春、秋季东海大桥海上风电场水域鱼类群落结构
2017-10-10侯俊利
宋 超, 侯俊利, 赵 峰, 张 涛, 杨 刚, 庄 平
春、秋季东海大桥海上风电场水域鱼类群落结构
宋 超, 侯俊利, 赵 峰, 张 涛, 杨 刚, 庄 平
(中国水产科学研究院东海水产研究所, 农业部东海与远洋渔业资源开发利用重点实验室, 上海200090)
为探明工程建设后东海大桥风电场水域鱼类群落结构特征, 作者利用等级聚类和多元统计等方法对该水域鱼类种类组成、优势种及群落结构进行分析。春、秋季共发现18种鱼类, 隶属6目9科15属; 鲈形目鱼类所占比例最高(50%), 其中又以虾虎鱼科鱼类最高(44.4%)。春季优势种为短吻红舌鳎()、棘头梅童鱼()、矛尾虾虎鱼()和孔虾虎鱼 (), 秋季优势种为棘头梅童鱼、龙头鱼()和凤鲚(); 棘头梅童鱼为春、秋季共同优势种。等级聚类和非度量多维标序排序表明, 东海大桥风电场水域鱼类群落可分为春季和秋季2个组群, 相似性检验(ANOSIM)显示2个组群差异极显著(=0.851,< 0.01)。生物与环境相关分析(BIOENV)表明温度和盐度是影响东海大桥风电场水域鱼类群落结构的最主要环境因子, 两者结合因子与鱼类群落的相关系数为0.791。本研究发现, 东海大桥风电场水域是棘头梅童鱼、凤鲚、龙头鱼等多种鱼类的育幼和索饵场所, 群落结构具有典型的季节特征。
鱼类; 种类组成; 群落结构; 东海大桥; 风电场
风电是一种清洁可再生能源, 海上拥有丰富的风能资源和广阔平坦的区域, 使得近海风力发电成为近年来研究和应用的热点[1-2]。中国风力资源储量丰富, 根据中国气象科学研究院统计评估, 中国陆地可开发的风能资源储量约为250 GW, 近海约为750 GW[3]。继2006年可再生能源法的实施, 中国风电市场持续增长, 风力发电在中国已经成为继水电之后最重要的可再生能源[4]。
海上风电场是指设立在离岸一定距离的、桩基设于海底的风力发电场。东海大桥海上风电场示范工程为国内首个海上风力发电项目, 包括一期工程位于东海大桥东侧1 km以外海域, 最北端距离南汇嘴岸线8 km, 最南端距岸线13 km; 二期工程位于东海大桥西侧, 场址距东海大桥1~5 km范围内, 距南汇岸线6~12 km。工程海域属潮坪相和三角洲前缘地貌单元, 在潮流作用下历史上以淤积为主, 海床表层主要为淤泥, 地形起伏不大[5]。该海域是重要的鳗苗资源分布水域, 每年l~4月有大量的鳗苗分布[6], 同时也是重要经济鱼类的索饵场和栖息地[7-8]。
海上工程对周边环境的影响研究包括海上核电站对海洋生态环境影响[9]、海上风电场对鸟类和海洋生物的影响等[10-14]。东海大桥海上风电工程建设后, 研究工程水域鱼类群落结构有利于全面了解工程建设后该水域的鱼类种类组成状况, 并进一步探明风电场建设对鱼类群落结构的潜在影响。
1 材料与方法
1.1 采样区域与调查方法
分别于2015年5月(春季)和9月(秋季)在东海风电场水域设置 9个站点进行渔业资源拖网调查(图 1)。拖网渔船为单船拖网船, 功率53 kW, 网具为单船底拖网, 网具总长10 m, 网口宽6 m, 网口高2m, 网囊网目20 mm。春季和秋季分别在每个站位拖网30 min, 拖速3 kn。各站位均用YSI Pro Plus同步监测温度、盐度和水深等环境因子, 样品的采集和分析均参照《海洋调查规范》(GB/T12763.6-2007), 对所有鱼类进行种类鉴定[15], 计数及称量体质量(g)。
1.2 数据标准化
