ABCG1在肿瘤坏死因子α诱导的氧化应激中的机制研究*
2017-08-28薛嘉虹朱参战胡艳超栾春红
薛嘉虹,朱参战,胡艳超,栾春红
(西安交通大学医学院第二附属医院 心内科,陕西 西安710004)
ABCG1在肿瘤坏死因子α诱导的氧化应激中的机制研究*
薛嘉虹,朱参战,胡艳超,栾春红
(西安交通大学医学院第二附属医院 心内科,陕西 西安710004)
目的 探讨三磷酸腺苷结合盒转运体G1(ABCG1)在肿瘤坏死因子α(TNF-α)诱导的氧化应激中的作用及可能的机制。方法 人脐静脉内皮细胞被特异性ABCG1 siRNA或ABCG1过表达质粒转染或使用LXR(肝X受体)激活剂T0901317预处理,随后给予肿瘤坏死因子(TNF-α)干预12 h。采用DCFHDAAM(2’7’-二氯荧光双乙酸盐)荧光探针检测细胞内活性氧簇(ROS)的水平,分光光度仪测量还原型烟酰胺腺嘌呤二核苷酸磷(NADPH)氧化酶活性,实时荧光定量聚合酶链反应法(qRT-PCR)和Western blot检测内皮细胞NADPH氧化酶亚型非吞噬细胞氧化酶4(NOX4)表达及超氧化物歧化酶(SOD)的表达。结果 ABCG1表达上调抑制TNF-α诱导的氧化应激,同时抑制促氧化应激的NADPH氧化酶的活性和NOX4的表达,促进抗氧化的SOD表达。相反,ABCG1表达下调进一步诱导ROS的产生,诱导NADPH氧化酶的活性和NOX4的表达,抑制SOD1表达。结论 ABCG1通过调节NADPH氧化酶/SOD抑制TNF-α诱导的氧化应激。
三磷酸腺苷结合盒转运体G1;氧化应激;还原型烟酰胺腺嘌呤二核苷酸磷氧化酶;超氧化物歧化酶
研究表明,氧化应激以及在氧化应激过程中产生的活性氧(reactive oxygen species,ROS)与多种心血管疾病的发生、发展有着密切关系[1-4]。内源性ROS产生除了主要来源于线粒体呼吸链外,还原型烟酰胺腺嘌呤二核苷酸磷(reduced nicotinamide adenine dinucleotide phosphate,NADPH)氧化酶是体内ROS产生的另一主要来源[2-4]。此外,机体也存在着清除过多产生ROS的抗氧化系统[3],当氧化和抗氧化作用失衡,即可导致大量ROS的产生。
ABCG1属于三磷酸腺苷结合盒转运体G1(adenosine triphosphate binding cassette transporter G1,ABCG1)家族成员之一(矛盾),具有调节细胞内游离胆固醇流出,防止泡沫细胞形成的作用[5-7]。此外,有研究发现,ABCG1也在内皮细胞高度表达,其促进内皮细胞内固醇流出的作用对保护内皮细胞正常功能具有十分重要的作用[8-11]。既往研究中,发现ABCG1表达上调可以降低TNF-α诱导的血管内皮细胞损伤,其作用可能与抑制肿瘤坏死因子-α(tumour necrosis factor-α,TNF-α)诱导的血管内皮细胞氧化应激相关[12]。本研究拟进一步探讨ABCG1抑制TNF-α诱导的氧化应激可能涉及的机制。
1 材料与方法
1.1 细胞培养及实验干预
人脐静脉内皮细胞(human umbilical vein endothelial cells,HUVECs)购自美国 ATCC 细胞库。内皮细胞培养在含10%胎牛血清的低糖培养基(dulbecco's modified eagle medium,DMEM)中,37℃、5%二氧化碳CO2培养箱中静置培养。培养的HUVECs给予10 ng/ml的TNF-α干预12 h,或者预先使用肝X受体(liver X receptors,LXRs)配体T0901317(5μg/ml,Sigma)预处理HUVECs 2 h,或HUVECs被转染特异性ABCG1 siRNA或ABCG1过表达质粒,24 h后,细胞再给予10 ng/ml TNF-α干预12 h。正常培养的HUVECs为对照组。
1.2 核糖核酸提取及逆转录和实时荧光定量聚合酶链反应
