雷公藤4α-甲基氧化酶基因(TwSMO1)的cDNA全长克隆及表达分析△
2017-09-21关红雨胡添源赵瑜君苏平童宇茹张逸风张夏楠高伟黄璐琦
关红雨,胡添源,赵瑜君,苏平,童宇茹,张逸风,张夏楠,高伟*,黄璐琦*
(1.首都医科大学 中医药学院,北京 100069;2.中国中医科学院 中药资源中心 道地药材国家重点实验室培育基地,北京 100700)
·基础研究·
雷公藤4α-甲基氧化酶基因(TwSMO1)的cDNA全长克隆及表达分析△
关红雨1,胡添源1,赵瑜君2,苏平2,童宇茹2,张逸风1,张夏楠1,高伟1*,黄璐琦2*
(1.首都医科大学 中医药学院,北京 100069;2.中国中医科学院 中药资源中心 道地药材国家重点实验室培育基地,北京 100700)
目的:克隆得到雷公藤一型4α-甲基氧化酶(TwSMO1)(GenBank KX987126)全长基因,并对其进行序列分析及初步的功能验证。方法:根据雷公藤转录组数据,利用cDNA末端快速扩增(RACE)技术克隆得到雷公藤4α-甲基氧化酶基因(TwSMO1),并利用相关软件对所得核酸序列及其所编码的氨基酸序列进行生物信息学分析。用茉莉酸甲酯(MeJA)诱导雷公藤悬浮细胞,用实时荧光定量(RT-PCR)的方法分析诱导不同时间点后雷公藤悬浮细胞中的TwSMO1基因的相对表达量。结果:克隆所得的TwSMO1基因cDNA全长1505 bp,开放阅读框900 bp,编码299个氨基酸,在线预测其所编码蛋白分子量为34.55 kDa,理论等电点为6.89。多重序列比对结果显示其与其他物种的SMO1氨基酸序列有较高同源性,且包含SMO1基因家族的3个保守域,系统进化树将其归为4α-甲基氧化酶第一家族基因,遂将其命名为TwSMO1。RT-PCR结果表明MeJA诱导后TwSMO1基因在1 h后表达量达到最高,约是对照组的450倍。结论:本研究首次克隆得到雷公藤TwSMO1基因,并对其进行生物信息学分析及MeJA诱导表达分析,为深入研究此基因功能及阐述雷公藤甾醇生物合成途径奠定基础。
雷公藤;4α-甲基氧化酶;克隆;生物信息学分析;诱导表达
甾醇类化合物普遍存在于真核生物中,对于调节植物生长发育具有重要作用,同时作为组成膜结构的基本成分,具有调节细胞膜流动性和渗透性的作用,可在细胞膜间交流脂类和蛋白类化合物,并可促使细胞膜完成细胞形态的变化及细胞分化等活动[1]。在高等植物中,甾醇类化合物经甲羟戊酸途径(MVA)合成,与三萜类化合物的上游合成途径基本一致,而环阿屯醇合酶(CAS)促使代谢流向甾体类化合物方向流动,在此后生成甾醇类化合物的过程中,植物甾醇若想发挥组成膜结构的功能,需要脱去C-4位的两个甲基[2]。SMO对C-4位的脱甲基反应起到起始氧化作用[3],继而有助脱甲基过程的发生。SMO分为两个家族,分别在甾醇下游的不同步骤进行C4位的脱甲基化反应。SMO1催化4,4-二甲基-9β,19-环丙烷环甾醇生成4,4-二甲基甾醇,而SMO2则有助于促进4α-甲基甾醇脱甲基生成C-4甲基甾醇。
