bcl-2阳性细胞数下调,Bax阳性细胞数上调。Cyt-C和Caspase-3在芒针干预后较芒针组有所升高,凋亡较明显。证实了芒针治疗激活PI3K/Akt和ERK1/2信号通路通过线粒体凋亡途径产生抗凋亡作用的。
综上所述,芒针治疗通过炎症抑制和PI3K/Akt及ERK1/2信号通路通过线粒体凋亡途径产生的抗凋亡作用,介导神经保护作用及改善神经运动功能,明确了芒针治疗脊髓损伤的具体机制,为临床应用芒针治疗急性脊髓损伤提供了理论依据。
[1] Fitch MT, Silver J. CNS injury, glial scars, and infl- amemation: Inhibitory extracellular matrices and regene- ration failure[J]., 2008,209(2): 294-301.
[2] Borgens RB, Liu-Snyder P. Understanding secondary injury[J]., 2012,87(2):89-127.
[3] Ferguson AR, Hook MA, Garcia G,. A simple post hoc transformation that improves the metric properties of the BBB scale for rats with moderate to severe spinal cord injury[J]., 2004,21(11):1601-1613.
[4] Lan WB, Lin JH, Chen XW,. Overexpressing neuro- globin improves functional recovery by inhibiting neuronal apoptosis after spinal cord injury[J]., 2014,1562:100-108.
[5] Fukunaga K, Miyamoto E. Role of MAP kinase in neurons[J]., 1998,16(1):79-95.
[6] Shehadah A, Chen J, Zacharek A,. Niaspan treatment induces neuroprotection after stroke[J]., 2010,40(1):277-283.
[7] Jung SY, Kim DY, Yune TY,. Treadmill exercise reduces spinal cord injury-induced apoptosis by activating the PI3K/Akt pathway in rats[J]., 2014,7 (3):587-593.
[8] Raghupathi R, Muir JK, Fulp CT,. Acute activation of mitogen-activated protein kinases following traumatic brain injury in the rat: implications for posttraumatic cell death[J]., 2003,183(2):438-448.
[9] Zhuang S, Schnellmann RG. A death-promoting role for extracellular signal-regulated kinase[J]., 2006,319(3):991-997.
[10] Luchetti F, Betti M, Canonico B,. ERK MAPK activation mediates the antiapoptotic signaling of melato- nin in UVB-stressed U937 cells[J]., 2009,46(3):339-351.
[11] Wu Y, Shang Y, Sun S,.Erythropoietin prevents PC12 cells from 1-methyl-4-phenylpyridinium ion-induced apoptosis via the Akt/GSK-3beta/caspase-3 mediated sig- naling pathway[J]., 2007,12 (8):1365-1375.
[12] Wang Y, Gao C1, Zhang Y,. Visfatin stimulates endometrial cancer cell proliferation via activation of PI3K/Akt and MAPK/ERK1/2 signalling pathways[J]., 2016,43(1):168-178.
[13] Lu KT, Cheng NC, Wu CY,. NKCC1-mediated traumatic brain injury-induced brain edema and neuron death via Raf/MEK/MAPK cascade[J]., 2008,36(3):917-922.
[14] Baptiste DC, Fehlings MG. Update on the treatment of spinal cord injury[J]., 2007,161: 217-233.
[15] Park J, Hopwood V, White AR,. Effectiveness of acupuncture for stroke: a systematic review[J]., 2001,248(7):558-563.
[16] 高锡朋.针刺治疗外伤性截瘫[J].中国针灸,1985,5 (5):3-4.
[17] 李观荣,戢良秀,张声泽,等.针刺治疗外伤性截瘫124例疗效观察[J].中医杂志,1985,26(12):34-36.
[18] Rapson LM, Wells N, Pepper J,. Acupuncture as a promising treatment for below-level central neuropathic pain: a retrospective study[J]., 2003, 26(1):21-26.
[19] Paola FA, Arnold M. Acupuncture and spinal cord medi- cine[J]., 2003,26(1):12-20.
[20] Genovese T, Mazzon E, Crisafulli C,. TNF-alphablockage in a mouse model of SCI: evidence for improved outcome[J]., 2008,29(1):32-41.
[21] Lü YL, Zhao LM, Chen HP,. Effect of electro- acupuncture at Jiaji acupoint on anti-inflammation, analgesia and immunoloregulation[J]., 2006,10 (19):171-173.
[22] Usichenko TI. Acupuncture as part of multimodal anal-gesia after caesarean section[J]., 2014,32 (3):297-298.
[23] Xu JN, Xiao D, Ju JH,. Effect of electro-acupuncture on treatment of spinal cord injuries in rats[J]., 2006,10(23):180-182.
[24] Wang F, Gao Z, Li X,. NDRG2 is involved in anti-apoptosis induced by electroacupuncture pre- treatment after focal cerebral ischemia in rats[J]., 2013,35(4):406-414.
[25] Hetman M, Gozdz A. Role of extracellular signal regu- lated kinases 1 and 2 in neuronal survival[J]., 2004,271(11):2050-2055.
