荻与五节芒种间杂交F1群体主要农艺性状的遗传及相关性分析
2017-09-13蒋建雄陈智勇覃静萍易自力
艾 辛, 蒋建雄, 肖 亮, 陈智勇, 覃静萍, 易自力*
(1. 湖南农业大学园艺园林学院, 湖南 长沙410128;2. 江苏大学环境与安全工程学院 生物质能源研究所, 江苏 镇江 212013;3. 湖南农业大学生物科学技术学院, 湖南 长沙410128)
芒属(Miscanthus)植物作为最有发展前景的C4草本能源植物备受国际关注[1-2],近几年来,国内外从资源植物学、生态与农学、细胞和分子水平,遗传改良以及能源利用等方面做了大量的研究工作[3-5],为芒属植物的能源化利用奠定了基础。芒属植物的利用前提是要有在边际性土地生产的成本低廉的原料[6],但现有的芒属植物资源各有优缺点,不适合直接作为品种进行大面积栽培,需要将芒属植物4个主要种的优良性状聚合培育高产、优质、抗逆的新品种。目前栽培较多的是来源于日本的天然三倍体杂交种‘奇岗’[7-8],但由于三倍体的不育性,无法以此为基础进行杂交育种工作。芒属植物的杂交育种工作因花期不遇的困难,进展缓慢,更缺乏对芒属植物植株及产量相关性状的研究,严重制约了对芒属植物资源的利用。
植物的遗传改良就是要将分散在不同个体间控制优良性状的优良基因聚合在一起,获得具有更多优良性质的新品种。植物的性状分为质量性状和数量性状,一些极为重要的经济性状(如作物植株高度、茎秆粗细、产量、生育期、籽粒重等)都是数量性状,在植物全部性状中占有很大的比重[9]。数量性状在一个群体内的各个体间表现为连续变异的性状,较易受环境影响,同一数量性状的遗传受为数甚多的基因支配,遗传较为复杂。盖钧镒等[10-11]认为,控制数量性状的多个基因在效应上可以是不等的,效应大的表现为主基因,效应微小的表现为多基因,即这类性状的遗传为主基因+多基因混合遗传模型。利用这一分析方法,对多种作物的生育期[12]、株高和叶片数[13]、生长特性[14]和逆境适应性状[15]等的遗传本质得到了进一步明确。