对所获鱼类鉴定后, 记录每一种类的数量和质量, 换算成为单位面积的生物量(g/100 m2)和丰度(个/100 m2)。将丰度数据进行平方根转换后, 用Primer 5.0软件构建Bary-Curtis相似性矩阵, 用Bary-Curtis相似性系数聚类对群落结构进行分析[16-17]。
1.3 分析方法
采用Pinkas等[18]的相对重要性指数(Index of Relative Importance, IRI)来确定群落的重要种类组成, 其中将相对重要性指数大于20定义为优势种, 10~ 20为常见种。
IRI =(+)××100%
式中,为某种的生物量与总生物量的百分比,为该种的丰度与总丰度的百分比,为该种的出现频率[19]。
利用未加权的组平均法, 即UPGMA 进行聚类分析(CLUSTER)。利用非度量多维标序排序方法(Non-metric Multidimensional Scaling, NMDS)验证聚类分析结果的准确性[20], 用胁强系数(stress)来衡量NMDS分析结果的优劣[21]。
利用相似性分析检验(Analysis of Similarities, ANOSIM)进行不同站位和季节间差异显著性检验, 利用相似性百分比 (Similarity Percentages, SIMPER) 分析鱼类群落结构的差异和变化, 计算对样本组内相似性和组间相异性贡献较大的种类, 从而确定造成组内相似和组间差异的关键物种[21]。
利用生物与环境相关分析(Biota-Environment Analysis, BIOENV)找出与鱼类群落结构相关性最大的环境因子[22]。以上计算均由PREMIER 5.0软件完成[23]。
2 结果与分析
2.1 种类组成
春、秋季共发现鱼类18种, 隶属6目9科15 属。鲈形目(Peciformes)所占比例最高为50%(图 2), 其中以虾虎鱼科(Gobiidae, 44.4%)最多。春季共捕获9种鱼类, 隶属3目5科8属, 秋季共捕获17种鱼类, 隶属6目8科14属, 两季非共有种类数分别为1种和9种, 占比分别为11.1%和52.9%。
2.2 优势种
表1显示春、秋季共有优势种6种, 其中春季优势种为短吻红舌鳎()、棘头梅童鱼()、矛尾虾虎鱼()和孔虾虎鱼(), 其重要种类的尾数和质量所占比例之和分别达到87.93%和53.51%; 秋季优势种为棘头梅童鱼、龙头鱼()和凤鲚(), 其重要种类的尾数和质量所占比例之和分别达到71.47%和58.44%; 两个时期的共有优势种为棘头梅童鱼。
2.3 群落相似性分析
基于Bray-Curtis相似性系数的聚类分析表明, 东海大桥风电场水域鱼类群落在30%相似性水平下可分为Ⅰ、Ⅱ两组(图3), Ⅰ组为春季组, Ⅱ组为秋季组。NMDS 二维分析显示, 胁强系数 stress为 0.08 (图 4), 说明 NMDS 图可以解释鱼类群落结构, 进一步支持了聚类分析的结果。ANOSIM分析表明, 春、秋季东海大桥风电场水域鱼类群落差异极显著(= 0.851,< 0.01)。

表1 春、秋季东海风电场水域鱼类种类组成
2.4 群落典型种与分歧种
SIMPER分析指出, 春季(Ⅰ组)对组内相似性贡献较高的典型种有矛尾虾虎鱼、棘头梅童鱼、短吻红舌鳎和孔虾虎鱼, 它们对组内相似性累计贡献率达96.14%; 秋季(Ⅱ组)对组内相似性贡献较高的典型种有龙头鱼、凤鲚和棘头梅童鱼, 它们对组内相似性累计贡献率达96.43%(表 2)。组Ⅰ和组Ⅱ群落组间差异性系数为82.11%, 对组间差异性贡献较高的分歧种是矛尾虾虎鱼、棘头梅童鱼、龙头鱼、凤鲚、短吻红舌鳎和孔虾虎鱼, 它们对春、秋季相异性贡献之和为90.64%(表 2)。