本研究中,根据雷公藤TripterygiumwilfordiiHook.f.转录组数据,克隆得到一条SMO基因,并通过生物信息学分析对该基因及其所编码的蛋白进行初步预测,同时分析了经茉莉酸甲酯诱导后悬浮细胞中基因表达含量的变化,为进一步研究该基因功能奠定基础。
1 材料
1.1 悬浮细胞
雷公藤悬浮细胞暗培养于0.5 mg·L-12,4-D+0.1 mg·L-1KT+0.5mg·L-1IBA的MS液体培养基中,培养条件为25 ℃、120 r·min-1。
1.2 菌株
本实验所用的E.coilTrans1-T1菌株购自北京全式金生物技术有限公司。
1.3 试剂及仪器
本研究所用的RNA纯化试剂盒及琼脂糖凝胶DNA回收试剂盒[天根生化科技(北京)有限公司];SMARTerTMRACE cDNA Amplification Kit(美国Clontech公司);克隆载体pEASY-T3 Cloning Kit及菌液PCR所用2×EasyTaq PCR SuperMix(北京全式金生物技术有限公司);KAPA SYBR©Tetra System(美国KAPA Biosystems公司);ABI verity PCR扩增仪;LightCycler©480II荧光定量PCR仪。
2 方法与结果
2.1 雷公藤悬浮细胞RNA的提取
用茉莉酸甲酯(MeJA)诱导雷公藤悬浮细胞,使MeJA终浓度为50 μmol·L-1,分别取诱导1、4、12、24、72、72 h后的悬浮细胞用CTAB法提取总RNA,并经DNase I去除基因组污染及RNA纯化试剂盒纯化,用1%的琼脂糖凝胶检测所提取RNA质量。
2.2 TwSMO1末端快速扩增
将上述纯化RNA用SMARTerTMRACE cDNA Amplification Kit反转录成cDNA。把转录组中所得基因片段在NCBI进行BlastX比对,结果显示该片段5′端完整,3′端有缺失,设计3′ RACE Primer以得到cDNA全长序列,引物序列为TwSMO1 (3′ RACE):CGTTTGCGGAGACTTTGTGGTTC,以3′ RACE Ready cDNA为模板进行末端扩增,反应体系为:5 μL 10×Adantage 2 PCR Buffer、1 μL dNTP mix (10 mmol·L-1)、1 μL 50×Advantage 2 polymerase及34.5 μL PCR-grade water,将上述体系稍稍混匀后再加入5 μL 10×UPM、1 μL 3′RACE Primer (10 μmol·L-1)和2.5 μL cDNA模板。反应程序为:94 ℃预变性 5 min;94 ℃ 30 s,68 ℃ 30 s,72 ℃ 3 min,共进行25个循环;72 ℃总延伸7 min。PCR产物用琼脂糖凝胶DNA回收试剂盒纯化后,连接pEASY-T3克隆载体,而后转入大肠杆菌Trans1-T1感受态细胞,涂布于LB+Amp (100 mg·L-1)的平板上,37 ℃培养过夜。挑取单克隆菌斑参照全式金EasyTaq Mix说明书进行菌液PCR验证,选择阳性菌斑进行测序验证,测序结果与先前转录组数据拼接得到TwSMO1全长cDNA全长序列。
2.3 TwSMO1全长ORF的克隆