收集干预结束后的HUVECs,用Trizol(美国in vitrogen公司)一步法提取细胞总核糖核酸(ribonucleic acid,RNA)。以紫外分光光度计测定总RNA的纯度(A260/A280)>1.8。取2μg总RNA逆转录成互补脱氧核糖核酸(complementary deoxyribonucleic acid,cDNA)。采用 SYBR Green I嵌合荧光法,以 3-磷酸甘油脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPD)基因为内参照,实时荧光定量聚合酶链反应(quantitative real time polymerase chain reaction,qRT-PCR)检测HUVECs中NADPH氧化酶亚型NOX4及超氧化物歧化酶(superoxide dismutase,SOD) 信使核糖核酸 (messenger ribonucleic acid,mRNA)的表达。引物的序列如下:NOX4正向引物5'-CTGGTGAATGCCCTCAACTT-3',反向引物:5'-GGCCAGGAACAGTTGTGAAG-3';SOD1 正 向 引物:5'-TAGCGAGTTATGGCGACGAA-3',反向引物:5'-TGCTGTATTATCTCCAAACT-3';SOD2 正 向 引物:5'-GCCCTGGAACCTCACATCA-3',反向引物:5'-TGACCACCACCATTGAACTT-3';GAPDH 正向引物:5'-TCATCCCTGCCTCTACTG-3',反向引物:5'-T GCTTCACCACCTTCTTG-3'。反应条件为:95℃温育10 s,95℃ 5 s,54℃ 20 s,读板温度;72℃ 12 s共 40个循环。每个反应做3个复孔。扩增完毕后行熔解曲线分析,确定扩增产物的特异性。因目的基因与内参照基因的扩增效率一致,qRT-PCR统计分析采用2-ΔΔCt法。ΔΔCt=对照组△Ct(目的基因Ct-管家基因Ct)-各组△Ct(目的基因Ct-管家基因Ct)。
1.3 Western blot免疫蛋白检测
收集干预结束后的HUVECs,加入RIPA裂解液进行细胞裂解,于4℃离心10 min,弃除沉淀,用BCA法进行蛋白质定量。取50 g蛋白质加入1×上样缓冲液中,在100℃加热10 min使蛋白质变性。用10%十二烷基硫酸钠(sodium dodecyl sulphate,SDS)聚丙烯酰胺凝胶(polyacrylamide gel electrophoresis,PAGE)进行电泳分离,转硝酸纤维素膜(nitrocellulose filter membrane,NC),封闭液封闭 2 h,按说明书以1∶200加入兔抗人NOX4(美国Santa Cruz Biotechnology公司),兔抗人ABCG1(美国Santa Cruz Biotechnology 公司),4℃培育过夜,TBST(Tris-Hcl)缓冲溶液+吐温洗3次,1∶2 000加入辣根过氧化物酶标记山羊抗兔二抗1∶5 000(美国Santa CruzBiotechnology公司),室温培育1h,TBST洗3次,用增强化学发光液(enhanced chemilumincscence,ECL)(美国Amersham公司),冷电荷耦合元件(chargecoupled device,CCD) 照相机直接拍摄,Quality one软件分析。β-actin(美国Santa Cruz Biotechnology公司)为对照组。
1.4 细胞内活性氧检测
干预结束后的HUVECs和20 mmol DCFH-DA荧光探针(美国Sigma-Aldrich公司)37℃孵育20min,磷酸盐缓冲溶液(phosphate buffered solution,PBS)清洗3次,荧光显微镜测定细胞内荧光素的强度以表示细胞内ROS产生的多少。
1.5 NADPH氧化酶活性检测
根据细胞NADPH氧化酶活性光度法定量检测试剂盒说明书进行(上海一基实业有限公司)。样品准备好后,根据说明书依次加入缓冲液、反应液、底物液,即刻放入分光光度仪,在λ=340nm处测定NADPH氧化酶的活性,结果为样品总活性-样品非特异性活性。