由于芒属植物高度自交不亲和[16],所以无法获得高世代自交系,异质化程度很高,在F1代就会有高度分离,接近纯合亲本杂交种F2代的分离水平,可以将芒属植物的杂交F1代群体作为假F2代群体[17-18]进行遗传分析,这一方法已在草坪草[19]和菊花[20]中得到了成功的运用。芒属植物中的荻(M.sacchariflorus)具有耐寒抗冻、耐干旱盐碱和发达的地下根状茎、纤维品质好和自然衰老等优良特性[21-22],但生物产量低;五节芒(M.floridulus)具有耐瘠薄、生长快、生物产量高等优点[23],但不能自然衰老和耐旱抗冻,如果能将荻和五节芒2个种间的优良性状聚合在一起,就能选育出逆境耐受性好、生物质产量高的新品种。本试验通过荻和五节芒2个种间杂交种F1群体的植株和产量相关农艺性状的表现和杂种优势分析,借用盖钧镒[10-11]关于单个分离世代的数量性状分离分析的方法,探讨芒属植物种间杂交后代农艺性状的遗传规律,明确相关性状的遗传模型和基因作用方式,估计主基因的遗传效应,为荻和五节芒杂交亲本选配、扩大荻和五节芒遗传基础和选择合适的育种方法提供理论依据,同时为今后开展芒属植物分子育种的深入研究奠定理论基础。
1 材料与方法
1.1 试验材料
供试材料为荻(Micanthus.sacchariflorus)B0134和五节芒(M.floridulus)A0430,荻B0134采集于陕西周至县(N 34.08°,E 106.75°),五节芒A0430采集于湖南洞口县(N 27.07°,E 110.53°),经过3年的观察,性状表现稳定,现保存于湖南农业大学芒属植物种质资源圃。在长沙地区,荻B0134的花期为6月中下旬,五节芒A0430的花期在7月上中旬,花期不遇,无法正常进行杂交。2010年春季将荻B0134和五节芒A0430移栽至口径45 cm的塑料盆中,通过光照和处理进行花期调控,2010年10月选择花期相遇的荻和五节芒的花序用硫酸纸袋进行套袋杂交,从荻B0134中获得的种子(荻B0134×五节芒A0430)为正交F1杂交种子,从五节芒A0430花序上收获的种子(五节芒A0430×荻B0134)为反交F1杂交种子。次年3月初将正反交杂交种分别播种在育苗盘中,6月上旬将正反交杂交种和父母本荻B0134和五节芒A0430的无性分蘖株,按照1.80 m×1.80 m的行株距移栽至湖南浏阳石佳村湖南农业大学芒属植物实验基地。移栽后浇足定蔸水,成活后不再施肥和浇水。根据杂交种的田间性状表现和室内分子标记分析,确定真假杂种[24]。
1.2 田间性状调查