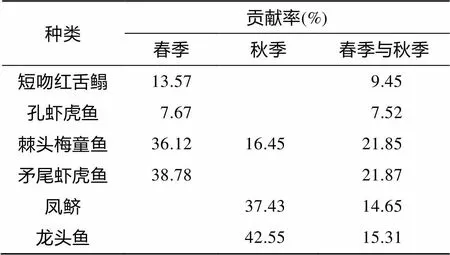
表2 组内相似性和组间相异性贡献率超过5%的种类
2.5 群落结构与环境因子的关系
BIOENV程序分析表明, 春、秋季鱼类群落结构与温度的相关性最高, 相关系数为0.778; 与盐度的相关性次之, 相关系数为0.760; 与水深的相关性最低。组合环境因子中, 群落结构与温度、盐度组合相关性最高, 相关系数为0.791。
3 讨论与结论
3.1 优势种组成
以往对东海大桥风电场及相邻水域研究表明, 东海大桥风电场春、夏季鱼类, 杭州湾海域春、秋季鱼类, 杭州湾北部水域夏、冬季鱼类以及洋山深水港水域的仔鱼组成均以鲈形目最多, 该结果与本次研究相一致[4, 24-26]。优势种方面, 本研究中春季的优势种为短吻红舌鳎、棘头梅童鱼、矛尾虾虎鱼和孔虾虎鱼, 秋季的优势种为棘头梅童鱼、龙头鱼和凤鲚。而谢旭等[24]研究得出杭州湾水域春季优势种为睛尾蝌蚪虾虎鱼、刀鲚和棘头梅童鱼, 秋季优势种为龙头鱼、棘头梅童鱼和凤鲚。上述调查与本研究中的采样季节一致, 优势种的组成相似, 秋季有共同的优势种组成, 春季的优势种有些差异, 这主要是由两者调查的区域不同所致。王淼等[25]对夏、冬季杭州湾北部鱼类调查发现夏季优势种为棘头梅童鱼, 冬季优势种为刀鲚、焦氏舌鳎(, 又名短吻红舌鳎)和髭缟虾虎鱼。上述调查与本研究的采样水域接近, 但采样季节不同, 造成两者研究中优势种的组成不同。可见, 优势种的组成与调查水域和采样季节有密切的关系, 尤其是季节对优势种的种类组成影响明显。
3.2 群落结构
基于Bray-Curtis相似性系数的ANOSIM检验表明, 春、秋季东海大桥风电场水域鱼类群落结构季节间存在显著性差异。这种结构差异主要体现在两方面。一方面是优势种组成及比例不同, 本研究中春季优势种有短吻红舌鳎、棘头梅童鱼、矛尾虾虎鱼和孔虾虎鱼, 而秋季优势种为棘头梅童鱼、龙头鱼和凤鲚, 其中棘头梅童鱼为两时期共有优势种。Cabral等[27]研究表明优势种占比不同, 群落受干扰程度也不同, 从本研究结果看, 优势种在两季节的数量及占比均不同, 说明群落结构存在一定差异, 群落受到不同程度的干扰。另一方面体现在群落典型种差异性, SIMPER分析表明, 春季典型种主要是矛尾虾虎鱼、棘头梅童鱼、短吻红舌鳎和孔虾虎鱼, 秋季典型种主要是龙头鱼、凤鲚和棘头梅童鱼, 其中组内典型种同时又是组间分歧种。不同季节间鱼类群落结构差异性较大, 但造成不同季节间差异的种类有所不同。其中, 造成春季差异的主要种类为矛尾虾虎鱼和棘头梅童鱼, 而造成秋季差异的主要种类为龙头鱼和凤鲚。在鱼类群落内, 物种分布季节动态多呈现为洄游性, 如生殖洄游、索饵洄游的更替节律, 导致鱼类群落结构的时序相对不稳定[15]。东海大桥风电场水域位于杭州湾北部近岸, 受长江径流和海洋洋流双重影响。该水域海洋生物资源丰富, 是多种鱼类的繁殖、育肥及生长的场所[28]。其中, 该水域为凤鲚重要的索饵场, 春季在长江口区盛产的凤鲚仔稚鱼, 随着长江径流漂移, 秋季时凤鲚的稚鱼和幼鱼集中到该水域进行索饵, 优势度较高。另外, 调查水域也是棘头梅童鱼的重要育幼场, 其在春季的4~6月份进行繁殖, 在春季调查中捕获的多为当年繁殖的稚鱼, 而秋季捕获的多为幼鱼, 表明该水域为棘头梅童鱼的主要的育幼和索饵场。综合ANOSIM 和 SIMPER 分析表明, 鱼类群落结构不同季节间存在显著差异, 造成这种差异的主要原因可能与棘头梅童鱼、龙头鱼和凤鲚等鱼类的繁殖和洄游有关。