利用在线ORF Finder工具从TwSMO1全长cDNA序列中找到TwSMO1 的开放阅读框即ORF区,并在其ORF两端设计特异性引物扩增其ORF全长序列,引物序列为:TwSMO1-F,ATGCTGCCGTACGCCACCATCG;TwSMO1-R,CTAATCAGATTTAACATTGTACAATCCTCC;用NEB Phusion HF高保真酶进行扩增。反应体系:正反引物各2.5 μL,2×Phusion HF PCR Master Mix 25 μL,cDNA模板1 μL,用水补足至50 μL。反应条件为:98 ℃ 30 s;98 ℃ 10 s,55 ℃ 30 s,72 ℃ 30 s,共35个循环;72 ℃终延伸7 min。反应产物经切胶回收后,连接pEASY-T3载体,转化E.coilTrans1-T1感受态细胞并于LB+Amp (100 mg·L-1)的固体培养基上培养过夜,挑取单克隆菌斑进行菌液PCR,挑取阳性菌斑测序,从而得到TwSMO1 基因完整ORF 。
2.4 TwSMO1基因生物信息学分析
将获得的TwSMO1全长序列用ORF Finder网站查找开放阅读框,并在NCBI网站与其他物种核酸及氨基酸序列进行同源比对,同时下载同源序列用DNAMAN软件进行多重序列比对,MEGA 5.2建立系统进化树;用Compute PI/Mw (http://web.expasy.org/compute_pi/) 网站预测其氨基酸序列所编码蛋白质的分子量和理论等电点;TMHMM(http://www.cbs.dtu.dk/services/TMHMM-2.0/)分析蛋白跨膜域结构;分别用Predictprotein (http://www.predictprotein.org/)和SWISS-MODEL(http://swissmodel.expasy.org/)网站进行蛋白二级结构分析及三维同源建模。
2.5 TwSMO1基因的诱导表达分析
茉莉酸甲酯(MeJA)作为一种非生物诱导子,具有促进次生代谢物累积的作用。将雷公藤悬浮细胞中提取得到的RNA反转为cDNA后,用实时荧光定量PCR(RT-PCR)方法检测TwSMO1在不同诱导时间表达含量的变化。根据RT-PCR引物设计原则,设计特异性引物(qTwSMO1-F:GTGGATAGTTTTGCGGC;qTwSMO1-R:TAGCTTTTTCTGATATCGAAAG)以扩增TwSMO1基因片段;所用内参基因为延伸因子基因(eukaryoticelongationfactor-1α)Efα1U和Efα1L;引物序列为:Efα1U,TAGCTTTTTCTGATATCGAAAG;Efα1L,CACTGGTGGTTTTGAGGCTGGTATCT[4]。20 μL反应体系包括:10 μL 2×KAPA SYBR® FAST qPCR Master Mix 2 Universal,各0.5 μL的正反向引物,1 μL cDNA及8 μL ddH2O。PCR反应条件为:95 ℃ 5 min;而后95 ℃ 3 s,60 ℃ 30 s,共重复40个循环。每个反应重复3次,用2-△△Ct的方法分析TwSMO1基因相对于内参基因的相对表达量。