1.6 ABCG1干扰和过表达
参考既往实验[13],选用化学合成的ABCG1干扰序列正向引物:5'-GAGUCUUUCUUCGGGAACATT-3',反向引物:5'-UGUUCCCGAAGAAAGACUCTT-3'(上海吉玛生物制药公司)。ABCG1 siRNA和随机siRNA序列使用TurboFect siRNA转染试剂(立陶宛Fermentas公司)转染HUVECs,转染24 h后细胞用于随后的实验干预。绿色荧光标记的ABCG1过表达质粒EX-Z0509-M61和对照载体EX-EGFP-M61由美国Gene CopoeiaTM公司设计合成并转染HUVECs,Western blot验证转染效率,结果见[13]。
1.7 统计学方法
数据分析采用SPSS 19.0统计软件,计量资料以均数±标准差(±s)表示,多组间比较采用单因素方差分析,两两比较用LSD-t检验,P<0.05为差异有统计学意义。
2 结果
2.1 ABCG1表达改变对TNF-α诱导的内皮细胞氧化应激的影响
TNF-α干预HUVECs后,内皮细胞内荧光强度较对照组增加,而使用T0901317预处理HUVECs组,内皮细胞内荧光强度则减弱,提示使用T0901317后内皮细胞内活性氧产生减少。当进一步使用ABCG1质粒转染HUVECs,使内皮细胞ABCG1表达增高后,再给予TNF-α干预,内皮细胞内荧光强度类似T0901317干预组,同样较单纯TNF-α干预的内皮细胞荧光强度减弱。相反,使用ABCG1 siRNA转染HUVECs后,结果发现细胞内荧光强度较单纯TNF-α干预的内皮细胞有所增强,提示活性氧进一步产生。见图1。

图1 ABCG1表达改变对TNF-α诱导的内皮细胞氧化应激的影响 (×100)
2.2 ABCG1表达改变对NADPH氧化酶活性的影响
在TNF-α干预组,HUVECs的ABCG1表达较对照组降低(t=31.00,P=0.001),NADPH 氧化酶活性则较对照组增加(t=30.10,P=0.001),使用 T0901317预处理或ABCG1质粒转染细胞后再给予TNF-α干预,结果发现,随着内皮细胞ABCG1表达增加,NADPH氧化酶活性被逆转,较TNF-α干预组降低(T0901317干预组 t=27.58,P=0.001,ABCG1质粒转染组t=29.76,P=0.001),几乎接近对照组的水平。在使用ABCG1 siRNA转染组,ABCG1表达进一步降低,内皮细胞NADPH氧化酶活性较对照组增加(FABCG1表达=12.858,PABCG1表达=0.000;FNADPH氧化酶活性=700.108,PNADPH氧化酶活性=0.000)。见图 2。

图2 ABCG1表达改变对NADPH mRNA氧化酶活性的影响
2.3 ABCG1表达改变对NOX4表达的作用NOX4作为NADPH氧化酶亚单位之一,与NADPH氧化酶的其他亚单位相比较,主要表达在内皮细胞。本实验结果提示,在TNF-α干预组,HUVECs的NOX4 mRNA表达和蛋白表达较对照组增加(t=48.00 和22.25,均P=0.000),而使用T0901317预处理或ABCG1质粒转染细胞后再给予TNF-α干预,HUVECs的NOX4 mRNA表达和蛋白表达均较TNF-α干预组降低(在mRNA表达方面,tT0901317组=26.50,PT0901317组=0.000,tABCG1质粒转染组=19.33,PABCG1质粒转染组=0.000;蛋白表达方面 tT0901317组=24.50,PT0901317组=0.000;tABCG1质粒转染组=21.60,PABCG1质粒转染组=0.000)。使用ABCG1 siRNA转染细胞后,内皮细胞NOX4 mRNA表达和蛋白表达较对照组增加(tNOX4mRNA表达=262.519,PNOX4mRNA表达=0.000;tNox4蛋白表达=304.004,PNOX4蛋白表达=0.000)。但与 TNF-α 干预组比较,差异无统计学意义(t=2.25和2.00,均P>0.05)。见图3。