2012年和2013年对父母本无性分蘖株、148份正交F1实生苗进行持续观察,测定亲本和正交F1群体中单株的茎节数(Non:Numbers of stem nodes)、叶片长(Lol:Length of leaf)、叶宽(Wol:Width of leaf)、主茎长(Lms:Length of main stem)、花茎长(Lfs:Length of flower stalk)、花序长(Loi:Length of inflorescence)、株高(Ph:Plant height)、基部茎径(Sd:Stem diameter)、分蘖数(Not:Number of tiller)、单茎均重(Wst:Average dry weight of single tiller)、最大茎重(Wbt:Dry weight of the biggest single tiller)、单株重(Wop:Dry weight of plan)等12个农艺性状,除分蘖数和单株重外,其他植株性状重复3次,并计算2年性状调查的平均值。性状测定的标准和方法参考文献[25]。叶片的长、宽在植株生长的旺盛期测定,其他性状的测定在每年的12月下旬。
1.3 杂种优势分析及显著性检测
杂种优势分别以中亲优势、中亲优势率和超高亲优势、超高亲优势率表示[26]。中亲值(mid-parents value,MPV)为双亲平均值[MPV=(P1+P2)/2],高亲值(high-parents value,HPV)为双亲中性状值最高的值。中亲优势(Hm)=F1-MPV,中亲优势率(RHm)=(F1-MPV)/MPV×100%。高亲优势(Hh)=Fm-HPV,高亲优势率(RHh)=(Fm-HPV)/HPV×100%。
1.4 杂交种植株性状的混合遗传分析
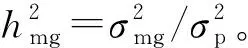
1.5 数据分析
采用DPS14.2统计软件进行基本描述性数据的统计分析及样本均值t检验、偏相关性分析方法对农艺性状进行相关性分析。
2 结果与分析
2.1 荻与五节芒杂交种主要农艺性状在亲本及杂交后代的分布