3 结果与分析
3.1 TwSMO1基因全长ORF的克隆
根据转录组所得的SMO1基因片段信息设计3′ RACE引物并进行cDNA末端扩增,随后将RACE测序结果与转录组数据拼接,得到TwSMO1全长序列,在ORF两端设计特异性引物扩增ORF全长,结果如图1所示,在900 bp处得到单一明亮的条带。
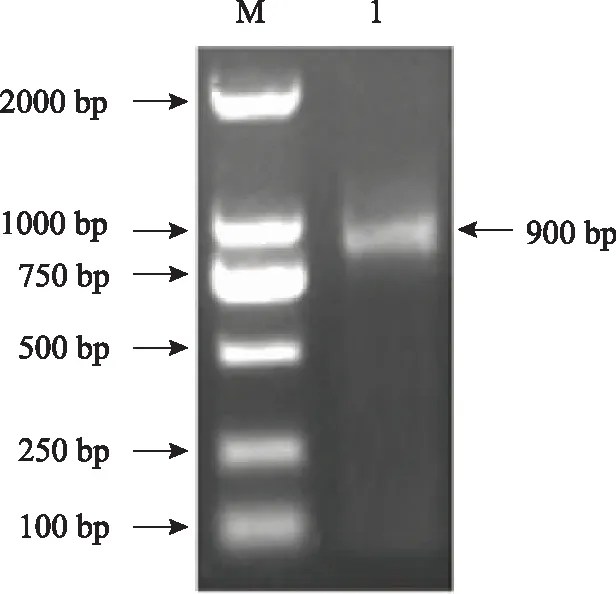
注:M.2000bp marker;1.TwSMO1基因ORF扩增条带。图1 TwSMO1基因全长ORF克隆
3.2 TwSMO1基因生物信息学分析
TwSMO1基因全长1505 bp,开放阅读框 900 bp,编码299个氨基酸。多重序列比对结果显示TwSMO1含有SMO1基因家族的3个保守域:一个HX3H及两个HX2HH。同时其氨基酸序列与毛果杨Populustrichocarpa及可可树Theoboromacacao的同源性均达81%,与川桑Morusnotabilis及蒺藜苜蓿的Medicagotruncatula的相似性分别为79%和78%,与其他物种比对结果如图2所示。
ExPASy在线预测其氨基酸所编码蛋白的分子量为34.55 kDa,理论等电点为6.89。TMHMM蛋白跨膜域结构预测结果显示(见图3):TwSMO1为膜蛋白,含有3个跨膜区域,分别位于37~59、92~114及184~206位氨基酸,此外,1~36和115~183位氨基酸位于膜外,60~91、207~299位氨基酸位于膜内。

注:TcSMO1.Theobroma cacao,XP_007039995.1;DhSMO1.Dorcoceras hygrometricum,KZV19433.1;PtSMO1.Populus trichocarpa,XP_006368716.1;MnSMO1.Morus notabilis,XP_010108130.1;MtSMO1.Medicago truncatula,XP_003608797.1;GaSMO1.Gossypium arboreum,KHG07507.1;AtSMO1.Arabidopsis thaliana,NP_567670.1;Region 1~3为SMO1基因家族的3个保守域:HX3H、HX2HH及HX2HH。图2 TwSMO1与其他物种的SMO1氨基酸序列比对

图3 TwSMO1蛋白跨膜域分析
Predictprotein预测结果表明其蛋白质二级结构以螺旋结构为主,占到65.89%,此外成环结构和线性结构分别占到31.77%和2.34%。SWISS-MODEL预测蛋白质3D结构结果如图4所示,以4zr 0.1.A为模板建模,序列同源性为22.14%,建模范围为126~268位氨基酸。
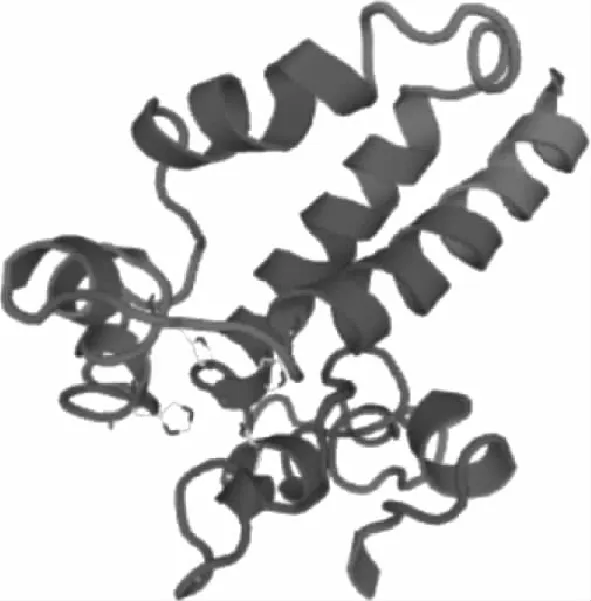
图4 模拟TwSMO1蛋白质的3D结构