3.3 群落结构与环境因子的关系
温度和盐度是造成鱼类群落结构季节变化的重要因素[29]。本次调查春季平均温度为19.0±0.5℃, 秋季平均温度为25.0℃±0.5℃, 春季平均盐度为16.4± 1.2, 秋季平均盐度为12.4±1.3, 两季节平均温、盐度分别具有显著性差异(< 0.01)。 BIOENV分析表明, 春、秋季东海大桥风电场水域鱼类群落结构与温度相关性最高, 其次是盐度, 组合环境因子中, 温度和盐度的组合对群落结构影响最大, 这说明温度和盐度是影响东海大桥风电场水域鱼类群落结构最主要的环境因子。
温度在时间尺度和盐度在空间尺度是影响海洋生物群落结构的主要环境因子, 如温度和盐度对长江口沿岸、潮间带及胶州湾和杭州湾等水域的鱼类群落结构均产生了明显影响[16, 25, 30-31]。另外, 对于洄游性鱼类其季节性洄游也是引起群落结构差异的因素之一[32]。由此可见, 群落结构的改变不仅与种类生态习性相关, 而且受多种环境因子综合作用的影响, 鱼类群落结构的形成与改变是鱼类在长期自然选择中表现出对环境的适应性结果, 东海大桥风电场工程建设是一种人为的环境干扰, 工程建设后该水域鱼类群落结构状况及其变化还需进行深入研究, 以期为该水域鱼类资源的保护和生态修复提供科学依据。
[1] 王晓蓉, 王伟胜, 戴慧珠. 我国风力发电现状和展望[J]. 中国电力, 2004, 37(1): 81-83. Wang Xiaorong, Wang Weisheng, Dai Huizhu. Present status and prospect of wind power in China[J]. Electric Power, 2004, 37 (1) : 81-83.
[2] 李伟立. 东海大桥海上风电项目二期工程场地工程地质研究[J]. 吉林水利, 2015, 7: 9-13. Li Weili. The geological research on Donghai bridge offshore wind power project site on phase II [J]. Jilin Water Resources, 2015, 7: 9-13.
[3] 肖运启, 贾淑娟. 我国海上风电发展现状与技术分析[J]. 华东电力, 2010, 38(2): 277-280.Xiao Yunqi, Jia Shujuan. Development status and technologyanalysis of offshore wind power in China[J]. East China Electric Power, 2010, 38(2): 277-280.
[4] 洪波, 王淼, 安传光, 等. 春、夏季东海大桥海上风电场鱼类组成及多样性[J]. 浙江海洋学院学报(自然科学版), 2014, 33(3): 234-239. Hong Bo, Wang Miao, An Chuanguang, et al. Component and diversity of fishes in the offshore wind farm of the East China Sea bridge in spring and summer[J]. Journal of Zhejiang Ocean University (Natural Science), 2014, 33(3): 234-239.