荻与五节芒的差异较大,他们的杂交种F1(荻B0134×五节芒A0430)148个群体植株和产量相关性状的差异较大,其描述性统计结果如表1所示。植物学性状包括叶片、茎杆、花序性状,各性状的表型值有较大的差异,各性状的范围为9.76%~42.87%,其中以叶片各性状的变异系数最大。各性状变异系数由大到小的顺序依次为:单株重>分蘖数>最大茎重>单茎均重>花茎长>茎节数>叶片宽>花序长>叶片长>主茎长>基部茎径>株高,总的来说,株高和基部茎径的相关性状变异较小。杂交种群体产量相关性状的变异系数普遍较高,说明产量相关性状在群体中有较大的分离,其中变异系数最大的为单株重,变异系数为42.87%,其次是分蘖数,变异系数为35.96%,说明了荻与五节芒种间杂交后代在产量性状上具有较大的选择价值,具有较高的选择效率。
将12个主要农艺性状的调查结果进行频次分布的分析,绘制次数分布图(图1)。由图可知,所有性状的分布是连续的,符合正态分布,以及不同程度的的偏态分布,符合主基因+多基因的遗传特征。
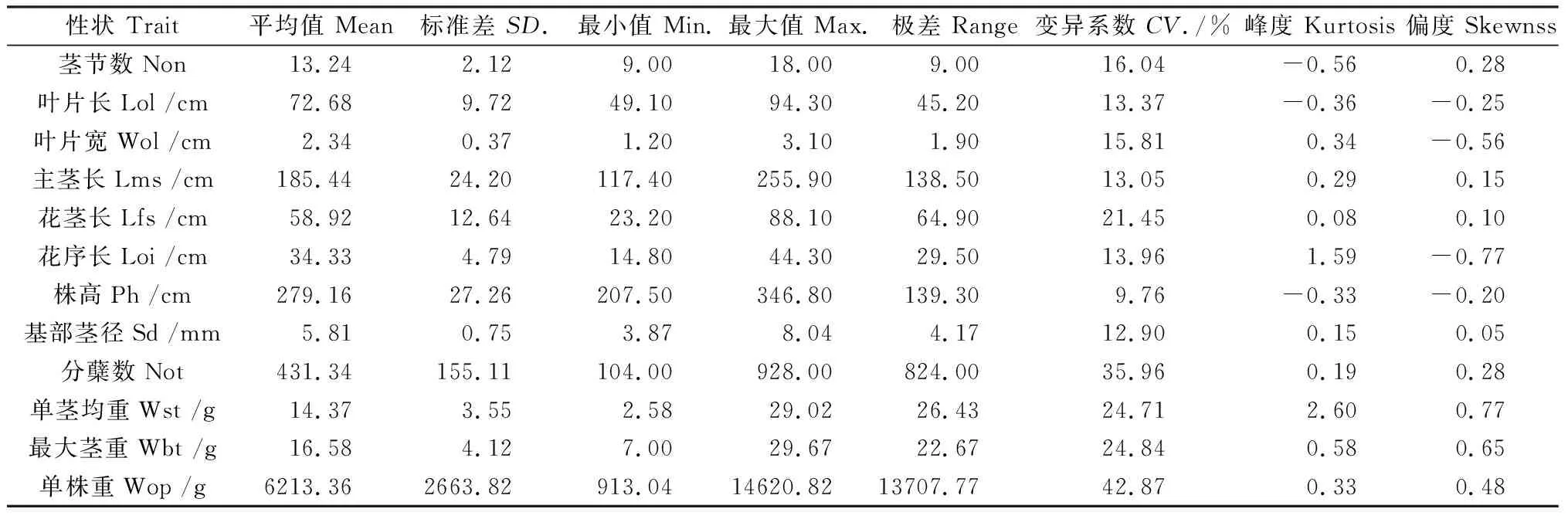
表1 荻与五节芒杂交F1群体主要数量性状表型特征值Table 1 Phenotypic statistical values of 12 agronomical traits in F1 population between M.sacchariflorus and M.floridulus
2.2 荻与五节芒杂交种主要数量性状的杂种优势表现
荻B0134与五节芒A0430杂交种的各性状杂种优势的度量值如表2所示。茎节数、主茎长、株高、分蘖数、单株重表现为超中优势,超中亲杂种优势率最高的为分蘖数(580.35%),其次为单株重(160.99%)、主茎长(64.62%)、茎节数(36.49%)和株高(30.65%),而基部茎径、单茎均重、最大茎重、叶片长、叶片宽为负中亲优势。杂交种在茎节数、主茎长、分蘖数、单株重表现为正向超高亲优势,以分蘖数的超高亲优势率最为显著,达到了304.63%,单株重也表现较高的超高亲优势率(33.40%)。杂交种F1群体中超中亲优势植株比例大于50%的性状有茎节数(100%)、主茎长(97.97%)、株高(80.41%)、分蘖数(100%)和单株重(93.24%),超高亲优势植株比例大于50%的性状有茎节数(61.49%)、主茎长(77.03%)、分蘖数(100%)和单株重(72.97%),说明荻与五节芒杂交种F1群体的重要农艺性状普遍存在杂种优势,可利用种间杂交种产量相关性状的杂种优势进行芒属植物遗传改良。
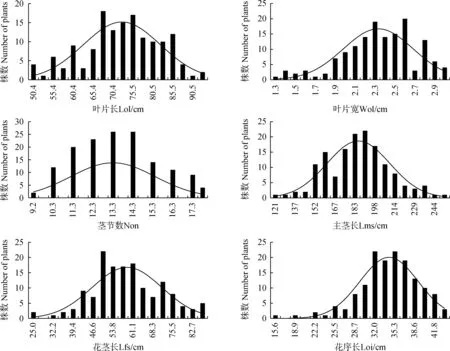