将不同物种的SMO1和SMO2家族基因氨基酸序列进行进化树归类,结果如图5所示,进化树按基因家族分为两大分支,SMO1为一分支,SMO2聚为另一分支,而克隆所得TwSMO基因的氨基酸序列与其他物种的SMO1序列归为一类,故其为SMO1家族基因,同时可看出雷公藤SMO1与毛果杨(Populustrichocarpa)中的SMO1亲缘关系最近。
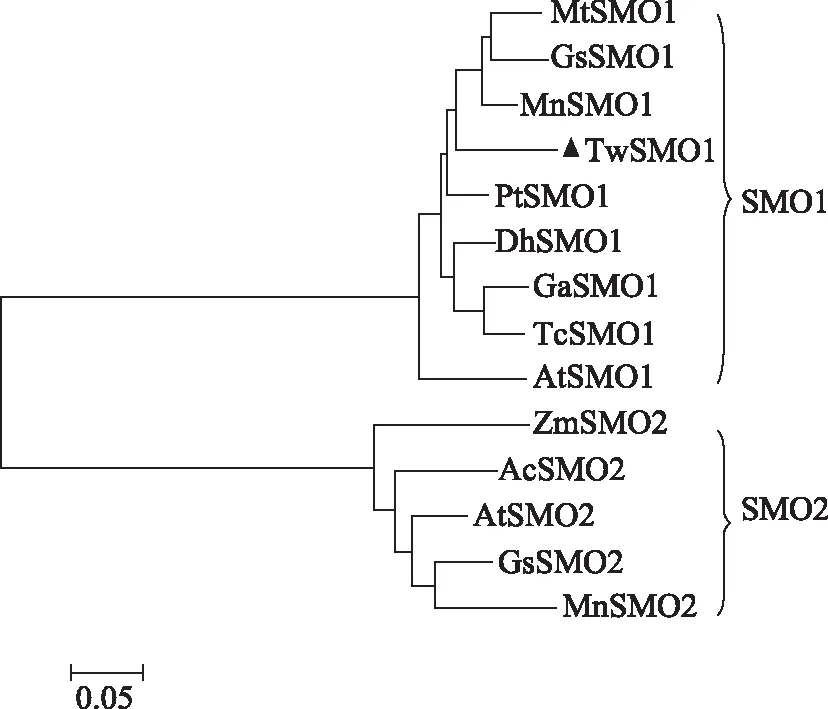
注:MtSMO1.Medicago truncatula,XP_003608797.1;GsSMO1.Glycine soja,KHN04765.1;MnSMO1.Morus notabilis,XP_010108130.1;PtSMO1.Populus trichocarpa,XP_006368716.1;DhSMO1.Dorcoceras hygrometricum,KZV19433.1;GaSMO1.Gossypium arboreum,KHG07507.1;TcSMO1.Theobroma cacao,XP_007039995.1;AtSMO1.Arabidopsis thaliana,NP_567670.1;ZmSMO2.Zostera marina,KMZ74989.1;AcSMO2.Ananas comosus,OAY85395.1;AtSMO2.Arabidopsis thaliana,Q9ZW22.2;GsSMO2.Glycine soja,KHN45289.1;MnSMO2.Morus notabilis,XP_010106554.1。图5 TwSMO1与其他物种的SMO氨基酸序列的进化树
3.3 TwSMO1基因的诱导表达分析
用实时荧光定量的方法检测经MeJA诱导不同时间的悬浮细胞中TwSMO1的表达量,并通过2-△△Ct法计算TwSMO1基因的相对表达量,结果如图6所示,经MeJA诱导1 h后,TwSMO1基因的表达量相对于对照组有显著提高,是对照组的450多倍,随后即呈下降趋势,在诱导后约12 h诱导组表达量与对照组趋于一致。
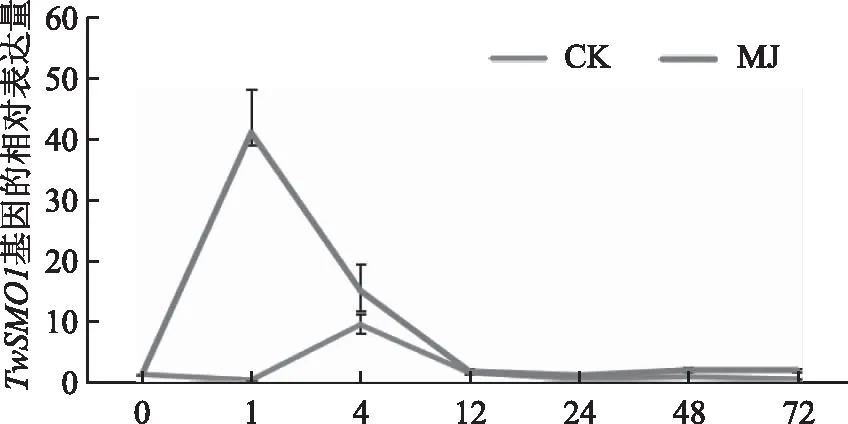
图6 MeJA诱导雷公藤悬浮细胞TwSMO1基因表达量分析
4 讨论
雷公藤作为我国传统中药,应用历史悠久,广泛应用于类风湿性关节炎、系统性红斑狼疮及麻风等疾病的治疗[5-6]。在前期研究中,许多参与植物次生代谢的重要功能基因已被克隆并进行了功能验证[7-12],但参与雷公藤甾醇类化合物生物合成的中下游基因研究才刚刚开始。
甾醇类化合物主要起调节植物生长发育及组成膜结构的作用,高等植物中甾醇种类丰富,主要为谷甾醇、豆甾醇及24-甲基胆甾醇等化合物[10]。在SMO的作用下,甾醇生物合成中C4位脱掉两个甲基从而使甾醇具有组成细胞膜结构的作用。同时有研究表明,将拟南芥中克隆得到的SMO1、SMO2基因分别在烟草属植物中沉默,分别导致了4,4-二甲基-9β,19-环丙烷环甾醇和4α-甲基甾醇的累积,从而可影响4,4-二甲基甾醇和C-4甲基甾醇的表达水平。更有研究表明,在烟草中过表达黄花蒿AaSMO1基因可提高总甾醇含量,从而使其后代能够更好地发芽和生长;此外,该基因还具有提高植物对干旱环境的耐受力作用[2]。