[5] 方宁. 东海大桥海上风电场对海域沉积物的环境影响[J]. 青岛理工大学学报, 2015, 36(2): 79-82. Fang Ning. Environmental impact of Donghai bridge offshore wind farm on marine sediments[J]. Journal of Qingdao Technological University, 2015, 36(2): 79-82.
[6] 智玉龙, 侯俊利, 张涛, 等. 长江口日本鳗鲡鳗苗时空分布特征[J]. 生态学杂志, 2013, 32(10): 2750-2755. Zhi Yulong, Hou Junli, Zhang Tao, et al. Spatiotemporal distribution patterns of elver () in the Yangtze River Estuary of China[J]. Chinese Journal of Ecology, 2013, 32(10) : 2750-2755.
[7] 周永东, 金海卫, 张洪亮, 等. 浙江北部沿岸春夏季凤鲚鱼卵及仔、稚鱼分布特征[J]. 浙江海洋学院学报(自然科学版), 2011, 30(4): 307-312. Zhou Yongdong, Jin Haiwei, Zhang Hongliang, et al. Distributions of ichthyoplankton ofin the coast of North Zhejiang Province in spring and summer[J]. Journal of Zhejiang Ocean University (Natural Science), 2011, 30(4): 307-312.
[8] 王淼, 洪波, 安传光, 等. 丰、枯水期杭州湾东北部渔业资源多样性及数量分布[J]. 上海海洋大学学报, 2016, 25(2): 246-254. Wang Miao, Hong Bo, An Chuanguang, et al. Diversity and density distribution of fishery resources in Northeast of Hangzhou Bay during flood and drought seasons[J]. Journal of Shanghai Ocean University, 2016, 25(2): 246-254.
[9] 兰志刚, 李新仲, 肖钢, 等. 海上浮式核电站温排水对海洋生态环境的影响[J]. 海洋科学, 2016, 40(6): 84-88. Lan Zhigang, Li Xinzhong, Xiao Gang, et al. Potential impacts of thermal discharge on marine environment from offshore floating nuclear power plant[J]. Marine Sciences, 2016, 40(6): 84-88.
[10] 施蓓, 丁玲, 徐凌云, 等. 东海大桥海上风电场对鸟类组成的影响[J]. 生态学杂志, 2014, 33(4): 1068-1075.Shi Bei, Ding Ling, Xu Lingyun, et al. Influence of Donghai bridge offshore wind farm on bird composition[J]. Chinese Journal of Ecology, 2014, 33(4): 1068-1075.
[11] Debusschere E, de Coensel B, Vandendriessche S, et al. Effects of offshore wind farms on the early life stages of[J]. Effects of Noise on Aquatic Life Ii, 2016, 875: 197-204.
[12] Bergstrom L, Kautsky L, Kautsky T, et a1. Effects of offshore wind farms on marine wildlife: A generalized impact assessment[J]. Environmental Research Letters, 2014, 9(3): 1-12.
[13] Stenberg C, Stottrup J G, van Deurs M, et al. Long-term effects of an offshore wind farm in the North Sea on fish communities[J]. Marine Ecology Progress Series, 2015, 528: 257-265.
[14] Goodale M W, Milman A. Cumulative adverse effects of offshore wind energy development on wildlife[J]. Journal of Environmental Planning and Management, 2016, 59: 1-21.
[15] 庄平, 王幼槐, 李圣法, 等. 长江口鱼类[M]. 上海: 上海科学技术出版社, 2006. Zhuang Ping, Wang Youhuai, Li Shengfa, et al. Fishes of the Yangtze Estuary[M]. Shanghai: Shanghai Scientific& Technical Publishers, 2006.
[16] 张涛, 庄平, 章龙珍, 等. 长江口中华鲟自然保护区底层鱼类的群落结构特征[J]. 生态学报, 2011, 31(6): 1687-1694.Zhang Tao, Zhuang Ping, Zhang Longzhen, et al. Communitystructure of demersal fish in Nature Reserve ofin Yangtze River Estuary[J]. Acta Ecologica Sinica, 2011, 31(6): 1687-1694.