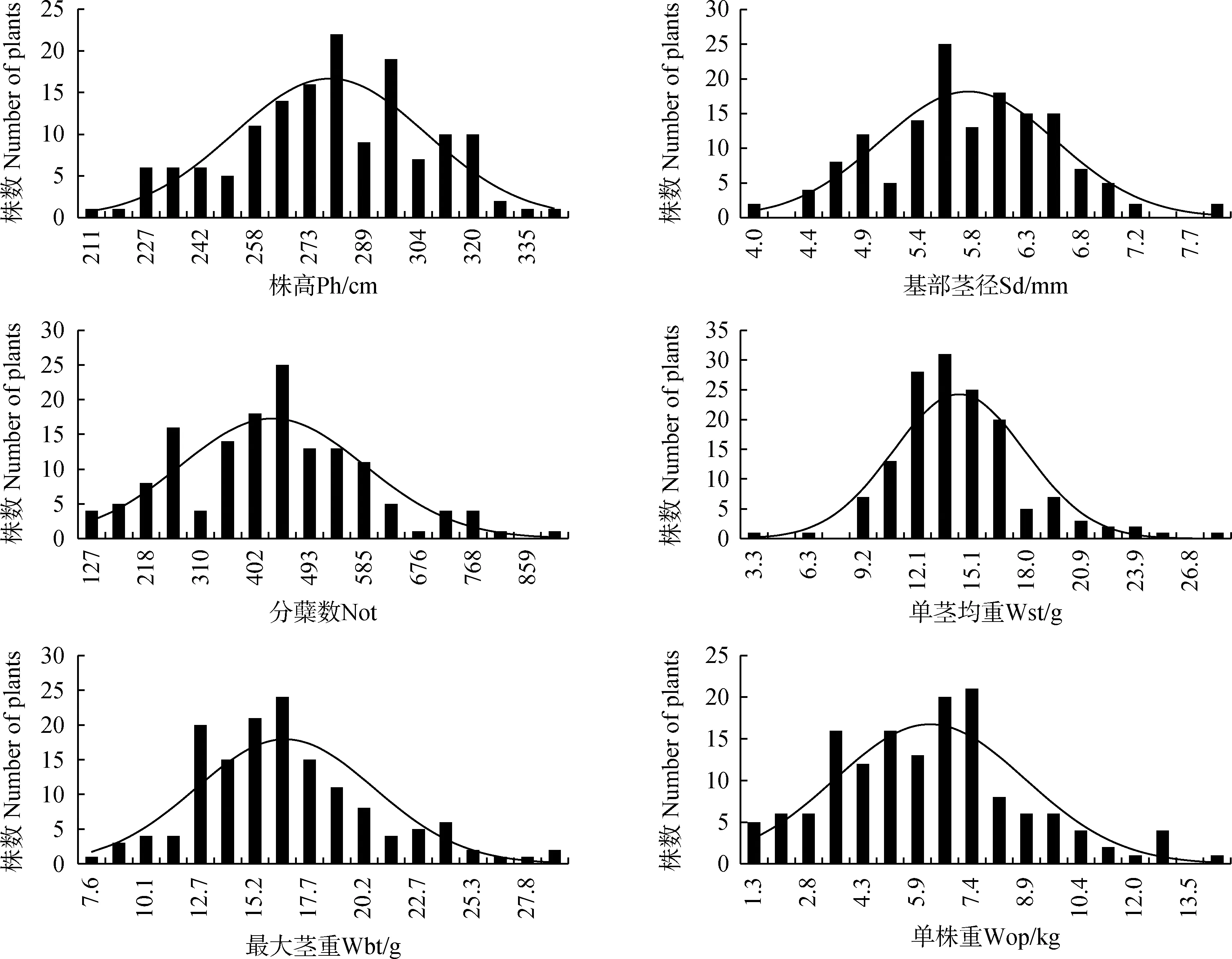
图1 荻与五节芒杂交种F1群体主要农艺性状的分布频次Fig.1 Distribution frequency of 12 agronomical traits in F1 population between M. sacchariflorus and M. floridulus
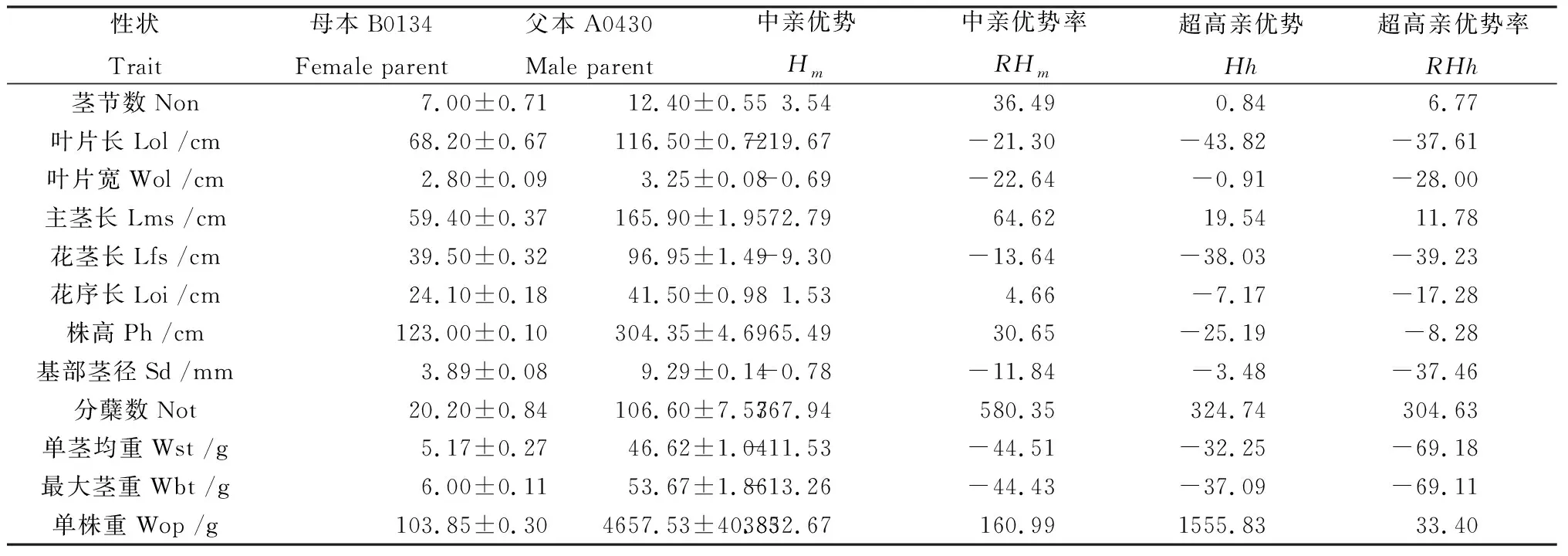
表2 荻与五节芒杂交种F1群体主要农艺性状的杂种优势Table 2 Heterosis of agronomical traits in F1 population between M. sacchariflorus and M. floridulus
2.3 12个主要农艺性状的相关性分析
应用DPS Statistics 14.7软件的Pearson方法对植株与产量相关的12个主要农艺性状进行相关性分析(表3),结果表明,株高和基部茎径是芒属植物的重要性状,这2个性状都与单茎均重和最大茎重达到极显著的相关,但都与单株产量没有相关性,株高与基部茎径显著相关;叶片大小与茎秆长短性状、分蘖数和产量存在显著或极显著的相关性,说明通过叶片大小的间接选择可以筛选高产单株;单株产量与叶片长宽、分蘖数、单茎均重、最大茎重有显著或极显著的相关性,尤其与分蘖数达到了极显著相关(r=0.826),与茎相关性状中只与主茎长达到了显著相关,说明单株分蘖数对单株产量具有特别重要的作用,这一结果对开展芒属植物高生物质产量的遗传改良有极重要的指导价值。
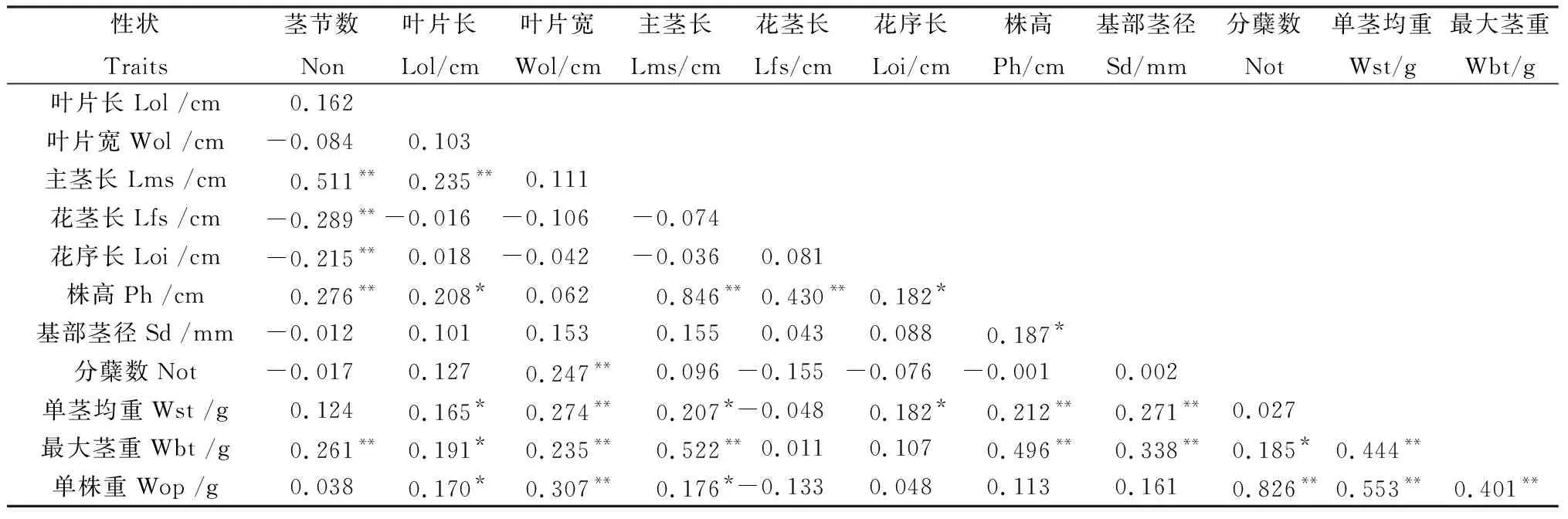
表3 荻与五节芒杂交F1群体主要农艺性状的表型相关分析Table 3 Correlation analyses among 12 agronomical traits in F1 population between M. sacchariflorus and M. floridulus
2.4 荻与五节芒杂交种F1群体主要农艺性状主效+多基因混合模型分析