本研究克隆得到一条全长TwSMO1基因,其cDNA全长为1505 bp,开放阅读框900 bp,编码299个氨基酸,多重序列比对及进化树分析将此基因归为SMO1基因家族,同时结果显示其与其他物种的SMO1氨基酸序列具有较高同源性,且实验表明,MeJA诱导雷公藤悬浮细胞1 h后,TwSMO1基因表达量急剧上升并达到峰值。此研究将为深入研究雷公藤SMO1基因功能及阐释雷公藤甾醇生物合成途径奠定基础,同时可对调节雷公藤植株的生长及抗旱能力起到一定的意义。此外,雷公藤红素作为雷公藤中具有重要药理活性的三萜类化合物,其药理活性广泛[13-14],其下游生物合成位于甾醇类化合物的旁路,利用RNA干扰等手段抑制甾醇通路上基因的表达可能有利于三萜类化合物的累积,对于促进雷公藤红素的生物合成具有一定意义。
[1] Singh A,Jindal S,Longchar B,et al.Overexpression ofArtemisiaannuasterol C-4 methyl oxidase gene,AaSMO1,enhances total sterols and improves tolerance to dehydration stress in tobacco[J].Plant Cell Tissue & Organ Culture,2015,121(1):167-181.
[2] Darnet S,Ranier A.Plant sterol biosynthesis:identificationof two distinctfamilies of sterol 4alpha-methyl oxidases[J].BiochemJ,2004 ,378(3):889-898.
[3] Rondet S,Taton M,Rahier A.Identification,characterization,and partia l purifica tionof 4alpha-carboxysterol-C3-dehydrogenase/C4-deca rboxylase fromZeamays[J].Arch BiochemBiophys,1999,366(2):249-260.
[4] 祝传书,陈欣,郭嘉,等.雷公藤1-脱氧-d-木酮糖-5-磷酸还原酶基因(twdxr)的克隆与表达分析[J].农业生物技术学报,2014,22(3):298-308.
[5] 黄玲.雷公藤药理作用研究进展[J].江西中医药,2000(2):45-46.
[6] 水光兴,万毅刚,蒋春明,等.雷公藤及其活性成分药效学和药理学研究的若干进展[J].中国中药杂志,2010,35(4):515.
[7] Tong Y R,Su P,Zhao Y J,et al.Molecular cloning and characterization ofDXSandDXRgenes in terpenoid biosynthetic pathway fromTripterygiumwilfordii.[J].Int J Mol Sci,2015,16:25516-25535.