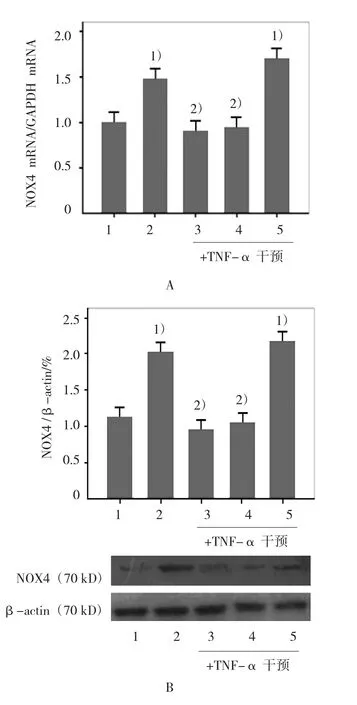
图3 ABCG1表达改变对NOX4 mRNA和蛋白表达的影响
2.4 ABCG1表达改变对SOD mRNA表达的影响
SOD是体内重要的抗氧化酶。当TNF-α干预HUVECs后,HUVECs的SOD1和SOD2 mRNA表达较对照组降低(t=3.62 和 3.10,均 P <0.05),而使用T0901317预处理或ABCG1质粒转染细胞后,随着ABCG1表达增加,内皮细胞的SOD1和SOD2 mRNA表达较TNF-α干预组增加(FSOD1mRNA=800.119,PSOD1mRNA=0.000;FSOD2mRNA=504.138,PSOD2mRNA=0.000)。当使用ABCG1 siRNA转染内皮细胞使ABCG1表达降低后,内皮细胞SOD1 mRNA表达较TNF-α干预组进一步降低(t=3.22,P <0.05),而 SOD2 mRNA表达较TNF-α干预组轻微降低,但差异无统计学意义(t=2.051,P=0.06)。见图 4。

图4 ABCG1表达改变对SOD1和SOD2 mRNA表达的影响
3 讨论
氧化应激为各种原因导致机体内氧化与抗氧化之间平衡失调,活性氧簇产生过多,使机体处于促氧化状态,引发各种各样心血管疾病。当前研究发现,炎症因子TNF-α能促进体外培养的内皮细胞的氧化应激产生,而作为脂质转运体的ABCG1可以通过影响NADPH氧化酶的促氧化作用和SOD的抗氧化作用从而调节TNF-α诱导的氧化应激的产生。
研究表明,ABCG1是促进细胞内胆固醇流出的重要膜转运体之一,能防止泡沫细胞形成,并有防止内皮细胞激活,改善内皮功能失调的作用[5-11]。ABCG1表达增加可以抑制内皮细胞炎症因子如TNF-α、IL-6及ICAM等的释放[9-10],从而有助于防治动脉粥样硬化的发生、发展。本研究进一步发现,ABCG1表达增加可抑制内皮细胞氧化应激的产生[11-12]。通过使用T0901317预处理内皮细胞,结果发现T0901317能降低TNF-α诱导的内皮细胞氧化应激。由于T0901317作为肝X受体配体,具有促进脂质转运体ABCG1表达的作用[5],因此提示T0901317干预后的抗氧化应激的作用可能与促进ABCG1表达相关,进一步通过ABCG1转染使ABCG1表达增加也发现内皮细胞氧化应激明显降低,而ABCG1干扰使ABCG1表达抑制则促进炎症因子诱导的内皮细胞氧化应激。正如TERASAKA[8]和TABET等[14]的研究所示,高密度脂蛋白降低氧化应激的作用依赖于ABCG1的表达,间接提示ABCG1具有抑制氧化应激的作用。而笔者既往研究发现,ABCG1能调节高糖诱导的氧化应激[11],进一步提示促进ABCG1表达可能具有抑制多种原因诱导的内皮细胞氧化应激产生的作用,但机制尚不明确。
虽然线粒体电子传递链及黄嘌呤氧化酶、一氧化氮合酶和NADPH氧化酶等均可产生ROS,但研究显示,NADPH氧化酶是人血管系统中产生的ROS主要来源之一[2-4]。其中NOX4可能作为血管内皮细胞中主要的NADPH氧化酶参与内皮细胞内ROS的生成。研究显示,TNF-α作为体内重要的炎症因子,可以通过激活NADPH氧化酶促进氧化应激的产生[15]。本研究同样发现,TNF-α能促进NADPH氧化酶的活性和表达增加,ABCG1表达改变不仅影响着NADPH氧化酶的活性,还影响着NOX4的基因和蛋白表达,从而提示ABCG1可能通过抑制血管内皮细胞中NADPH氧化酶的表达和活性发挥抗氧化应激的作用。
另一方面,机体是一个有机的整体,当氧化应激产生过多时,抗氧化系统可代偿性激活,以减少ROS的产生[3]。体内已知的抗氧化系统包括抗氧化酶类系统和非酶类抗氧化剂,其中SOD是体内重要的抗氧化酶,可将毒性高的ROS转换为毒性较低的过氧化氢(hydrogen peroxide,H2O2),随后由谷胱甘肽过氧化物酶和H2O2酶继续作用,使H2O2转变成完全无害的水(H2O)和氧气(Oxygen,O2)。SOD 有 3种亚型,其中有核细胞主要表达SOD1和SOD2。本研究发现,TNF-α抑制抗氧化的SOD的表达,而ABCG1表达增加促进TNF-α干预下内皮细胞SOD1和SOD2基因的表达,而ABCG1表达降低则抑制内皮细胞SOD1基因的表达,SOD2表达轻微降低,但差异无统计学意义。提示ABCG1可能通过调节血管内皮细胞中抗氧化的SOD的表达进一步发挥抗氧化应激的作用。