2.4.1荻与五节芒杂交种F1群体主要农艺性状最适遗传模型的适合性分析 利用植物数量性状主基因+多基因混合遗传模型单个分离世代的分离分析方法,对荻B0134×五节芒A0430杂交种F1群体植株12个性状2年表型数据均值进行混合遗传模型分析,获得了无主基因、1对主基因(A)和2对主基因(B)共计11种模型与表型资料的配伍情况,得到极大似然值和AIC值(表4)。根据AIC最小准则,AIC值最小者为最优模型,与最小AIC值比较接近的遗传模型作为备选模型。
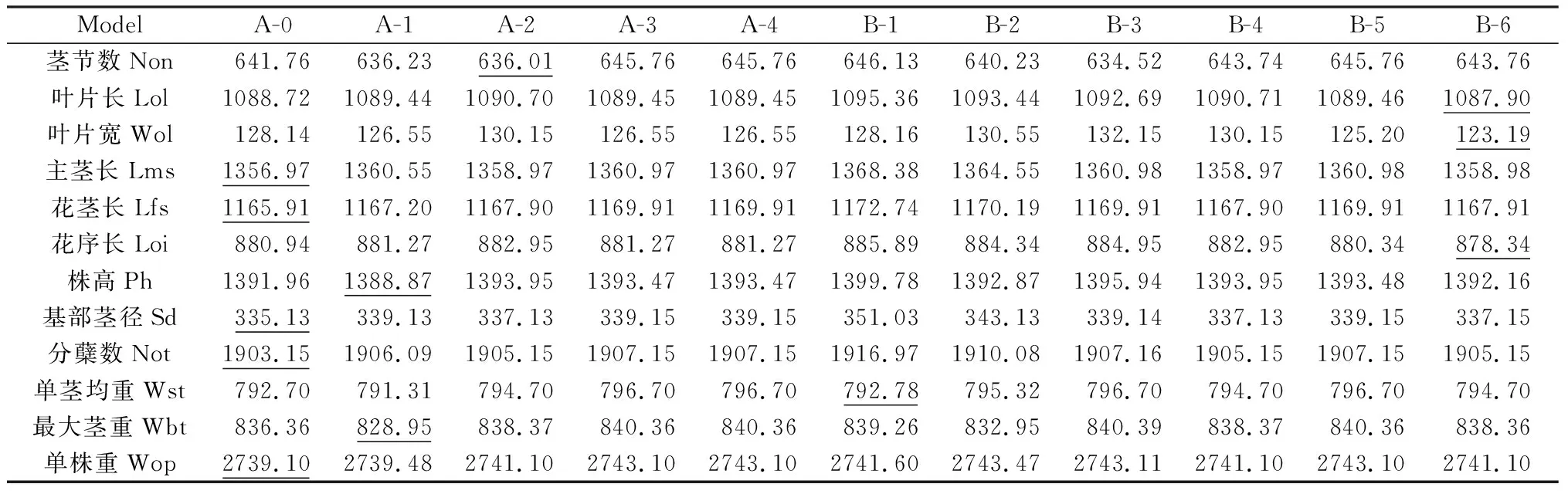
表4 荻与五节芒杂交F1群体不同遗传模型下的AIC值Table 4 Akaike's information criterion (AIC) under different genetic model for different traits of F1 population between M. sacchariflorus and M. floridulus
2.4.2荻与五节芒杂交种F1群体主要农艺性状遗传参数估计 表6是根据各主要农艺性状遗传模型参数的极大似然估计值,估计最适遗传模型的遗传参数。由表6可知,控制茎节数性状的遗传模型为1MG-A,1对主基因控制的加性效益值为2.61。控制叶片性状叶片长、叶片宽和花序长性状的都是2MG-EEAD遗传模型,3个性状的2对主基因加性效应均为正值(5.46,0.23,2.71),2对主基因加性效应和显性效应基本相等,且为上位性效应。控制株高和最大茎重的1对主基因表现为加性和显性效应,株高以加性效应为主(34.43),而最大茎重的加性效应(3.57)与显性效应几乎相等,但显性效应为负值(-3.56)。单茎均重的2对主基因的加性效应大小接近(2.35,2.23),2对基因的显性效应均为负值(-3.27,-2.07),2对主基因的加-加效应、加-显效应、显-加效应和显-显效应分别为2.09,-2.12,-1.00和3.25,说明主基因之间的上位性互作效应并不太大。茎节数、叶片宽、株高的基因型方差大于表型方差,主基因遗传率较高分别为76.24%,60.74%,75.49%,均为较高遗传力性状。其他性状都检测不到主基因,尤其是杂种优势非常显著的单株重和分蘖数,其原因有待进一步探讨。
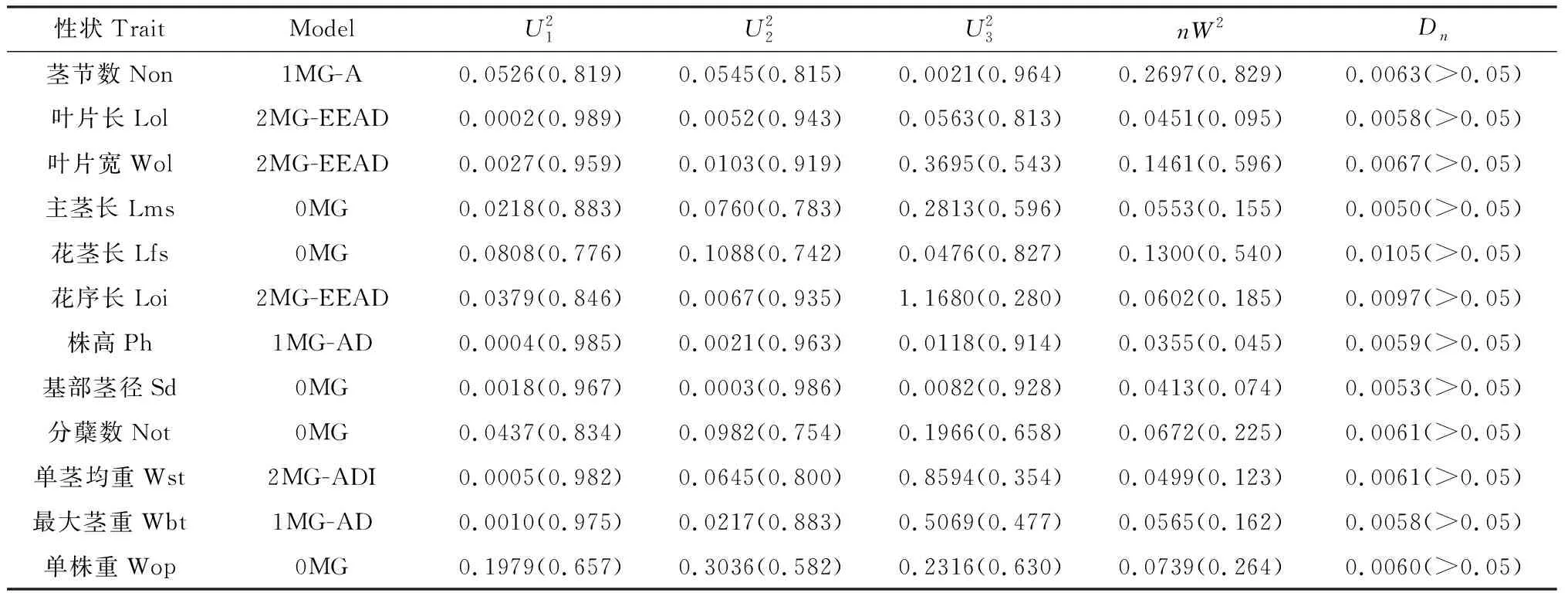