[17] 张晓举, 王海棠, 周建波, 等. 辽东湾西部倾废区临近海域大型底栖动物群落结构[J].海洋科学, 2014, 38(11): 41-16. Zhang Xiaoju, Wang Haitang, Zhou Jianbo, et al. Macrobenthos community in the western Liaodong Bay ocean with dumping ground[J]. Marine Sciences, 2014, 38(11): 41-16.
[18] Pinkas L, Olinphamt M S, Iverson I L K. Food habits of albacore, Bluefin tuna, and bonito in Californian waters[J]. State of California, the Resources Agency Department of Fish and Game, Fish Bulletin, 1971, 152: 1-105.
[19] 王智, 曲方圆, 隋吉星, 等. 夏季辽东湾西部海域大型底栖动物群落结构与多样性[J]. 海洋科学, 2016, 40(1): 40-47. Wang Zhi, Qu Fangyuan, Sui Jixing, et al. Community structure and diversity of macrobenthos in the western waters of Liaodong Bay during summer[J]. Marine Sciences, 2016, 40(1): 40-47.
[20] Clarke K R, Ainsworth M. A method of linking multivariate community structure to environmental variables[J]. Marine Ecology Progress Series, 1993, 92: 205-219.
[21] 周红, 张志南. 大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J]. 青岛海洋大学学报, 2003, 33(1): 58-64. Zhou Hong, Zhang Zhinan. Rationale of the multivariate statistical software PRIMER and its application in benthic community ecology[J]. Journal of Ocean University of Qingdao, 2003, 33(1): 58-64.
[22] 汪振华, 王凯, 赵静, 等. 贻贝养殖区底层鱼类群落结构特征分析[J]. 海洋科学, 2015, 39(6): 21-31. Wang Zhenhua, Wang Kai, Zhao Jing, et al. Study of benthic fish community structure in mussel farming habitat of Ma’an Archipelago[J]. Marine Sciences, 2015, 39(6): 21-31.
[23] Clarke K R, Warwick R M. Change in marine communities: an approach to statistical analysis and interpretation (2nd edition)[M]. Plymouth, UK: Primer-E, 2001.
[24] 谢旭, 俞存根, 周青松, 等. 杭州湾海域春、秋季鱼类种类组成和数量分布[J]. 海洋与湖沼, 2013, 44(3): 656-663. Xie Xu, Yu Cungen, Zhou Qingsong, et al. Species composition and quantity distribution of fishes in the Hangzhou Bay during spring and autumn[J]. Oceanologia et Limnologia Sinica, 2013, 44(3): 656-663.
[25] 王淼, 洪波, 张玉平, 等. 夏、冬季杭州湾北部游泳动物群落结构[J]. 广东海洋大学学报, 2015, 35(3): 56-62. Wang Miao, Hong Bo, Zhang Yuping, et al. Community structure of nektons in northern Hangzhou Bay in summer and winter[J]. Journal of Guangdong Ocean University, 2015, 35(3): 56-62.
[26] 蒋玫, 王云龙, 林钦, 等. 洋山深水港水域鱼卵仔鱼分布特征[J]. 海洋环境科学, 2008, 27(1): 43-46. Jiang Mei, Wang Yunlong, Lin Qin, et al. Distribution characteristics of fish eggs and larvae around Yangshan port in Hangzhou Bay[J]. Marine Environmental Science, 2008, 27(1): 43-46.
[27] Cabral H N, Costa M J, Salgado J P. Does the Tagus Estuary fish community reflect environmental changes? [J]. Climate Research, 2001, 18(5): 119-126.
[28] 俞存根, 陈全震, 陈小庆, 等. 舟山渔场及邻近海域鱼类种类组成和数量分布[J]. 海洋与湖沼, 2010, 41(3): 410-417. Yu Cungen, Chen Quanzhen, Chen Xiaoqing, et al. Species composition and quantity distribution of fish in the Zhoushan fishing ground and its adjacent waters[J]. Oceanologia et Limnologia Sinica, 2010, 41(3): 410- 417.