总之,本研究提示ABCG1具有拮抗TNF-α诱导的氧化应激的作用,其机制可能涉及调节NADPH氧化酶/SOD的促氧化应激和抗氧化应激间的平衡。期望今后的临床治疗将促进ABCG1表达作为新型的抗氧化措施,为心血管疾病提供新的防治方案。
[1]MADAMANCHI N R,RUNGE M S.Redox signaling in cardiovascular health and disease[J].Free Radical Biology&Medicine,2013,61(8):473-501.
[2]DAN S,WEISS D,LASSÈGUE B,et al.Superoxide production and expression of nox family proteins in human atherosclerosis[J].Circulation,2002,105(12):1429-1435.
[3]BO C,LU Y,CHEN Y,et al.The role of Nrf2 in oxidative stress-induced endothelial injuries[J].Journal of Endocrinology,2015,225(3):83-99.
[4]GARCÍA-REDONDO A B,AGUADO A,BRIONES A M,et al.NADPH oxidases and vascular remodeling in cardiovascular diseases[J].Pharmacological Research,2016,114(10):110-120.
[5]DEMINA E P,MIROSHNIKOVA V V,SCHWARZMAN A L.Role of the ABC transporters A1 and G1,key reverse cholesterol transport proteins,in atherosclerosis[J].Molecular Biology,2016,50(2):223-230.
[6]MAULDIN J P,NAGELIN M H,WOJCIK A J,et al.Reduced expression ofATP-binding cassette transporterG1 increases cholesterol accumulation in macrophages of patients with type 2 diabetes mellitus[J].Circulation,2008,117(21):2785-2792.
[7]XUE J H,YUAN Z,WU Y,et al.High glucose promotes intracellular lipid accumulation in vascular smooth muscle cells by impairing cholesterol influx and efflux balance[J].Cardiovascular Research,2010,86(1):141-150.
[8]TERASAKA N,YU S,YVAN-CHARVET L,et al.ABCG1 and HDL protect against endothelial dysfunction in mice fed a highcholesterol diet[J].Journal of Clinical Investigation,2008,118(11):3701-3713.