表5 荻与五节芒杂交F1群体农艺性状入选模型的适合性检验Table 5 Fitness tests of selected genetic model of 12 agronomical traits of F1 population between M. sacchariflorus and M. floridulus
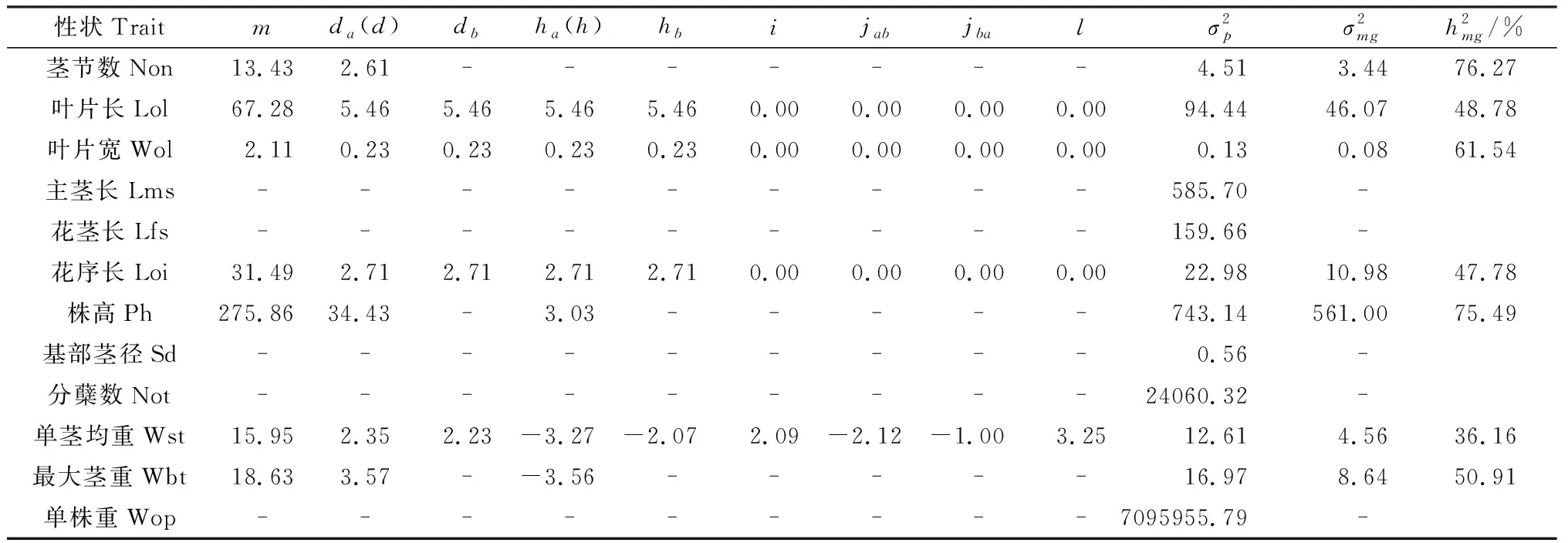
表6 农艺性状在入选模型下的遗传参数估计Table 6 Genetic parameter estimate of agronomical traits under the selected model
3 讨论
荻与五节芒的种间杂交优势显著,F1群体植株的多数性状倾向中亲优势,但分蘖数、单株重出现显著的超亲优势,群体内株间差异明显,遗传多样性丰富,可供选择的范围广泛,这与假俭草和菊花杂交后代的研究结果相似[19-20]。荻与五节芒杂种F1群体主要农艺性状的变异系数在9.81%~91.73%之间,株高和基部茎径的变异系数相对较小,其他性状的变异系数都在13%以上,尤以分蘖数、单茎均重、最大茎重等与产量关系密切的相关性状变异系数大,说明这些性状比较容易通过系统选育或杂交育种进行遗传改良选育新品种。更重要的是荻与五节芒杂交种F1群体在主要农艺性状上具有较大的杂种优势,尤其是主茎长、分蘖数、单株重的中亲优势率和高亲优势率均达到了极显著水平,与前人在杂合性比较高的植物中杂种优势的研究结果一致[26,28]。本研究发现杂交后代植株中出现杂种优势的比例极高,尤其是在茎节数、主茎长、分蘖数和单株重性状上超过60%的杂交后代植株有超亲优势,尤其是所有的杂交后代植株在分蘖数性状上全部超过了高亲,这在植物杂交育种和杂种优势利用上是少见的现象。荻具有很强的地下根状茎扩张能力[21-22],五节芒具有较强的萌芽能力和对贫瘠土壤的耐受力[23],性状差异大,性状互补,亲缘关系远,遗传差异大,各种基因效应都存在,显性效应和互补效应对杂种优势可能具有较大的贡献[29],具体原因还有待进一步的研究。
在主要农艺性状的相关性方面,分蘖数与最大茎重呈显著正相关性,与叶片宽和单株重呈极显著正相关性,其他性状没有相关性,这与鸭茅(Dactylisglomerata)品种(系)表型分析研究结果基本一致[30],可以独立的进行选择。基部茎径与株高呈显著正相关,与单茎均重和最大单茎均重呈极显著正相关,而与其他性状无相关性,尤其是与分蘖数和单株产量无相关性,也同样可以独立选择。