[26] Zhu Y, Culmsee C, Klumpp S,. Neuroprotection by transforming growth factor-beta1 involves activation of nuclear factor-kappaB through phosphatidylinositol-3- OH kinase/Akt and mitogen-activated protein kinase- extracellular-signal regulated kinase1,2 signaling path- ways[J]., 2004,123(4):897-906.
[27] Yao R, Cooper GM. Requirement for phosphatidyli-nositol-3 kinase in the prevention of apoptosis by nerve growth factor[J]., 1995,267(5206):2003-2006.
[28] Mazzucchelli C, Brambilla R. Ras-related and MAPK sig- nalling in neuronal plasticity and memory formation[J]., 2000,57(4):604-611.
[29] Raghupathi R, Muir JK, Fulp CT,. Acute activation of mitogen-activated protein kinases following traumatic brain injury in the rat: implications for posttraumatic cell death[J]., 2003,183(2):438-448.
[30] Bleicken S, Wagner C, García-Sáez AJ. Mechanistic diff- erences in the membrane activity of Bax and Bcl-xL correlate with their opposing roles in apoptosis[J]., 2013,104(2):421-431.
[31] Xin M, Deng X. Nicotine inactivation of the proapoptotic function of Bax through phosphorylation[J]., 2005,280(11):10781-10789.
[32] Wiener Z, Band AM, Kallio P,. Oncogenic mutations in intestinal adenomas regulate Bim-mediated apoptosis induced by TGF-b[J]., 2014,111 (21):E2229-E2236.
[33] Irles C, Nava-Kopp AT, Morán J,. Neonatal mater- nal separation up-regulates protein signalling for cell sur- vival in rat hypothalamus[J]., 2014,17(3):275-284.
Mechanism Study of the Promoting Action of Elongated Needle Acupuncture on Functional Repair in Rats with Acute Spinal Cord Injury
-1,-1,-1,1,-1,-2.
1.,311201,; 2.,361102,
Objective To observe the effect of elongated needle acupuncture on inflammatory reactions and apoptosis in acute spinal cord injury, investigate if PI3K/Akt and MAPK/ERK signal transduction pathways are involved in the neuroprotective effect of elongated needle acupuncture and explore the promoting action of elongated needle acupuncture on spinal cord injury repair. Method One hundred and fifty adult male SD rats were randomized into groups A, B, C, D and E, 30 each. A model of moderate spinal cord injury was made by modified Allen's method. Group A received a sham operation without spinal cord injury and no elongated needle acupuncture. Group B did not receive elongated needle acupuncture after model making. Group C received elongated needle acupuncture after model making. Group D received an intrathecal injection of LY294002 at 0.5 hour before model making and elongated needle acupuncture after model making. Group E received an intrathecal injection of PD98059 at 0.5 hour before model making and elongated needle acupuncture after model making. Rat spontaneous activity was examined using the BBB rating. Inflammatory cytokines TNF-α, IL-6, IL-1β and NF-kB contents were measured by ELISA. The degree of apoptosis was determined by TUNEL. Bcl-2- and Bax-positive cell levels were measured by an immunohistochemical method. Spinal p-Akt and p-ERK expressions were determined by Western blot. In vivo expressions of Cyt-C and Caspase-3 were determined by RT-PCR. The in vivo inhibitory effect on downstream Akt and ERK signaling pathways was investigated using specific inhibitors LY294002 and PD98059. Result Inflammatory reactions and neuronal apoptosis due to the inhibition of PI3K/Akt and MAPK/ERK signal pathways were involved in the damage in a rat model of spinal cord injury. The neuroprotective effect of elongated needle acupuncture was related to a decrease in the number of Bax protein-positive neurons and an increase in the number of Bcl-2 protein-positive neurons. Elongated needle acupuncture treatment improved rat motor function, activated p-Akt and p-ERK in PI3K/Akt and MAPK/ERK signal pathways by down-regulating Bax protein expression and up-regulating Bcl-2 protein expression and inhibited the expression of Cyt-C, a key factor in the mitochondrial apoptosis pathway. TUNEL detected and inhibited the activation of caspase-3 cascade in neuronal apoptosis. Application of specific inhibitors LY294002 and PD98059 to PI3K/Akt and MAPK/ERK signaling pathways inhibited the expression p-Akt and p-ERK expressions. Conclusion JIN’s three needle therapy plus mirror therapy for knee joint movement is an effective approach in treating hemiplegia after cerebral infarction, and it can improve the lower-limb motor function and the ADL.
Big needle therapy; Elongated needles; Spinal cord injury; inflammatory reaction; P-Akt; P-ERK; Bcl-2; Bax; Cyt-C; Caspase-3; Apoptosis; Rats
1005-0957(2017)03-0343-11
R2-03
A
10.13460/j.issn.1005-0957.2017.03.0343
浙江省中医药科研计划项目(2008CB067)
李长明(1988—),男,住院医师,硕士,Email:lcmmail@126.com
全仁夫(1969—),男,教授,Email:quanrenfu@126.com
2016-08-21