[9]WHETZEL A M,STUREK J M,NAGELIN M H,et al.ABCG1 deficiency in mice promotes endothelial activation and monocyteendothelial interactions[J].Arteriosclerosis Thrombosis&Vascular Biology,2010,30(4):809-817.
[10]XUE S,WANG J,ZHANG X,et al.Endothelial ATP-binding cassette G1 in mouse endothelium protects against hemodynamic-induced atherosclerosis[J].Biochemical& Biophysical Research Communications,2016,477(2):247-254.
[11]XUE J,WANG C,ZHU C,et al.ATP-binding cassette transporter G1 protects against endothelial dysfunction induced by high glucose[J].Diabetes Research&Clinical Practice,2013,101(1):72-80.
[12]薛嘉虹,宋安齐,李欢,等.三磷酸腺苷结合盒转运体G1表达上调降低肿瘤坏死因子-α诱导的血管内皮细胞损伤[J].中国现代医学杂志,2013,23(5):1-5.
[13]XUE J,WEI J,DONG X,et al.ABCG1 deficiency promotes endothelial apoptosis by endoplasmic reticulum stress-dependent pathway[J].The Journal of Physiological Sciences,2013,63(6):435-444.
[14]TABET F,LAMBERT G,CUESTA TORRES L F,et al.Lipidfree apolipoprotein A-I and discoidal reconstituted high-density lipoproteins differentially inhibit glucose-induced oxidative stress in human macrophages[J].Arteriosclerosis Thrombosis&Vascular Biology,2011,31(5):1192-1200.
[15]LIN C P,HUANG P H,LAI C F,et al.Simvastatin attenuates oxidative stress,NF-κB activation,and artery calcification in LDLR-/-mice fed with high fat diet via down regulation of tumor necrosis factor-α and TNF receptor 1[J].PloS One,2015,10(12):e0143686.
ABCG1 inhibits TNF-α-induced oxidative stress by regulating NADPH oxidase and superoxide dismutase*
Jia-hong Xue,Can-zhan Zhu,Yan-chao Hu,Chun-hong Luan
(Department of Cardiovascular Medicine,the Second Affiliated Hospital of College of Medicine,Xi'an Jiaotong University,Xi'an,Shaanxi 710004,China)
ObjectiveTo explore the role of ATP-binding cassette sub-family G member 1 (ABCG1)in oxidative stress production induced by tumor necrosis factor α (TNF-α)and its possible mechanisms.MethodsHuman umbilical vein endothelial cells (HUVECs)were transfected with specific ABCG1 siRNA or ABCG1 overexpression plasmid or pretreated with liver X receptor agonist T0901317,then were cultured with TNF-α for 12 hours.Intracellular reactive oxygen species (ROS)levels were measured using 6-carboxy-2,7-dichlorodihydrofluorescein diacetate,diacetoxymethyl ester(CDCFHDA-AM)fluorescence and nicotinamide adenine dinucleotide phosphate-oxidase (NADPH oxidase)activity was measured by spectrophotometer.Real time PCR and Western blot were employed to measure the expression of Nox4,one of NADPH oxidase subunit and the expression of superoxide dismutase (SOD).ResultsABCG1 upregulation inhibited TNF-α-induced oxidative stress.Furthermore,the activity of NADPH oxidase and the expression of Nox4 were also suppressed by ABCG1 overexpression,but the expression of antioxidant SOD was promoted.Conversely,downregulation of ABCG1 by ABCG1 siRNA both increased the ROS production and promoted the NADPH oxidase activity and Nox4 expression.However,the expression of SOD1 was inhibited.ConclusionsThe results suggest thatABCG1 attenuates TNF-α-induced oxidative stress by regulating NADPH oxidase and SOD.
adenosine triphosphate binding cassette transporter G;oxidative stress;reduced nicotinamide adenine dinucleotide phosphate oxidase;superoxide dismutase
R363
A
10.3969/j.issn.1005-8982.2017.11.003
1005-8982(2017)11-0014-06
2016-10-27
国家自然科学基金(No:81100210)