在整个农艺性状相关性分析中,只有茎节数与花茎长、花序长呈极显著的负相关,其他有相关性的性状中,都是正相关,说明对某一性状的选择都对其他性状有促进作用,尤其是单株重与其他性状没有出现负相关关系,这对选择高生物质产量的优良单株具有较大的指导作用。
本研究利用盖钧镒等[10]提出的主基因+多基因混合遗传模型对荻和五节芒种间杂交种F1群体的12个主要农艺性状进行了遗传分析,只有茎节数、叶片长、叶片宽、株高、分蘖数和单茎均重6个性状能检测到主基因的存在,主茎长、花茎长、花序长、基部茎径、最大茎重、单株重共6个性状为无主基因模型。由于缺乏芒属植物性状遗传的数据,本试验的结果是否能合理的解释芒属植物性状遗传情况,以及荻和五节芒种间杂交群体是否存在远缘杂交中经常会发生的异常遗传的现象,也是一个需要继续深入探讨的问题,尤其需要建立多个遗传群体,包括种间和种内的杂交群体。对于在本研究中没有主基因存在的性状,因亲本的杂合性和模型的限制,是否存在多基因以及多基因的效应无法做出判断,这种情况在植物的遗传研究中也充分的得到体现[13,15,31-32],其遗传机制的分析需要通过同质群体提供误差方差以鉴别其有无遗传变异存在。对于有主基因存在且主基因遗传率较高的性状,可以在早期世代进行选择,育种时可以选用含有主基因的亲本,或者通过QTL基因定位的方法找到同这些基因紧密连锁的分子标记,采用分子标记辅助选择的方法来提高育种效果[29]。对于无主基因或在不同群体中主基因有无不确定的性状,建议采用轮回选择或回交育种的方法强化这些性状的表现。
在芒属植物的育种工作中,基部茎径性状不需要过多考虑[25],应重点关注单株分蘖数性状及主茎长和最大茎重等性状,尤其是分蘖数在芒属植物种间杂交种生物质产量中的作用。由于芒属植物高度自交不亲和[16],传统的杂交育种和杂种优势育种方法存在较大的局限。根据本研究荻和五节芒种间杂交种后代12个主要农艺学性状分离和杂种优势的表现,尤其是在生物质产量相关性状上的超高亲优势,容易从F1群体中筛选出生物质产量高且综合性状优良的单株,通过无性繁殖的方法选育无性系品种。还可以选择优良单株建立轮回选择群体,选育群体品种,不仅能选育出高生物质产量的优良品种,也解决了无性系品种种苗繁殖的困难,这应该是今后芒属植物遗传改良的主要方向。