[29] 蒋日进, 钟俊生, 李黎, 等. 长江口沿岸碎波带仔稚鱼类的群落结构特征[J]. 上海海洋大学学报, 2009, 18(1): 42-46. Jiang Rijin, Zhong Junsheng, Li Li, et al. The community structure of fish larvae and juveniles in the surf zone of the Yangtze Estuary[J]. Journal of Shanghai Ocean University, 2009, 18(1): 42-46.
[30] 杨刚, 洪巧巧, 张涛, 等. 长江口中华鲟自然保护区潮间带鱼类群落结构[J]. 生态学杂志, 2012, 31(5): 1194-1201. Yang Gang, Hong Qiaoqiao, Zhang Tao, et al. Fish community structure in intertidal area ofnatural reserve in Yangtze River Estuary[J]. Chinese Journal of Ecology, 2012, 31(5): 1194-1201.
[31] 翟璐, 韩东燕, 傅道军, 等. 胶州湾及其邻近海域鱼类群落结构及与环境因子的关系[J]. 中国水产科学, 2014, 21(4): 810-821. Zhai Lu, Han Dongyan, Fu Daojun, et al. Fish community structure and the relationship with environmental factors in Jiaozhou Bay and adjacent waters[J]. Journal of Fishery Sciences of China, 2014, 21(4): 810-821.
[32] 姜亚洲, 林楠, 袁兴伟, 等. 象山港游泳动物群落结构及多样性[J]. 生态学杂志, 2013, 32(4): 920-926. Jiang Yazhou, Lin Nan, Yuan Xingwei, et al. Community structure and species diversity of nektons in XiangshanBay of East China[J]. Chinese Journal of Ecology, 2013, 32(4): 920-926.
Fish community structure in the offshore wind farm of the East China Sea Bridge in spring and autumn
SONG Chao, HOU Jun-li, ZHAO Feng, ZHANG Tao, YANG Gang, ZHUANG Ping
(Key Laboratory of East China Sea and Oceanic Fishery Resources Exploitation and Utilization, Ministry of Agriculture, East China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Shanghai 200090, China)
To determine the composition of the fish community at the offshore wind farm located at the East China Sea Bridge, a cluster analysis and multivariate statistics are applied to analyze species composition, dominant species, and fish communities. Results identify 18 species belonging to 6 orders, 9 families, and 15 genera. Peciformes are the largest percentage order (50%) and Gobiidae the largest percentage family (44.4%) within this order.Dominant species in spring are,,, and, and dominant species in autumn are,, and;is the dominant species with respect to its representation in both season. Cluster and non-metric multidimensional scaling indicate that the fish community at this offshore wind farm can be clustered into two groups, spring and autumn assemblages, and ANOSIM analysis indicates significant differences in the community structure between these two groups (= 0.851,< 0.01). The relationship between fish community pattern and water temperature and salinity is determined as dominant using the BIOENV program, with a correlation coefficient of 0.791. Results show that fish such as,, andmigrate annually to the wind farm for spawning and feeding. There is an obvious difference in the community structure during different seasons, which is mainly related to the different breeding and feeding grounds of typical assemblage species during spring and autumn.
Fish; species composition; community structure; East China Sea Bridge; wind farm
(本文编辑: 谭雪静)
Oct. 27, 2016
[Special Fund for Agro-scientific Research in the Public Interest, No. 201203065]
S932.4
A
1000-3096(2017)06-0034-07
10.11759//hykx20161027001
2016-10-27;
2016-12-08
公益性行业(农业)科研专项(201203065)
宋超(1980-), 男, 山东济宁人, 博士, 主要从事鱼类生理生态学与生态修复研究, 电话: 13564403570, E-mail: songc@ecsf.ac.cn;庄平,通信作者, E-mail: Pzhuang@hotmail.com