[8] Zhang M,Su P,Zhou Y J,et al.Identification of geranylgeranyl diphosphate synthase genes fromTripterygiumwilfordii[J].Plant Cell Rep,2015,34:2179-2188.
[9] 张萌,苏平,刘雨佳,等.雷公藤牻牛儿基牻牛儿基焦磷酸合酶基因全长cDNA的获得及生物信息学分析[J].中国中药杂志,2015,40(6):1066-1070.
[10]童宇茹,苏平,张萌,等.雷公藤MCT基因的全长克隆与表达分析[J].中国中药杂志,2015,40(22):4378-4383.
[11]Zhao Y J,Chen X,Zhang M,et al.Molecular cloning and characterisation of farnesyl pyrophosphate synthase fromTripterygiumwilfordii[J].PLoS One,2015,10:1-13.
[12]赵瑜君,张萌,刘雨佳,等.雷公藤乙酰CoA酰基转移酶基因全长cDNA克隆及表达分析[J].中国中药杂志,2015,40(5):847-852.
[13]陈铭祥,冯玉静,王定勇,等.雷公藤红素的研究进展[J].中成药,2010,32(3):473-476.
[14]Zhang L X,Yu F K,Zheng Q Y,et al.Immunosuppressive and-antiinflammatory activities of tripterine[J].Acta Pharm Sin,1990,25:573-577.
CloningandExpressionAnalysisof4α-methylOxidase1cDNAinTripterygiumwilfordii
GUANHongyu1,HUTianyuan1,ZHAOYujun2,SUPing2,TONGYuru2,ZHANGYifeng1,ZHANGXianan1,GAOWei1*,HUANGLuqi2*
(1.SchoolofTraditionalChineseMedicine,CapitalMedicalUniversity,Beijing100069,China; 2.StateKeyLaboratoryofDao-diHerbs,NationalResourceCenterforChineseMateriaMedica,ChinaAcademyofChineseMedicalSciences,Beijing100700,China)
Objective:To obtain the full-length cDNA of 4α-methyl oxidase 1(TwSMO1)fromTripterygiumwilfordii,and further demonstrate the sequence information and function.Methods:Based on the transcriptome data ofT.wilfordii,the rapid-amplification of cDNA ends (RACE) technology was used to clone 4α-methyl oxidase gene (SMO1).The bioinformatics software was used to analyze the information of nucleic acid sequence and the coding amino acids sequence.T.wilfordiisuspension cells were induced by methyl jasmonic acid (MeJA),and the genetic differential expression ofTwSMO1 were detected through RT-PCR method.Results:The full-length cDNA sequence ofTwSMO1 gene was 1505 bp,containing a 900 bp open reading frame and encodes 299 amino acids.The prediction of molecular weight of coding protein was 34.55 kDa and the theory of isoelectric point was 6.89.Multiple sequence alignment results showed that the amino acid sequence of TwSMO1 had a high homology with other species,including three conservative domains of SMO1 gene family.Phylogenetic tree analysis indicated that the gene was classified as first family of 4α-methyl oxidase gene,hence we named itTwSMO1.RT-PCR results showed that,after induced by MeJA,the level ofTwSMO1 gene expression was about 450 times higher than the control group at 1 h.Conclusion:TheTwSMO1 gene was cloned fromT.wilfordiifirstly,the bioinformation of the sequence was analyzed and the enzyme function was verified.The results laid the foundation for further research on its gene function and sterol biosynthetic pathway ofT.wilfordii.
Tripterygiumwilfordii;4α-methyl oxidase;cloning;Bioinformatics analysis;inducible expression
国家自然科学优秀青年科学基金项目(81422053);国家杰出青年科学基金项目(81325023)
] 高伟,教授,博士生导师,研究方向:分子生药学与中药资源学,Tel:(010)83911633,E-mail:weigao@ccmu.edu.cn;黄璐琦,院士,研究方向:分子生药学与中药资源学,Tel:(010)64014411,E-mail:huangluqi01@126.com
10.13313/j.issn.1673-4890.2017.6.004
2016-09-30)
*[
